bab ii tinjauan pustaka - sinta.unud.ac.id · meningen, terbukti dari pemeriksaan kultur css, pcr...
TRANSCRIPT

BAB II TINJAUAN PUSTAKA
Fisiologi Cairan Serebrospinal
Cairan serebrospinal (CSS) terdapat dalam ruang ventrikel otak dan ruang
subarachnoid. Cairan serebrospinal penting sebagai lapisan pelindung sistem saraf
pusat dan mempunyai beberapa fungsi penting, yaitu sebagai medium fisiologis
untuk otak, menyediakan dukungan mekanik untuk otak, dimana otak mengambang
dalam CSS. Beberapa data terbaru menyebutkan bahwa CSS mempunyai peranan
penting untuk menjaga keseimbangan cairan interstitial dalam parenkim otak dan
mengatur fungsi sistem saraf (Oreskovic dan Klarica, 2010; Sakka dkk., 2011).
Secara anatomi CSS pada otak terbagi atas 2 bagian yang saling
berhubungan, yaitu: internal unit yang terdiri dari sistem ventrikel (2 ventrikel
lateralis, ventrikel ketiga, dan ventrikel keempat) dan eksternal unit berupa central
spinal canal yang terdiri dari ruang subarachnoid dengan sisterna (Oreskovic dan
Klarica, 2010).
Rata rata volume CSS 150 ml dalam ventrikel dan 125 ml di dalam ruang
subarachnoid. CSS terutama disekresi oleh plexus choroidalis, walaupun demikian
cairan interstitiil otak, ependima dan kapiler juga berperan dalam sekresi CSS. Villi
arachnoidalis dalam otak dan spinal merupakan tempat utama penyerapan CSS ke
dalam aliran sistem vena. Cairan serebrospinal diperbaharui sekitar 4 kali dalam 24
jam. Ruang CSS merupakan suatu sistem tekanan yang dinamis. Tekanan CSS
ditentukan oleh tekanan intrakranial, yang secara fisiologis antara 3-4 mmHg
sebelum usia 1 tahun dan antara 10-15 mmHg pada orang dewasa. Fungsi CSS
10

11
adalah sebagai proteksi hidromekanik dari SSP, berperan dalam perkembangan
otak dan regulasi homeostasis cairan interstitiil dalam jaringan otak yang
mempengaruhi fungsi neuron (Sakka dkk., 2011).
Komposisi CSS tidak sesederhana ultrafiltrat plasma. Konsentrasi Na, Cl
dan Mg lebih rendah dibandingkan dalam plasma. Jumlah sel dalam CSS biasanya
tidak melebihi 5 sel/mm. Variasi dalam komposisi CSS yang teregulasi secara
tertutup dapat digunakan untuk tujuan diagnosis (Sakka dkk., 2011).
Sirkulasi CSS merupakan fenomena dinamis dan regulasi sirkulasi CSS
berespon terhadap homeostasis serebral. CSS bersirkulasi dari tempat sekresi ke
tempat absorpsi. Aliran berjalan satu arah yaitu rostrocaudal pada rongga ventrikel
dan ke berbagai arah pada rongga subarachnoid. Aliran CSS bersifat pulsatil,
berperan pada denyut sistolik pada arteri choroidalis. CSS diproduksi oleh pleksus
choroideus pada ventrikel lateralis kemudian berjalan melalui foramen
interventrikular ke ventrikel ketiga, kemudian menuju ke ventrikel keempat melalui
aqua duktus serebri dan akhirnya memasuki rongga subarachnoid melalui median
aperture (foramen Magendie) pada ventrikel keempat. Pada rongga subarachnoid
cranial, CSS bersirkulasi secara rostral ke villi-villi tempat absorpsi, atau secara
caudal ke rongga subarachnoid spinal. CSS sebagian diabsorpsi oleh villi arachnoid
spinal, bersirkulasi secara rostral ke rongga subarachnoid cranial (Oreskovic dan
Klarica, 2010; Sakka dkk., 2011).

12
Definisi Meningitis bakterialis
Meningitis merupakan suatu terminologi yang menggambarkan adanya
inflamasi pada membran meningen dan/atau cairan serebrospinal (CSS) yang
membungkus dan melindungi otak dan korda spinalis. Meningitis dapat disebabkan
oleh berbagai kondisi baik infeksi maupun non infeksi. Meningitis akibat infeksi
dapat disebabkan oleh virus, bakteri, jamur dan parasit (Van de Beek, 2012).
Meningitis bakterialis khas ditandai oleh adanya sindrom infeksi dan pada
pemeriksaan CSS (cairan serebrospinal) dibuktikan adanya bakteri dan/atau terjadi
gambaran analisis yang abnormal secara bermakna. Adanya infeksi bakteri pada
meningen, terbukti dari pemeriksaan kultur CSS, PCR dari CSS, pengecatan gram
atau tes antigen. Secara klinis, suspek meningitis ditandai adanya gejala klinis
dan/atau secara klinis terdapat marker inflamasi pada CSS yaitu hitung leukosit,
kadar protein dan glukosa dalam CSS (SWAB, 2012).
Epidemiologi dan Etiologi Meningitis bakterialis
Insiden dan angka kematian bervariasi di berbagai negara. Angka kejadian
meningitis bakterialis di Amerika Serikat selama tahun 1998-2007 sebanyak 17,4
juta orang dan pada tahun 2006-2007 mencapai 1,38 per 100.000 orang/tahun
dengan angka kematian 14,3% (Thigpen dkk., 2011). Centers for Disease Control
and Prevention (CDC) pada tahun 2012 melaporkan angka kejadian meningitis di
negara Amerika Serikat sekitar 4100 kasus/tahun dan angka kematian akibat
meningitis mencapai 500 orang/tahun. Data WHO menyatakan kasus meningitis
diperkirakan mencapai kurang lebih 1,2 juta jiwa setiap tahunnya dan angka

13
kematian mencapai kurang lebih 135.000 jiwa. Meningitis menjadi salah satu dari
10 penyakit infeksi yang menyebabkan kematian di dunia serta 30-50% akan
mengalami sequele neurologis. Angka kematian mencapai 25% di negara maju dan
lebih tinggi di negara berkembang. Meningitis bakterialis terutama menyerang anak
usia dibawah 2 tahun, dengan puncak angka kejadian pada usia 6-18 bulan. Secara
keseluruhan tingkat kematian pasien meningitis bakterialis antara 2-30%
tergantung dari bakteri penyebab meningitis (Maimaiti dkk., 2012).
Angka insiden meningitis di Negara Amerika Serikat sekitar 4100
kasus/tahun dan meninggal akibat meningitis 500 orang/tahun (CDC, 2012). Di
Indonesia pada tahun 2010 jumlah kasus meningitis terjadi pada laki-laki kurang
lebih 12.010 jiwa, pada perempuan kurang lebih 7.371 jiwa, dan dilaporkan yang
meninggal dunia kurang lebih 1.025 jiwa (Menkes RI, 2012).
Meningitis disebabkan oleh virus, bakteri, jamur, mycobacteria maupun
parasit, organisme tersebut merupakan patogen fatal yang dapat menyebabkan
kematian. Tiga patogen utama yang menyebabkan meningitis bakterialis antara lain
Streptococcus pneumoniae (IPD= invasive pneumococcal diseases), Haemophilus
influenza type B (Hib), dan Neisseria meningitides (Houri dkk., 2017).
Meningitis bakterialis merupakan penyakit infeksi yang sangat merusak
dengan angka mortalitas di seluruh dunia mencapai 20-30% walaupun telah
mendapat terapi antibiotika. Mortalitas pada fase akut mencapai 10-20% dengan
sekuele 30%. Sebesar 50% dari pasien yang bertahan hidup mengalami gejala sisa
berupa gangguan pendengaran, kejang dan mengalami gangguan belajar dan
prilaku (Brouwer dkk., 2012; Jusot dkk., 2013).

14
Patogenesis Meningitis bakterialis
Secara umum meningitis bakterialis dapat terjadi ketika bakteri masuk ke
dalam sirkulasi sistemik dan selanjutnya invasif ke dalam SSP atau secara langsung
menyebar selama infeksi di sekitar SSP seperti infeksi telinga tengah atau
mastoiditis. Multiplikasi bakteri di dalam SSP merupakan trigger respon imun lokal
yang ditandai oleh masuknya leukosit. Pada kondisi normal atau sehat SSP terbebas
dari leukosit, namun pada keadaan patologi leukosit masuk ke dalam otak sebagai
respon terhadap berbagai stimulus (Chavez-Bueneo, 2005).
Meningitis disebabkan oleh bakteri pathogen yang memiliki virulensi poten,
selain itu faktor host yang rentan dan lingkungan yang mendukung memiliki
peranan besar dalam patogenesis infeksi. Infeksi dapat mencapai selaput otak
melalui beberapa cara: (Liechti dkk., 2015; Barichello dkk., 2013).
a. Aliran darah (hematogen) karena adanya infeksi di tempat lain seperti
faringitis, tonsilitis, endokarditis, pneumonia, infeksi gigi. Meningitis
bakterialis sebagian besar terjadi akibat penyebaran hematogen, dimana
bakteri melekat pada sel epitel mukosa sebagai port the entry kemudian
memperbanyak diri dalam aliran darah serta menimbulkan bakterimia.
Bakterimia dapat berlanjut masuk ke dalam cairan serebrospinal (melewati
sawar darah otak) dan memperbanyak diri dalam cairan serebrospinal
sehingga menimbulkan peradangan pada selaput otak (meningen) dan otak.
b. Perluasan langsung dari infeksi yang disebabkan oleh infeksi dari sinus
paranasalis, mastoid, abses otak dan sinus kavernosus.

15
c. Implantasi langsung dapat terjadi pada trauma kepala terbuka, tindakan
bedah otak atau pungsi lumbal.
d. Meningitis pada neonatus dapat terjadi oleh karena aspirasi cairan amnion
yang terjadi pada saat bayi melalui jalan lahir atau oleh kuman yang normal
ada pada jalan lahir.
Meningitis bakterialis dimulai dari masuknya bakteri ke dalam susunan
saraf pusat melalui makrovaskular otak atau pleksus koroid. Sawar darah otak
normal terdiri atas sel endotel dari kapiler darah otak. Astrosit akan membentuk
tight junction yang padat yang menghalangi lewatnya zat terlarut dalam darah
(elektrolit dan protein) atau sel sehingga lingkungan ekstrasel otak akan terpisah
dari darah dan mencegah terpajannya sel saraf terhadap perubahan elektrolit,
transmiter, hormon, faktor pertumbuhan dan reaksi imun (Srinivasan dkk., 2016).
Pada bayi dan anak dengan meningitis, tight junction terbuka sehingga
bakteri masuk dalam cairan serebrospinal, terjadi reaksi radang dan menyebabkan
permeabilitas sawar darah otak semakin meningkat. Bakteri yang masuk akan
bereplikasi, tersebar secara pasif mengikuti aliran cairan serebrospinal melalui
sistem ventrikel ke seluruh ruang subaraknoid (Srinivasan dkk., 2016).
Bakteri yang berkembang biak atau lisis melepaskan dinding sel/komponen
membran sel menyebabkan kerusakan jaringan otak serta menimbulkan
peradangan di selaput meningen. Komponen membran sel dari bakteri merangsang
sel endotel dan makrofag di susunan saraf pusat (sel astrosit dan mikroglia)
memproduksi mediator inflamasi seperti interleukin-1 (IL-1) dan tumor necrosis
factor (TNF). Mediator inflamasi berperan dalam proses awal dari beberapa

16
mekanisme yang menyebabkan peningkatan tekanan intrakranial, yang
selanjutnya mengakibatkan menurunnya aliran darah otak. Toksin dari bakteri
awalnya menimbulkan hiperemi pembuluh darah selaput otak disertai migrasi
neutrofil ke ruang subaraknoid, selanjutnya merangsang timbulnya kongesti dan
meningkatkan permeabilitas pembuluh darah, mempermudah adhesi sel fagosit
dan sel polimorfonuklear untuk menembus pembuluh darah melalui tight junction
dan memfagosit bakteri. Peningkatan tekanan intrakranial menyebabkan
penurunan aliran darah otak. Penurunan autoregulasi (penurunan tekanan darah
<60 mmHg sistole) dan kejang akan menurunkan tekanan perfusi serebral
menyebabkan iskemi dan kerusakan pada sel saraf sehingga menimbulkan gejala
sisa. Gangguan aliran darah otak dan peningkatan tekanan intrakranial akan
menyebabkan peningkatan kadar asam laktat dan penurunan pH cairan
serebrospinal yang disebabkan metabolisme anaerobik. Keadaan ini menyebabkan
penggunaan glukosa meningkat dan berakibat timbulnya glukosa CSS rendah
(Prasad dkk., 2014; Aggrawal dkk., (2017).
Bakteri mencapai SSP bisa secara hematogen atau melalui penyebaran
secara langsung dari regio yang berdekatan. Pada bayi dan anak berbagai organisme
yang menyebabkan meningitis berkolonisasi pada saluran pernafasan bagian atas,
kemudian terjadi bakterimia dan selanjutnya patogen berpenetrasi ke sawar darah
otak untuk masuk ke dalam ruang sub-arachnoid. Inokulasi langsung organisme ke
dalam SSP dapat terjadi pada kondisi trauma kepala, adanya defek seperti
meningomyelocele atau adanya perluasan dari fokal infeksi supuratif di
parameningeal. Protein pada permukaan bakteri yang telah diketahui dapat

17
memfasilitasi invasinya ke dalam sawar darah otak adalah protein E. Coli IbeA,
IbeB dan ompA; protein S.pneumoniae CbpA; dan protein N. Meningitidis Opc,
Opa, dan PilC, yang merupakan villi protein (Chavez-Bueno, 2005).
Inflamasi yang intensif terjadi akibat produk produk bakteri seperti
lipopolisakarida (LPS) dari bakteri gram negatif atau peptidoglikan dari bakteri
gram positif yang menetap setelah bakteri dihancurkan oleh respon host dan terapi
antibiotika. Substansi ini menginduksi produksi berbagai mediator inflamasi oleh
astrositosit, sel glia, sel ependymal, dan sel endotel. Mediator inflamasi yang
terlibat diantaranya: tumor necrosis factor α (TNF-α); interleukin (IL)-1, IL-6, IL-
8 dan IL-10; makrofag menginduksi protein 1 dan 2 dan mediator lain seperti nitric
oxide, matrix metalloproteinase-2 dan prostaglandin (Chavez-Bueno, 2005).
Meningitis bakterialis khas ditandai oleh pleositosis dalam CSS dengan
predominan polimorfonuklear (PMN). Rekruitmen leukosit merupakan kunci
proteksi respon imun melawan invasi mikroorganisme, tetapi bukti bukti penelitian
men unjukkan bahwa akumulasi leukosit juga berkontribusi penting dalam
terjadinya kerusakan jaringan otak pada infeksi meningitis bakterialis (Zwijnenburg
dkk., 2006).
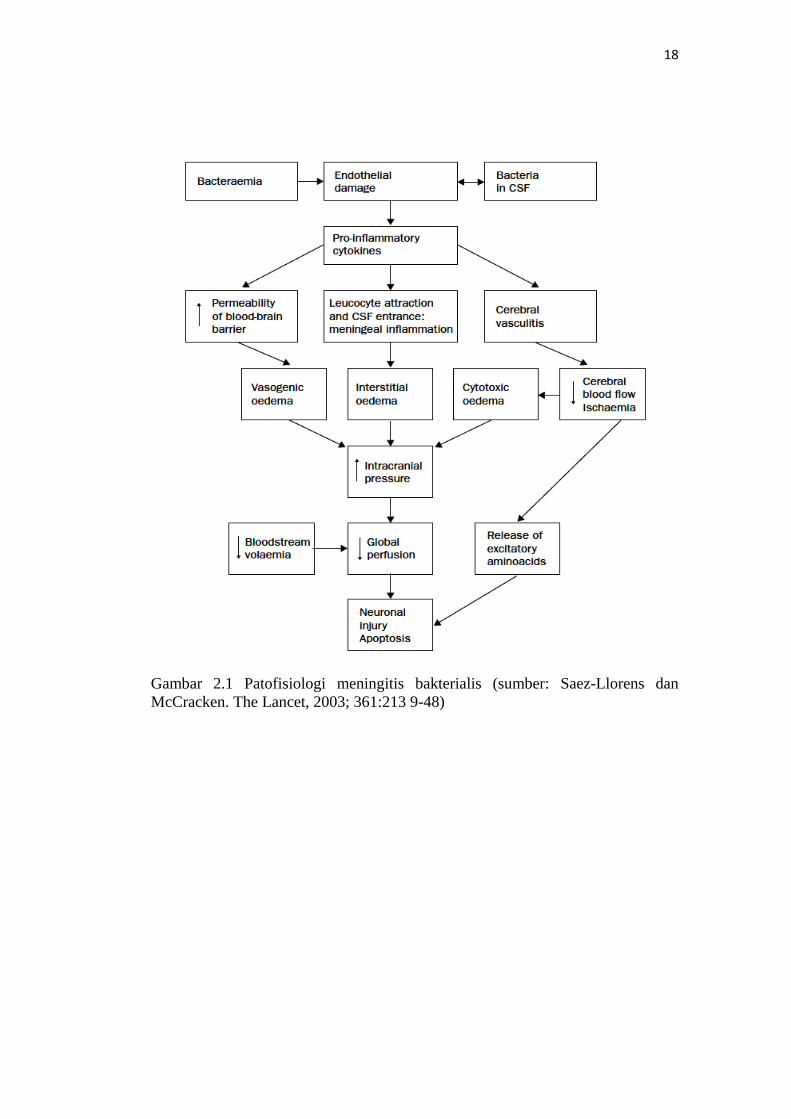
18
Gambar 2.1 Patofisiologi meningitis bakterialis (sumber: Saez-Llorens dan McCracken. The Lancet, 2003; 361:213 9-48)

19
Manifestasi Klinis Meningitis
Tanda dan gejala meningitis dapat bervariasi. Trias klasik meningitis adalah
demam, kaku kuduk dan penurunan kesadaran, namun tidak selalu tampak. Sebuah
penelitian prospektif menunjukkan trias meningitis yang klasik hanya ditemukan
44% dari 696 kasus dengan proven community-acquired bacterial meningitis
dewasa. Pada 95% pasien meningitis paling tidak ditandai 2 gejala dari 4 gejala:
sakit kepala, demam, kaku kuduk dan penurunan kesadaran. Beberapa gejala yang
sering pada meningitis adalah: sakit kepala (87%), kaku kuduk (83%), demam
(77%), dan penurunan kesadaran (69%). Trias klasik juga lebih sering muncul pada
pasien dengan meningitis pneumococcal (58%) dibandingkan dari meningitis
meningococcal (27%). Adanya rash terjadi pada 26% pasien dan lebih sering
dihubungkan dengan infeksi meningococcal. Rash juga dapat terjadi pada pasien
dengan hasil kultur positif untuk S pneumoniae, Staphylococcus aureus, H
influenzae, Listeria monocytogenes, dan group B streptococcus. Tanda Kernig and
Brudzinski, bersama sama dengan kaku kuduk merupakan tanda klasik “meningeal
signs” namun dapat tidak terjadi pada sebagian besar meningitis bakterialis
(Brouwer, dkk., 2012).
Faktor faktor yang dapat menjadi predisposisi meningitis bakterialis adalah
infeksi paru, telinga dan sinusitis, terjadi pada sekitar 40% pasien. Endokarditis
dapat juga merupakan faktor risiko tetapi sering terjadi bersama-sama. Pasien
dengan meningitis bakterialis juga sering menunjukkan gejala syok septik pada
sekitar 10-25% kasus (Brouwer dkk., 2012).

20
Tidak semua pasien meningitis bakterialis akut menunjukkan gejala
demam, kaku kuduk dan penurunan kesadaran. Demam diatas 38oC terjadi pada
sekitar 77% kasus. Gejala klinis dapat berupa infeksi akut dalam beberapa jam
sampai selama 1-2 hari dan bahkan beberapa hari. Sekitar 75% didahului oleh
infeksi saluran pernafasan akut, sekitar 20-30% disertai kejang. Kaku kuduk dapat
dijumpai pada anak yang lebih besar (>12-18 bulan) sekitar 80% kasus. Defisit
neurologis fokal dapat terjadi pada 33% kasus. Dengan demikian pada pasien yang
menunjukkan gejala suspek meningitis memerlukan pemeriksaan penunjang
diagnosis (Tunkel, dkk., 2004; NSW, 2014; Le Saux, 2014).
Pemeriksaan Analisis Cairan Serebrospinal pada Meningitis bakterialis
Punksi Lumbal harus dilakukan pada pasien dengan suspek meningitis.
Analisis CSS dapat menunjukkan warna keruh tergantung pada jumlah leukosit,
eritrosit, bakteri dan protein. Pada meningitis bakterialis yang belum mendapat
terapi antibiotika sel leukosit meningkat antara 1000-5000 sel/mm3, meskipun
dapat berkisar antara 100 sampai lebih dari 10.000 sel/mm3. Lebih dari 90% kasus
meningitis bakterialis akut menunjukkan hasil hitung leukosit dalam CSS >100
sel/µL. Walaupun demikian, beberapa laporan kasus menunjukkan adanya
meningitis bakterialis dengan hitung leukosit dalam CSS masih normal. Sekitar 80-
90% pasien menunjukkan PMN predominan dan hanya 10% pasien dengan limfosit
predominan. Beberapa penelitian menunjukkan 19% meningitis bakterialis yang
terdiagnosis melalui kultur menunjukkan hitung leukosit dibawah 100 sel/mm3
Tunkel dkk., 2004; Brouwer dkk., 2012; Agueda dkk., 2013).

21
Pada pasien meningitis bakterialis kadar glukosa CSS menurun
(hypoglycorrhachia) sedangkan kadar protein CSS meningkat. Kadar glukosa CSS
<40 mg/dL terjadi paling banyak pada 50-60% pasien. Rasio kadar glukosa
CSS/serum < 0,4 memiliki sensitivitas 80% dan spesifisitas 98% untuk diagnosis
meningitis bakterialis pada anak diatas 2 bulan. Pada neonatus rasio glukosa
CSS/serum <0,6 dapat digunakan sebagai acuan abnormal.
Kadar protein dalam CSS meningkat pada 90% pasien meningitis bakterialis
akut. Peningkatan kadar protein >50 mg/ dL dan penurunan kadar glukosa CSS
<40% dibandingkan kadar glukosa serum dianggap abnormal (Brouwer dkk., 2012;
Agueda dkk., 2013).
Pemeriksaan Mikrobiologi
Ada beberapa metode mikrobiologi untuk membuktikan adanya infeksi
bakteri pada meningitis, yaitu pemeriksaan kultur CSS, kultur darah, pewarnaan
gram CSS, aglutinasi lateks, dan PCR. Di Negara Negara berkembang, untuk
mendeteksi dan mengetahui karakteristik bakteri penyebab meningitis adalah
dengan melakukan pemeriksaan kultur, pengecatan gram dan latex aglutinasi.
Pemeriksaan kultur darah sampai saat ini merupakan metode baku emas
dalam mengidentifikasi adanya patogen bakteri penyebab infeksi. Pemeriksaan
kultur memerlukan waktu beberapa hari bahkan dalam hitungan minggu untuk
mendapatkan hasilnya, dimana dapat terjadi false-positive dan false negative
sebesar 2-3%. Faktor yang dapat mempengaruhi hasil antara lain jumlah sampel

22
darah yang kurang, sudah mendapat antibiotik sebelumnya, atau patogen penyebab
merupakan organisme yang khusus atau spesifik (Matsuda dkk., 2011).
Kultur merupakan standar baku emas untuk konfirmasi klinis, namun angka
positif-nya relatif rendah akibat penyimpanan dan transport specimen yang kurang
adekuat, metode kultur, atau pemberian terapi antibiotika sebelum pengambilan
specimen. Pengecatan gram merupakan pemeriksaan yang penting, tidak mahal dan
harus dilakukan jika memungkinkan, hanya saja pengecatan gram hanya
memberikan clue genus dan spesies dari bakteri penyebab. Latex aglutinasi bersifat
subyektif dan sulit untuk interpretasi, terutama jika bacterial load rendah (CDC,
2014).
2.7.1 Kultur dan tes sensitivitas bakteri
Cairan serebrospinal sangat diperlukan untuk mengidentifikasi organisme
penyebab dan pemeriksaan sensitivitas terhadap antibiotik secara in vitro. Kultur
harus tetap digunakan sebagai standar baku emas untuk diagnosis meningitis
bakterialis, karena merupakan sumber data untuk suseptibilitas antibiotika,
penentuan subtipe secara lengkap, ekspresi antigen termasuk untuk mengetahui
patofisiologi isolat serta penelitian vaksin di masa yang akan datang (Brouwer dkk.,
2010; 2012; CDC, 2014a).
Kultur CSS positif pada 80-90% pasien meningitis bakterialis yang didapat
dari komunitas, jika belum mendapat terapi antibiotika tetapi hasil kultur baru
diketahui setelah 5-7 hari (Brouwer dkk., 2010; 2012).
Jika pasien diduga meningitis, pemeriksaan kultur darah harus dilakukan
secara rutin sebelum pemberian antibiotika. Kultur darah harus selalu dilakukan

23
pada saat pasien masuk rumah sakit karena sangat membantu pada pasien yang
telah mendapat antibiotika sebelum dikerjakan punksi lumbal. Kultur darah dapat
membantu isolasi organisme jika pengambilan CSS dilakukan beberapa jam
sebelum pemberian antibiotika dan pada kasus dimana tidak didapatkan sampel
CSS untuk pemeriksaan analisis maupun kultur CSS. Kultur darah mengidentifikasi
organisme penyebab sekitar 50-80%. Sensitivitas kultur darah menurun 20% pada
pasien yang telah mendapat antibiotika sebelumnya (Brouwer dkk., 2012).
2.7.2 Pengecatan Gram
Pemeriksaan pengecatan Gram CSS merupakan pemeriksaan penting yang
dapat dilakukan dengan cepat, tidak mahal dan cukup valid untuk menilai adanya
bakteri dalam CSS. Pengecatan gram juga dapat memberikan gambaran genus dan
spesies bakteri penyebab. Pengecatan gram memiliki sensitivitas 90% dan
spesifisitas 97% dalam diagnosis meningitis bakterialis. Kelemahan pengecatan
gram adalah memerlukan jumlah sampel yang lebih banyak untuk memperoleh
hasil yang baik dan manfaat pengecatan gram dapat menurun 20% pada pasien yang
sebelumnya telah mendapatkan terapi antibiotika (Brouwer dkk., 2012)
2.7.3 Latex aglutinasi
Latex aglutinasi merupakan tes diagnosis yang cepat dengan sensitivitas 50-
100% tergantung pada bakteri patogen. Tes ini sangat sensitif terhadap infeksi oleh
H. influenza dan paling sensitif terhadap infeksi oleh N. meningitides. Tes ini tidak
dapat mengubah keputusan terapi jika positif sehingga sangat membantu untuk
diagnosis pada pasien yang telah mendapat terapi antibiotika dimana pengecatan
gram dan kultur menunjukkan hasil yang negatif (Brouwer dkk., 2012).

24
2.7.4 Identifikasi bakteri dengan PCR (Polymerase Chain Reaction)
Pada penegakan diagnosis meningitis bakterialis, metode pengecatan gram
adalah metode yang cepat dan spesifik dalam mendeteksi adanya bakteri di cairan
serebrospinal namun memiliki sensitivitas yang rendah. Pemeriksaan kultur dinilai
lebih sensitif, dimana dapat mengidentifikasi >80% patogen pada pasien dengan
meningitis bakterialis sebelum mendapat terapi, akan tetapi hasil pemeriksaan ini
bergantung pada jumlah bakteri yang dapat dibiakkan atau tumbuh dalam media
kultur. Pemeriksaan PCR yaitu suatu metode penjamakan asam nukleat sehingga
dapat mengidentifikasi DNA patogen walaupun dalam jumlah sedikit.
Tes PCR didasarkan pada penguatan konservasi gen rRNA mampu
mendeteksi dan membedakan barbagai bakteri dan jamur. Bakteri patogen dideteksi
dengan PCR 16s rRNA pada cairan serebrospinal pasien dengan meningitis
bakterialis dengan nilai sensitivitas dan spesifisitas yang sangat baik, selain itu 30%
kasus kecurigaan meningitis bakterialis memiliki hasil tidak ada pertumbuhan
kuman pada pemeriksaan kultur cairan serebrospinal namun memiliki hasil PCR
yang positif, hal ini menunjukan sensitifitas PCR lebih tinggi dibandingkan
pemeriksaan kultur (Meyer, 2014).
Teknologi PCR telah digunakan untuk mengamplifikasi jumlah DNA dari
pasien yang menderita meningitis yang disebabkan oleh patogen meningeal yang
umum, seperti N. meningitidis, S. pneumoniae , H. influenzae tipe B, S. agalactiae,
dan L. monocytogenes.
Pemeriksaan PCR digunakan secara luas dalam diagnosis dan surveilens
bakteri patogen karena memiliki sensitivitas dan spesifisitas yang tinggi. PCR dapat

25
menjadi pelengkap metode klasik yang berdasarkan fenotip seperti kultur,
pengecatan gram dan latex aglutinasi. Sensitivitas dan spesifisitas dari PCR
dilaporkan berkisar 91% pada penelitian yang menggunakan sampel CSS (Matsuda,
dkk., 2011).
Pada PCR, target DNA secara eksponensial diamplifikasi melalui 3 tahap
yaitu: (1) Denaturasi double stranded DNA (DNA utas ganda) menjadi single
stranded DNA (DNA utas tunggal); (2) annealing dari primers terhadap
complementary single stranded target sekuensing ; (3) Ekstensi primers dalam
urutan 5’ ke 3’ oleh heat-stable DNA polymerase sehingga menghasilkan molekul
DNA utas ganda. Metode PCR tidak memerlukan sel yang intak atau sel hidup
sehingga PCR merupakan metode yang bernilai untuk deteksi bakteri penyebab dari
spesimen dimana bakterinya sudah mati atau lisis akibat penyimpanan yang kurang
sesuai atau akibat pemberian antibiotika sebelumnya (CDC, 2014b).
Untuk pemeriksaan PCR metode konvensional membutuhkan waktu yang
sangat banyak, kurang sensitif dan kurang spesifik dibandingkan real time-PCR
(RT-PCR) (Wu dkk., 2013).
Teknik PCR memiliki potensi yang menjanjikan untuk mengidentifikasi
bakteri penyebab meningitis. Pengembangan lebih lanjut dari teknik yang ada dapat
menghasilkan kegunaan PCR lebih banyak lagi untuk mengidentifikasi pasien
penderita meningitis yang menunjukkan hasil negatif pada pemeriksaan dengan
pewarnaan gram maupun kultur (CDC, 2014b).

26
2.7.5 PCR dengan universal primers
Sebuah penelitian klinis menggunakan PCR untuk diagnosis meningitis
telah dilakukan pada tahun 2003, menggunakan broad-range bacterial primers.
Primers dikonservasi dari bakteri region gen 16S rRNA. Teknik ini dapat digunakan
untuk mengidentifikasi organism S. pneumoniae, Nisseria meningitides, Listeria
monocytogenes dan Mycobacterium tuberculosis dengan menggunakan species-
specific primers. Broad range bacterial PCR ini menunjukkan sensitivitas 100%,
spesifitas 98,2%, nilai prediktif positif (NPP) 98,2% dan nilai prediktif negatif
(NPN) 100%. Oleh karena itu, broad-based PCR dapat berguna untuk
mengidentifikasi bakteri, sehingga dapat menentukan diagnosis dan terapi yang
tepat (Saravolatz dkk., 2003).
Penerapan strategi universal primers telah menjadi teknik konvensional
yang unggul dalam mendeteksi bakteri pada cairan tubuh yang steril. Namun risiko
relatif kontaminasi menjadi masalah utama dalam penggunaan PCR universal
tunggal sebagai uji spesifik dalam menegakkan diagnosis. Spesifisitas dari broad
range universal PCR telah dikembangkan dengan analisis lebih lanjut terhadap
produk PCR dengan berbagai metode seperti restriction fragment length
polymorphism (RFLP), probing atau sequenzing (Chakrabarti dkk., 2009).
Metode pemeriksaan dengan menggunakan primers 16S rDNA berbasis
PCR merupakan temuan yang menjanjikan sebagai sebuah uji skrining untuk
diagnosis meningitis bakterialis akut, khususnya di rumah sakit dimana hampir
setiap pasien telah mendapatkan antibiotik. Metode 16S rDNA berbasis PCR

27
mengunakan primers bakteri universal yang berguna mendeteksi bakteri patogen
dalam sampel cairan serebrospinal.
Gen RNA ribosom yang penting dari bakteri yaitu 23 rRNA region
menunjukkan variasi yang lebar antar berbagai spesies bakteri jika dibandingkan
dengan 16S rRNA region. Sensitivitas, spesifisitas, nilai prediktif positif (NPP) dan
nilai prediktif negatif (NPN) secara keseluruhan dari PCR 16S rDNA masing-
masing adalah 79.24%, 97.6%, 89.36% dan 94.88%, dimana kultur digunakan
sebagai baku emas (Chakrabarti dkk., 2009).
2.7.6 Identifikasi bakteri dengan PCR gene 16S rRNA
Gen 16S ribosomal RNA (16S rRNA) memiliki daerah yang conserved
(lestari) sehingga tepat digunakan dalam Polymerase Chain Reaction dan analisis
sekuensing untuk menentukan taksonomi, filogeni dan keanekaragaman antar
spesies. Gen ini juga memiliki hypervariable region yang merupakan ciri khas tiap
mikroorganisme. Metode berbasis molekuler ini dinilai cepat dan akurat dalam
mengidentifikasi bakteri pathogen serta memiliki sejumlah keunggulan
dibandingkan metode mikrobiologi konvensional.
Analisis sekuensing gen 16S rRNA (16S ribosomal Ribonucleic acid/ asam
ribonukleat pengkode ribosum 16S) merupakan metode identifikasi berbasis
molekuler yang lebih cepat dengan tingkat sensitivitas dan spesifisitas yang tinggi.
Huruf S menyatakan “Svedberg” yaitu satuan ukuran ribosum. Gen 16S rRNA juga
sering disebut 16S rDNA (16S ribosomal deoxyribose nucleic acid). Namun istilah
16S rRNA lebih tepat menurut konsensus American Society for Microbiology
(ASM) (Rinanda, 2011).

28
Gen pengkode RNA ribosomal (rRNA) adalah gen yang paling lestari
(conserved). Porsi sekuens rRNA dari tiap organisme yang secara genetik
berkolerasi umumnya adalah sama. Daerah yang lestari ini juga yang menyebabkan
gen ini dapat digunakan sebagai primer universal yang digunakan dalam
Polymerase Chain Reaction (PCR) serta dapat ditentukan urutan nukleotidanya
melalui sekuensing. Penggunaan sekuens 16S rRNA dipelopori oleh Carl Woese,
yang juga menemukan klasifikasi 3 domain terbesar makhluk hidup, yaitu bakteri,
archaea dan eukaria (Rinanda, 2011).
Gen 16S dan 23S rRNA memiliki ukuran yang cukup untuk dianalisis. Gen
16S rRNA berukuran sekitar 1550 pasang basa dan sekitar 500 basa di bagian ujung
sekuens merupakan daerah yang disebut dengan hypervariable region. Daerah ini
merupakan bagian yang membedakan antar organisme. Primer yang digunakan
dalam Gen 16S rRNA adalah salah satu gen yang telah dikarakterisasi dengan baik
sehingga digunakan dalam identifikasi mikroorganisme. Sebagian besar prokariot
memiliki 3 jenis rRNA, yaitu 5S, 16S dan 23S. Sejak ditemukan pertama kali oleh
Woose sekuens 16S rDNA semakin banyak digunakan (Rinanda, 2011).
Primer yang digunakan dalam PCR adalah primer 16S rRNA yang bersifat
universal berukuran sekitar 1500pb, sehingga dapat mengamplifikasi daerah 16S
rRNA dari seluruh bakteri. Sekuens DNA terbentuk dari hasil pensejajaran
pembacaan primer reverse dan forward dan umumnya digunakan sebagai sekuens
consensus (consensus sequence). El Aila, dkk (2010) melakukan identifikasi S.
pneumonia dengan analisis sekuensing gen 16S rRNA dan memberikan hasil yang

29
lebih akurat (Rinanda, 2011). Aplikasi 16S rRNA pada mikrobiologi adalah sebagai
berikut:
1. Penandaan gen 16S rRNA dinyatakan sebagai baku standar identifikasi dan
klasifikasi spesies bakteri.
2. Rangkaian gen 16s rRNA memiliki variabel luas sehingga dapat memberikan
urutan tanda khas suatu spesies yang berguna untuk identifikasi suatu spesies
bakteri.
3. Pada mikrobiologi medis penandaan 16S rRNA merupakan alternatif metode
yang cepat dan murah untuk mengetahui fenotif suatu bakteri.
4. Proses penandaan ini juga mampu mengklasifikasikan suatu spesies bakteri
baru atau setidaknya mengetahui genus dari bakteri tersebut.
5. Teknik penandaan gen ini dapat digunakan untuk mendiskripsikan suatu spesies
baru yang tidak berhasil diidentifikasi pada metode kultur di laboratorium.
2.7.7. Penggunaan primer 16S rRNA pada meningitis bakterialis.
PCR berbasis 16S rRNA mengunakan universal bacterial primers yang
berguna mendeteksi bakteri pathogen dalam sampel cairan serebrospinal.
Penggunaan primer 3-spesies secara simultan dalam format PCR multiplex dan
seminested telah berhasil mengidentifikasi keberadaan Streptococcus pneumonia,
Haemophilus influenza dan Neisseria Meningitidis dalam 4 jam.
Sensitivitas, spesifisitas, nilai prediktif positif (NPP) dan nilai prediktif
negatif (NPN) secara keseluruhan dari PCR 16S rRNA, dimana kultur digunakan
sebagai baku emas adalah masing-masing 79,24%; 97,6%; 89,36% dan 94,88%

30
Keterbatasan kemampuan deteksi dari PCR 16S rRNA ditentukan oleh 1000 cfu/ml
E. coli dan 4000 cfu/ml S. pneumoniae.
Spesifisitas jangkauan PCR universal yang luas didapatkan meningkat
dengan analisa lebih lanjut terhadap produk PCR dengan metode yang berbeda
seperti restriction fragment length polymorphism (RFLP), penyelidikan atau
mengurutkan. Secara keseluruhan, PCR 16S rDNA merupakan temuan yang
menjanjikan sebagai sebuah uji skrining untuk diagnosis meningitis bakterialis
akut, khususnya di rumah sakit dimana hampir setiap pasien telah mendapatkan
antibiotik.
2.7.8. Strategi PCR
Universal primers (u3, ru8) memperkuat area 1000 bp spesifik dari 16S
rRNA hampir seluruh bakteri. Primers spesifik dapat digunakan untuk S.
penumoniae (STREP), H. influenzae (HI), dan N.meningitidis (NM), yang
merupakan 3 agen penyebab tersering meningitis bakterialis pada anak. Primers
diperkuat pada area spesifik yaitu dengan panjang area 1000 bp 16S rRNA yang
telah kuat dengan universal primers (tabel 2.1).
Streptococcus pneumoniae ATCC 49619, Haemophilus influenzae ATX
33391, dan Neisseria meningitidis ATCC 13077 rantai grup B digunakan sebagai
persiapan DNA standar. Kurang lebih 106cfu/ml dari masing-masing rantai DNA
bakteri telah diambil dan diekstraksi dan dipurifikasi dan telah dilakukan dengan
metode phenol-chloroform yang konvensional sebagaimana dijelaskan pada
Sambrook dkk.

31
Tabel 2.1 Oligonukleotida primers yang digunakan untuk identifikasi bakteri
pada meningitis dengan 16S rRNA
Kode Primer Urutan Primer
Ukuran
Produk
u3 5’ GTG CCT GCA GCC GCG GTA AT3’ 1000 bp
ru8 5’ AAG GAG GGG TGT GTA C 3’
STREP
(S.pneumoniae)
5’ GTA CAA CGA GTC GCA AGC 293 bp
HI
(H.influenzae)
5’ CCT AAG AAG AGC TCG AG 3’ 543 bp
NM
(N. meningitides)
5’ TGT TGG GCA ACC TGA TTG 3’ 710 bp
2.8 Pemeriksaan Pencitraan pada Meningitis
CT scan sering dilakukan sebelum punksi lumbal dalam diagnosis
meningitis. Hal ini untuk menyingkirkan adanya peningkatan tekanan intrakranial
yang dapat menimbulkan herniasi jika dilakukan punksi lumbal (Brouwer dkk.,
2012). Pencitraan pada meningitis (CT scan kepala atau MRI kepala) bukan
merupakan pemeriksaan rutin. Pencitraan hanya diindikasikan pada kondisi tertentu
atau kondisi spesifik dan hanya dilakukan jika pasien dalam kondisi stabil.
Pemeriksaan CT scan kepala bukan untuk diagnosis meningitis bakterialis tetapi
untuk menentukan adanya komplikasi meningitis bakterialis bila pengobatan tidak
memberikan respon yang adekuat atau untuk menyingkirkan kemungkinan
diagnosis yang lain seperti perdarahan intracranial (NICE, 2010; NSW, 2014).
Pada pasien dengan suspek meningitis, CT scan kepala hanya dilakukan jika
terdapat gejala neurologis fokal, penurunan kesadaran, adanya kejang yang terus

32
menerus, kejang parsial atau umum. Punksi lumbal merupakan indikasi kontra pada
pasien yang menunjukkan tanda-tanda efek massa atau herniasi pada pemeriksaan
CT scan kepala (Duval dkk., 2008).
2.9 Skor Meningitis dan Diagnosis Meningitis
Diagnosis yang cepat dan terapi yang tepat dapat menurunkan morbiditas
dan mortalitas akibat meningitis bakterialis. Namun diagnosis dapat terlambat
karena presentasi klinis yang tidak khas. Riwayat penyakit dan pemeriksaan fisik
saja tidak cukup untuk menegakkan diagnosis. Berbagai sumber pustaka berusaha
menjelaskan bagaimana membedakan antara meningitis bakterialis dari aseptik
meningitis (NSW, 2014).
Untuk menegakkan diagnosis pasti meningitis dan untuk menyingkirkan
meningitis, dokter harus melakukan punksi lumbal pada pasien yang diduga
meningitis atau yang menunjukkan gejala rangsang meningen (meningeal sign).
Punksi lumbal merupakan baku emas dalam diagnosis meningitis. Namun
seringkali hasil analisis CSS menunjukkan hasil yang tidak khas sehingga
menyulitkan dalam menegakkan diagnosis. Demikian juga hasil pemeriksaan
mikrobiologi, dimana pengecatan gram seringkali menunjukkan hasil yang negatif
dan pemeriksaan kultur kuman memiliki sensitivitas yang sangat rendah.
Pada pasien dengan hasil pengecatan gram atau kultur yang negatif,
diagnosis meningitis bakterialis akut seringkali sulit ditegakkan maupun
disingkirkan. Kombinasi gejala klinis dengan maupun tanpa hasil pemeriksaan
laboratorium telah dikembangkan menjadi suatu model yang memiliki akurasi

33
prediksi untuk membedakan meningitis bakterialis akut dan kemungkinan
penyebab lain terutama meningitis virus/aseptik. Namun sampai saat ini tidak ada
metode yang memiliki sensitivitas dan spesifisitas 100% untuk mengidentifikasi
meningitis bakterialis dan meningitis aseptik. Untuk menentukan diagnosis
meningitis pada anak seringkali digunakan kombinasi beberapa kriteria (Brouwer
dkk., 2012).
Dilema dalam diagnosis meningitis menjadi lebih sulit jika penderita sudah
mendapatkan terapi antibiotika sebelumnya sehingga menunjukkan hasil analisis
CSS lebih tidak spesifik dan hasil mikrobilogi tidak menemukan adanya
pertumbuhan kuman (Brouwer dkk., 2012).
Model prediksi diagnosis yang menggunakan kombinasi gejala klinis
dengan atau tanpa pemeriksaan laboratorium penunjang dapat digunakan
membedakan meningitis bakterialis akut dari penyebab non bakteri, namun model
ini tidak sepenuhnya dapat digunakan. Tabel 2.2. berikut ini adalah model prediksi
klinis untuk meningitis bakterialis (Brouwer dkk., 2012).

34
Tabel 2.2
Beberapa Model Prediksi Klinis untuk meningitis bakterialis
Studi& design Populasi aturan prediksi item skor Oostenbrink meningitis score
Original /prospektif, (n=286; v=74) Original/retrospektif (n=227) validasi/Prospektif (n=226)
Children aged 1 month–15 years anak usia 1 bln - 15 th
tidak ada menigitis bakteri bila nilai <8-5 pada 44 skala nilai (kisaran 0-44)
Durasi keluhan=1 poin per hari (maksimal 10), muntah=2, iritasi meningeal =7·5, cyanosis=6·5, petechiae or ecchymosis=4, teganggu kesadaran=8, CRP=0·5 points setiap peningkatan 10 mg/L, hitung PMN CSS =0–4*†, rasio glukosa CSS terhad darah=0·5 poin setiap penurunan 0·1*
Bacterial meningitis score
Nigrovic (o/r, n=456, v=240),56 Nigrovic (v/r, n=3295) 20 Dubos (v/r, n=198)57
anak dan dewasa muda usia 29 hari - 19 th (16th pada studi Dubos)
kemungkinan tidak meningitis bakterialis bila nilai 0 pada 6 skala nilai (kisaran 0-6)
Positif pada pewarnaan gram CSS=2, protein CSS 0·8 g/L =1, nilai absolut neutrofil perifer 10 000 cells per μL=1, kejang sebelum atau pada saat masuk rumah sakit =1, nilai absolut neutrofil CSS>1000 cells per μL=1
Spanos CSF prediction model
Spanos (o/r, n=422),43 McKinney (v/r, n=160),58 Hoen (v/r, n=500),59 Leblebicioglu (v/r, n=30)60
anak usia >1bln dan dewasa (pada studi McKinney, didefinisikan sebagai>17th
kemungkinan menigitis bila terdapat 1 karakteristik CSS
Kadar glukosa CSS <1·9 mmol/L,rasioglukosa CSS terhadap darah <0·23, CSF, kadar protein>2·2 g/L, leukosit CSS>2000/μL, neutrofil CSS>1180/μL
Hoen CSF prediction model
Hoen (o/r, n=500),59 Leblebicioglu (v/r, n=30),60 Baty (v/p, n=109)61
anak usia >1bl dan dewasa
kemungkinan tidak meningitis bakterialis bila skor <0·1
Formula risiko meningitis bakterialis =1/(1+e"L), dimana L=32·13×10"4×CSF hitung PMN (106/L)+2·365× protein CSS (g/L)+0·6143× glukosa darah (mmol/L)+0·2086×hitung seldarahputih (109/L)–11
Sumber: Brouwer dkk. Lancet. 2012; 380:1684-92.
2.9.1 Skor meningitis menurut “Oostenbrink”
Oostenbrink dkk. (2004) telah mengembangkan suatu model prediksi klinis
untuk menuntun dalam mengambil keputusan klinis berupa punksi lumbal dan
pemberian antibiotika empiris pada anak usia 29 hari sampai 15 tahun dengan
suspek meningitis bakterialis akut. Model ini dikenal dengan “Oostenbrink clinical

35
decision rule” dan merupakan salah satu cara untuk memprediksi risiko meningitis
bakterialis pada anak dengan tanda rangsang meningeal. Model Oostenbrink
melibatkan berbagai variabel dari riwayat penyakit, pemeriksaan fisik, dan
pengukuran kadar CRP serum. Lumbal punksi harus dilakukan pada anak yang
memiliki gejala gejala sesuai kriterianya dan memiliki total skor 8,5 atau lebih.
Pada uji validasi tidak seorangpun anak yang memiliki skor <9,5 menderita
meningitis bakterialis akut dan punksi lumbal dilakukan pada sekitar 35% anak
dengan tanda rangsang meningen, namun tidak ada satupun kasus meningitis
bakterialis yang diagnosisnya terlewatkan. Dengan nilai batas skor 9,5 atau lebih
dapat digunakan menentukan secara klinis meningitis bakterialis akut atau bukan.
Kriteria yang digunakan dapat dilihat pada tabel 2.3 berikut ini (Oostenbrink, 2002;
Dubos dkk., 2006; 2010).
Tabel 2.3
Kriteria klinis meningitis menurut Oostenbrink
Faktor risiko Skor/nilai Riwayat lamanya keluhan utama 1/ hari (maksimal 10) Riwayat muntah 2 Pemeriksaan fisik:
Sianosis 6,5 Kesadaran menurun 8,0 Iritasi meningen 7,5 Petechie 4,0
Kadar serum CRP (mg/dL) atau mg/L <5.0 (50) 0 5,0-9,9 (50-99) 0,5 10,0-14,9 (100-149) 1 15,0-19,9 (150-199) 1,5 >20 (200) 2
Total skor = *Apabila total skor >8 harus dilakukan tindakan punksi lumbal
Sumber: Oostenbrink dkk.Arch Pediatr Adolesc Med. 2002; 156:1189-94.

36
2.9.2 Bacterial Meningitis Score (BMS)
Model prediksi lain telah dikembangkan oleh Nigrovic dkk. Kriteria
meningitis yang dikembangkan oleh Nigrovic ini dikenal dengan Nigrovic
Bacterial Meningitis Score (Nigrovic BMS). Komponen dalam kriteria Nigrovic
BMS meliputi: pengecatan gram CSS yang positif, adanya kejang yang
berhubungan dengan gejala klinis, hitung neutrophil pada darah tepi ≥10,000
sel/μL, hitung neutrophil pada CSS ≥1,000 sel/μL, dan kadar protein CSS ≥80
mg/dL. Apabila tidak ada satupun kriteria terpenuhi, maka diduga pasien memiliki
risiko rendah menderita meningitis bakterialis. Kriteria skor meningitis bakterialis
oleh Nigrovic ini telah dilakukan validasi dan hasilnya menunjukkan sensitivitas
98.3% (95% IK: 94,2-99,8), spesifisitas 61,5% (95% IK:59,7-63,3) dengan
negative predictive value (NPV) sebesar 99,9% (95%IK: 99,6-100); Nilai rasio
LLH positif 2,56 dan rasio LLH negatif 0,03 (Nigrovic dkk., 2007; 2012)
Meskipun studi lain telah memberikan konfirmasi bahwa Nigrovic BMS
sangat berguna dibandingkan model lain, namun untuk aplikasi sistim scoring ini
tidak selalu sesuai. Sebuah studi retrospektif yang menggunakan skoring Nigrovic
pada 21 anak usia 0-15 tahun dengan suspek meningitis bakterialis, mendapatkan 5
orang yang tidak memenuhi kriteria dan dinyatakan berisiko rendah dan akhirnya
tidak mendapatkan terapi antibiotika. Peneliti ini akhirnya mengembangkan scoring
baru dengan menambahkan kadar CRP serum < 20 mg/L, kadar glukosa CSS> 2,89
mmol/L dan kadar protein CSS < 1 g/L. Nilai skor 0 dapat membedakan meningitis
virus dari meningitis bakterialis akut pada 54 dari 70 anak dengan akurasi 100%
dan spesifisitas 100% (Brouwer dkk., 2012)

37
Walaupun Nigrovic BMS dapat mengidentifikasi meningitis bakterialis
dengan sensitivitas yang sangat tinggi (99,6%), masih ada kasus yang tidak
terdeteksi sehingga seorang klinisi harus tetap menyadari bahwa tidak ada cara yang
dapat secara tepat menyingkirkan meningitis bakterialis. Demikian juga bahwa
meningitis bakterialis dapat terjadi tanpa adanya pleositosis dalam CSS walaupun
sangat jarang. Dengan demikian sampai saat ini diagnosis meningitis masih cukup
sulit ditegakkan sedangkan perlu tatalaksana yang segera dan tepat.
Sebuah penelitian menggunakan skor meningitis yang modifikasi dari
Nigrovic telah dilakukan di Gangnam Severance Hospital oleh Kwon dkk. Kriteria
skor meningitis yang digunakan dapat dilihat pada tabel 2.4. Kesimpulan penelitian
ini adalah bahwa BMS dapat digunakan untuk mengambil keputusan klinis pada
anak dengan pleositosis CSS dan masih perlu dilakukan evaluasi apakah BMS dapat
digunakan dengan aman untuk menurunkan angka rawat inap dan penggunaan
antibiotika pada anak dengan meningitis aseptik. Sangat berbeda dengan jika BMS
>2 sangat mendukung kecurigaan meningitis bakterialis dengan sensitivitas dan
spesifisitas yang tinggi. Meningitis bakterialis dapat terjadi pada pasien tanpa CSS
pleositosis dan BMS menunjukkan risiko rendah, oleh karena itu seharusnya BMS
digunakan secara berhati hati terutama pada bayi usia di bawah 2 bulan karena BMS
kurang akurat. Masih diperlukan penilaian klinis, epidemiologi dan pemeriksaan
laboratorium yang lain (Kwon dkk., 2013).

38
Tabel 2.4
Komponen Bacterial Meningitis Score (BMS)
Prediktor BMS Poin
Ada Tidak
Pengecatan gram CSS positif 2 0
Hitung netrofil absolute > 1000 sel/mm3 1 0
Protein CSS > 80 mg/Dl 1 0
Hitung netrofil absolut dalam darah tepi>10.000 sel/mm3 1 0
Kejang saat/sebelum timbul gejala 1 0
Kriteria:
• Total poin 0 = sangat mendukung diagnosis meningitis aseptic
• Total poin 1 = cendrung meningitis aseptik
• Total poin 2-6 = lebih mendekati diagnosis meningitis bakterialis
Sumber : Kwon dkk. J Korean Child Neurol Soc. 2012; 21(2): 46-520
2.9.3 Meningitest
Metode untuk mengambil keputusan klinis untuk pasien anak yang
berdasarkan pada BMS telah dievaluasi dan kemudian dimodifikasi dan disebut
Meningitest. Komponen yang digunakan untuk menilai berbeda dengan BMS,
dimana hitung jenis netrofil dalam serum dan CSS dikeluarkan dari sistem skoring
dan biomarker procalcitonin (PCT) dimasukkan dalam kriteria. Metode ini
digunakan untuk merekomendasi penggunaan terapi antibiotik dan rawat inap untuk
anak yang menunjukkan gejala meningitis dengan gejala minimal salah satu dari
komponen kriteria. Tabel 2.5 menunjukkan komponen kriteria Meningitest.

39
Tabel 2.5
Komponen kriteria Meningitest
Kejang
Penampakan klinis toksik
Purpura
Pengecatan gram CSS positif
Kadar procalcitonin serum ≥0.5 ng/mL
Kadar protein CSS ≥50 mg/dL
Sumber: Quenot dkk. Annal of Intensive care. 2013,3:21
Meningitest menunjukkan hasil validasi eksternal yang baik dengan sensitivitas
100% dan spesifisitas 37-62%. Namun saat ini, pemeriksaan procalcitonin tidak
selalu tersedia di sebagian besar rumah sakit, tetapi dapat bermanfaat di kemudian
hari untuk menghindari rawat inap yang tidak perlu dan mengurangi penggunaan
terapi antibiotika yang tidak perlu (Quenot dkk., 2013).
2.10 Biomarker Diagnostik
Berbagai penelitian tentang penggunaan marker diagnostik pada meningitis
bakterialis akut telah banyak dilakukan. Penelitian penelitian sebelumnya terutama
terfokus terhadap bagaimana membedakan meningitis bakterialis dari meningitis
virus. Beberapa biomarker yang telah diteliti sebagai marker untuk meningitis
bakterialis adalah HBP, kadar kortisol, soluble triggering receptor expressed on
myeloid 1, interleukin 6 (IL-6), interleukin 12 (IL-12), interleukin 1β (IL-1β), tumor
necrosis factor α (TNF-α). Sebagian besar studi ini hanya melibatkan pasien dalam

40
jumlah sedikit (<40 pasien) sehingga hasilnya kurang dapat digeneralisasi (Quenot
dkk., 2013).
Suatu biomarker idealnya memiliki sensitivitas 100% dan juga memiliki
spesifisitas yang dapat diterima, walaupun jika digunakan tersendiri. Biomarker
yang telah ada seperti procalcitonin (PCT), C-reactive protein (CRP), interferon
gamma (IFN-γ) dan sebagainya, nampaknya tidak memiliki sensitivitas dan
spesifisitas yang cukup tinggi jika digunakan sebagai dasar memutuskan terapi
karena risiko hasil test yang negatif palsu (Quenot dkk., 2013).
2.10.1 Laktat Dehidrogenase
Laktat dalam CSS diproduksi oleh bakteri anaerob sebagai hasil
metabolismenya dan tidak dipengaruhi oleh kadar laktat dalam darah. Kadar laktat
di dalam CSS merupakan potensial biomarker untuk meningitis bakterialis pada
anak (Sakushima dkk., 2011).
Anak dengan meningitis bakterialis memiliki kadar laktat CSS lebih tinggi
dibandingkan dengan anak dengan meningitis aseptik. Pada nilai titik potong 3,0
mmol/L memiliki sensitivitas 95% pada 95% interval kepercayaan (IK) 83-99%
dan spesifisitas 94% (IK 90-96%); NPV 99,3% (IK 97,7-99,9%) untuk meningitis
bakterialis (Filho dkk., 2014).
Peningkatan kadar laktat dalam CSS dapat digunakan untuk membedakan
meningitis bakterialis dari non meningitis bakterialis pada pasien yang belum
mendapatkan terapi antibiotika. Salah satu penelitian mendapatkan bahwa kadar
laktat dalam CSS lebih dari 4.2 mmol/L memiliki korelasi yang positif dengan

41
meningitis bakterialis dengan sensitivitas 96% dan spesifisitas 100% (Devlin dan
Byars, 2011).
Sebuah meta-analisis yang melibatkan 32 penelitian telah dilakukan.
Hasilnya adalah kadar laktat CSS memiliki sensitivitas 93% (95%IK: 0,89-0.96);
spesifisitas 96% ( 95%IK: 0,93-0,98); rasio LLH positif 22,9 (95%IK:12,6-41,9);
dan rasio LLH negatif 0,07 (95%IK:0,05-0,12); rasio odds diagnosis 313
(95%IK:141-698). Pada pasien yang telah mendapat antibiotika sebelum punksi
lumbal, kadar laktat CSS memiliki sensitivitas yang lebih rendah dibandingkan
dengan yang belum mendapat antibiotika yaitu 49% dan 98% secara berurutan.
Kadar laktat yang tinggi sekitar 35 mg/dL memiliki sensitivitas dan spesifisitas
yang lebih tinggi dibandingkan dengan kadar sekitar 27 mg/dL (Sakushima dkk.,
2011).
Peningkatan kadar laktat CSS dinyatakan spesifik untuk meningitis, namun
peningkatan kadar laktat CSS juga dapat disebabkan oleh iskemia serebri dan
metabolism anaerob akibat inflamasi sehingga kadar laktat tidak ditambahkan
untuk mendiagnosis meningitis bakterialis. Kadar laktat CSS juga meningkat pada
pasien dengan penyakit saraf yang berat seperti stroke dan trauma kepala sehingga
hal ini merupakan keterbatasannya. Karena rendahnya spesifisitas peningkatan
kadar laktat maka saat ini pengukuran kadar laktat tidak direkomendasikan dalam
manajemen meningitis yang khas (Devlin dan Byars, 2011).
2.10.2 Komponen komplemen B (CB) dan komplemen 3 (C3)
Dalam membedakan antara meningitis bakterialis dan meningitis virus,
kadar complement component B dalam CSS memiliki sensitifitas dan spesifisitas

42
100% pada orang dewasa, kadar complement component 3 memiliki sensitivitas
100% dan spesifisitas 95% dan kadar HBP memiliki sensitivitas 100% dan
spesifisitas 99,2% (Brouwer dkk., 2012).
2.10.3 C-Reactive Protein (CRP)
C-Reactive Protein merupakan reaktan fase akut yang dibuat dalam hepar
dan disekresi dalam 6 jam setelah reaksi inflamasi akut. Konsentrasi CRP >40mg/L
memiliki spesifisitas 100% dan sensitivitas 93% untuk mengidentifikasi meningitis
bakterialis (Brouwer dkk., 2012). Pengukuran kadar CRP serum pada meningitis
bakterialis memiliki sensitivitas 69%-99% dan spesifisitas 28%-99% dan nilai OR
untuk meningitis bakterialis adalah 150 (95%CI, 44-509). CRP serum bermanfaat
untuk membedakan meningitis bakterialis jika hasil pengecatan gram negatif,
dengan sensitivitas 96% dan spesifisitas 93% dan NPV 99%. Nilai CRP yang
normal memiliki nilai prediksi negatif yang tinggi dalam diagnosis meningitis
bakterialis (Tunkel dkk., 2004).
2.10.4 Procalcitonin (PCT)
Procalcitonin merupakan suatu potent partial agonist (50-60% dari efikasi
CGRP) dari reseptor CGRP1 dan inhibited CGRP-dependent cAMP responses dan
weak partial agonist activity pada reseptor AMY1 (Gude, 2012).
Penelitian dari Alkholi, dkk (2011) menunjukkan kadar PCT lebih tinggi
secara bermakna pada pasien anak dengan meningitis bakterialis (rerata 24,8 ng/ml)
dibandingkan dengan non meningitis bakterialis (rerata 0,3 ng/ml) dengan nilai p
<0,001. Kadar PCT pada kelompok meningitis bakterialis menurun setelah 3 hari
post terapi antibiotika, tetapi masih lebih tinggi dibandingkan kelompok non

43
meningitis bakterialis (rerata 10,5 ng/ml). Kadar PCT serum >2 ng/ml memiliki
sensitivitas 100%, spesifisitas 66%, PPV 68% dan NPV100% untuk diagnosis
meningitis bakterialis (Alkholi dkk., 2011).
Untuk anak dengan suspek meningitis, kadar procalcitonin (PCT) >0,5
ng/mL dapat digunakan untuk membantu dalam menentukan diagnosis meningitis
bakterialis tetapi dapat terjadi negatif palsu sehingga lebih dianjurkan
menggunakan Bacterial Meningitis Score (BMS) atau Meningitest® dibandingkan
menggunakan PCT secara tunggal. Sebaiknya menggunakan PCT dalam suatu
“clinical role” untuk membantu dalam menentukan keputusan. Procalcitonin (PCT)
dapat membantu memprediksi infeksi, walaupun data data yang ada tidak
mencukupi untuk mengambil keputusan memulai pemberian antibiotika (Quenot
dkk., 2013).
Skor Meningitest® (European patent EP1977244) memasukkan PCT dalam
kriteria untuk mendiagnosis suspek meningitis. Meningitest® merekomendasikan
memulai terapi antibiotika apabila minimal memenuhi 1 dari kriteria berikut:
kejang, penampakan toksik, purpura, PCT >0,5ng/mL, pengecatan gram positif atau
kadar protein dalam CSS >0,5 g/L. Kadar serum PCT>0,5 ng/mL secara tunggal
memiliki sensitivitas dan spesifisitas hampir sama dengan BMS, sedangkan
kombinasi CRP dengan kadar protein CSS memiliki sensitivitas dan spesifisitas
yang lebih rendah (Quenot dkk., 2013).
Meta-analisis dari enam studi retrospektif yang melibatkan 198 anak
meningitis bakterialis menunjukkan bahwa peningkatan kadar PCT serum >0,5
µg/L dan kadar CRP>20 mg/L dapat diasosiasikan dengan meningitis bakterialis

44
akut dengan rasio odds 434 terhadap peningkatan PCT dan 9,9 terhadap
peningkatan CRP. Walaupun demikian apakah pemeriksaan tambahan PCT dan
CRP serum memberi nilai tambah masih belum jelas (Brouwer dkk., 2012). Kadar
PCT serum pada meningitis telah diteliti pada anak anak. Pada sebuah penelitian,
kadar PCT lebih dari 0,5 μg/L memiliki sensitivitas 94% untuk meningitis
bakterialis pada anak, dengan spesifisitas 100% (Quenot dkk., 2013).
2.11 Peranan Sitokin Pada Meningitis bakterialis
Sitokin merupakan molekul molekul yang terlibat dalam modulasi proses
imun dan inflamasi. Tumor necrosis factor-alpha (TNF-α) merupakan suatu
aktivator netrofil dan mediator yang berperan pada kemotaksis dan degranulasi.
Interleukin-6 menstimulasi proliferasi limfosit B yang telah berdiferensiasi menjadi
sel yang menghasilkan antibodi. Interleukin-8 (IL-8) bekerja sebagai kemoatraktan
terhadap netrofil di tempat inflamasi. Kadar TNF-α dan IL-8 pada pasien meningitis
bakterialis telah dilaporkan pada berbagai penelitian. Rerata kadar TNF-α, IL-6, IL-
8 dalam CSS secara bermakna lebih tinggi pada meningitis bakterialis
dibandingkan dengan meningitis virus. Peningkatan kadar TNF-α, IL-6 dan IL-8
dalam CSS anak dengan meningitis bakterialis, menunjukkan bahwa sitokin
tersebut berperan pada patogenesis meningitis bakterialis (Prasad dkk., 2014).
Interleukin-12 (terminologi dari IL-12p70 dan biasa disebut IL-12)
merupakan suatu sitokin imunoregulator yang penting yang terutama dihasilkan
oleh Antigen Precenting Cells (APC). Ekspresi IL-12 selama terjadi infeksi adalah
mengatur respon alamiah dan menentukan tipe respon imun adaptif. Interleukin-12

45
menginduksi produksi interferon gamma (INF-γ) dan sebagai trigger CD4+ sel T
untuk berdiferensiasi menjadi sel T-helper tipe-1 (Th1). Interleukin-12 dapat
memainkan peranan yang vital dalam proses penyembuhan penyakit seperti infeksi
virus, bakteri dan kanker. Struktur heterodimerik yang unik dari IL-12 dimana IL-
12 bersama sama dengan anggota familinya yaitu IL-23, IL-27 dan IL-35 akhir
akhir ini lebih menarik perhatian untuk dipahami mekanisme yang mengatur fungsi
IL-12 (Hamza dkk., 2010).
Interleukin-12 merupakan suatu sitokin yang heterodimeric yang
diproduksi oleh sebagian besar sel fagositik sebagai respon terhadap bakteri, produk
bakteri dan parasit intraseluler dan beberapa sel limfosit B. Pada infeksi bakteri IL-
12 berperan sebagai penginduksi yang kuat terhadap respon imun Th-1 dan
mediator penting antara imunitas alamiah dan didapat (Hamza dkk., 2010).
2.12 Respon imun terhadap infeksi
Selama infeksi atau akibat inflamasi yang berat, molekul mikroba atau
molekul yang memberikan sinyal bahaya (endogenous danger signal) dan mediator
dari host mampu memodulasi homeostasis dari host. Respon inflamasi lokal
maupun sistemik dapat bermanfaat atau merugikan. Respon yang bermanfaat
menghasilkan terkendalinya proses infeksi. Respon inflamasi yang merugikan
dapat terjadi dengan mekanisme sebagai berikut (Conquy dan Cavaillon, 2009):
a. Perjuangan melawan agen infeksi dapat terlalu berlebihan dan
menyebabkan disfungsi organ.
b. Respon anti-inflamasi yang bertujuan untuk mengurangi proses inflamasi
dapat mengubah status imun.

46
c. Terjadi perubahan keseimbangan antara status prokoagulan dan
antikoagulan dari host.
Bone dkk. (1996) pertama kali memperkenalkan konsep systemic inflamatory
response syndrome (SIRS). Berdasarkan teorinya dijelaskan bahwa, respon imun
terhadap infeksi atau trauma mulai pada lokasi infeksi dan berhubungan dengan
produksi mediator pro-inflamasi dan anti-inflamasi, yang terutama dihasilkan oleh
sel sel respon imun alamiah (innate) seperti netrofil. Respon imun ini bertujuan
untuk menjaga homeostasis dan sebagai respon awal reaksi inflamasi yang
fisiologis. Telah diketahui pada infeksi yang persisten atau adanya infeksi tambahan
dari lokasi lain seperti pada saluran pernafasan dapat meningkatkan kelainan respon
imun sistemik yang dapat memicu sepsis dan kegagalan multiorgan. Berdasarkan
konsep tersebut dapat terjadi kelainan sistem imun yang dapat menimbulkan
sindrom imun paralisis (kekacauan respon imun), yang dapat meningkatkan
komplikasi dan mortalitas (Binkowska dkk., 2015).
Apoptosis dari sel sel sistem imun (makrofag, netrofil, limfosit) memainkan
peranan yang sangat penting yang dapat membangkitkan imunosupresi dan
merupakan elemen penting dalam respon terhadap infeksi. Respon patologi dari
SIRS dan CARS (compensatory anti-inflammatory response syndrome) pada
proses terjadinya kegagalan multiorgan (MODS) adalah akibat kurangnya
mekanisme koordinasi atau kerjasama antar sel sistem imun yang terlibat dalam
merusak organ. Telah ditunjukkan hubungan antara beratnya SIRS/CARS/MODS
dan sel yang mengalami apoptosis, melibatkan respon pro-inflamasi dan anti-
inflamasi terutama netrofil, yang meningkatkan produksi sitokin dan radikal bebas.

47
Respon imun yang tidak terkontrol terhadap infeksi dapat memicu self
destruction jaringan dan disfungsi organ yang jauh dari lokasi infeksi. Konsep baru
dikembangkan oleh Murphy dkk. (2004) bahwa CARS terjadi secara paralel
(bersamaan) dengan respon kompensasi SIRS, terutama diasosiasikan dengan
peningkatan produksi sitokin antiinflamasi IL-4 dan IL-10 sehingga berkembang
menjadi imunosupresi (Binkowska dkk., 2015).
2.13 Mekanisme brain-mediated systemic anti-inflamatory syndrome yang
mengakibatkan depresi sistem imun.
Untuk mencegah aksi sitokin proinflamasi yang berlebihan dan merusak
setelah efek awal yang bermanfaat, sistem imun dapat mengeluarkan beberapa
mediator anti-inflamasi, diantaranya interleukin 10 (IL-10), interleukin-1 receptor
antagonist (IL-1ra) dan tumor necrosis factor receptors, yang selanjutnya
mengawali suatu compensatory anti-inflammatory response syndrome (CARS).
Secara in vivo terdapat keseimbangan antara respon pro-inflamasi dan anti-
inflamasi yang juga dikontrol oleh sistem saraf pusat (SSP). Sitokin pro-inflamasi
menstimulasi aksis hypothalamic-pituitary-adrenal (HPA) dan memicu aktivitas
sistem saraf simpatik. Mediator dari jalur neuroimun ini dapat juga menekan fungsi
sel imun untuk mengontrol inflamasi sistemik.
Jalur inhibisi sistem imun SSP dapat diaktivasi di luar inflamasi sistemik.
Hal ini dapat dihasilkan dari produksi sitokin di dalam otak setelah infeksi, trauma,
atau iskemia atau dalam respon terhadap berbagai stress atau jika terjadi iritasi pada
batang otak. Teori brain mediated immunodepression dapat menjelaskan hal ini.

48
Inaktivasi monosit dengan menurunkan kemampuan presentasi antigen dan
menekan sekresi sitokin pro-inflamasi meningkatkan risiko komplikasi infeksi.
Sitokin pada otak dan stres dapat membangun imuno-depresi sistemik pada tingkat
monosit. Dalam hal ini katekolamin terinduksi untuk mengeluarkan sitokin anti-
inflamasi yang kuat (IL-10) yang merupakan penemuan baru dalam mekanisme
brain mediated monocyte deactivation, disamping aksi imunosupresif oleh
glukokortikoid yang telah diketahui dengan baik sebelumnya. Neuropeptida yang
lain seperti α-melanocyte-stimulating hormone dan β-endorphin yang dapat
dihasilkan dalam situasi stres juga dapat menghambat efek pada sel sel imun.
Infeksi lokal maupun trauma steril menginduksi respon inflamasi lokal.
Pengeluaran mediator pro-inflamasi seperti TNF- α, IL -1β, IL-6 mengaktifasi
kaskade inflamasi yang memperbaiki penyembuhan pada luka dan pertahanan anti
mikroba. Aktivasi sistem imun yang luar biasa dapat menimbulkan sindrom respon
inflamasi sistemik (SIRS) dan syok septik. Untuk mengontrol potensi respon
inflamasi yang berbahaya, sebaliknya sistem imun mengeluarkan beberapa
mediator anti inflamasi diantaranya IL-10, IL-1ra dan soluble TNF receptors, yang
menghambat produksi (IL-10) menetralisasi soluble TNF receptors 1 dan 2 atau
berkompetisi secara antagonis dengan mediator pro inflamasi (IL-1ra). TNF- α, IL
-1β dan prostaglandin merupakan induser kuat dari CARS.
Keseimbangan antara respon pro-inflamasi dan anti-inflamasi dikontrol oleh
brain-dependent central mechanisms. Komunikasi dua arah antara sistem imun
dengan SSP dibuktikan dengan peningkatan sekresi berbagai mediator oleh kelenjar
pituitary dan adrenal yang dinduksi oleh inflamasi sistemik atau infeksi. TNF-α, IL-

49
1β, IL-6 merupakan imun aktivator dari hypothalamic pituitary adrenal axis (HPA)
yang menghasilkan sekresi glukokortikoid. Peranan penting aksis HPA adalah
mencegah inflamasi yang berlebihan. Mekanisme umpan balik negatif diperkuat
oleh induksi berlebihan dari IL-10 yang dicetuskan oleh glukokortikoid sebagai
respon terhadap inflamasi trauma. Aktivasi aksis HPA, sitokin pro inflamasi dapat
mempengaruhi aktivitas sistem saraf simpatetik (SNS).
Katekolamin meningkatkan pengeluaran sitokin anti inflamasi IL-10 setelah
stimulasi endotoksin pada sel mononuclear darah perifer dan sebaliknya produksi
TNF-α akan menurun. Mekanisme ini merupakan jalur alternatif dimana SSP
bekerja sama dengan fungsi sistem imun dalam menjaga keseimbangan antara SIRS
dan CARS untuk mempertahankan homeostasis.
Neuropeptide lain seperti α melanocyte stimulating hormone (MSH) dan β
endorphin dapat menghambat efek inhibisi sel imun dan dapat mengeluarkan situasi
stress. Dengan demikian dapat disimpulkan bahwa sitokin otak dapat menjadi
pencetus respon anti inflamatori sistemik tanpa didahului inflamasi sistemik.
Akibatnya terjadi depresi inflamasi dan sistem imun seluler yang tidak adekuat dan
meningkatkan resiko infeksi.
Jalur neuro imun yang paling signifikan adalah aksis HPA dan sympathetic
nervous system (SNS), yang sangat penting dalam mengontrol respon imun
sistemik, yang memiliki peran protektif melawan inflamasi sistemik yang
berlebihan. Interleukin (IL-10) merupakan kunci mediator yang disebut sentral
CARS.
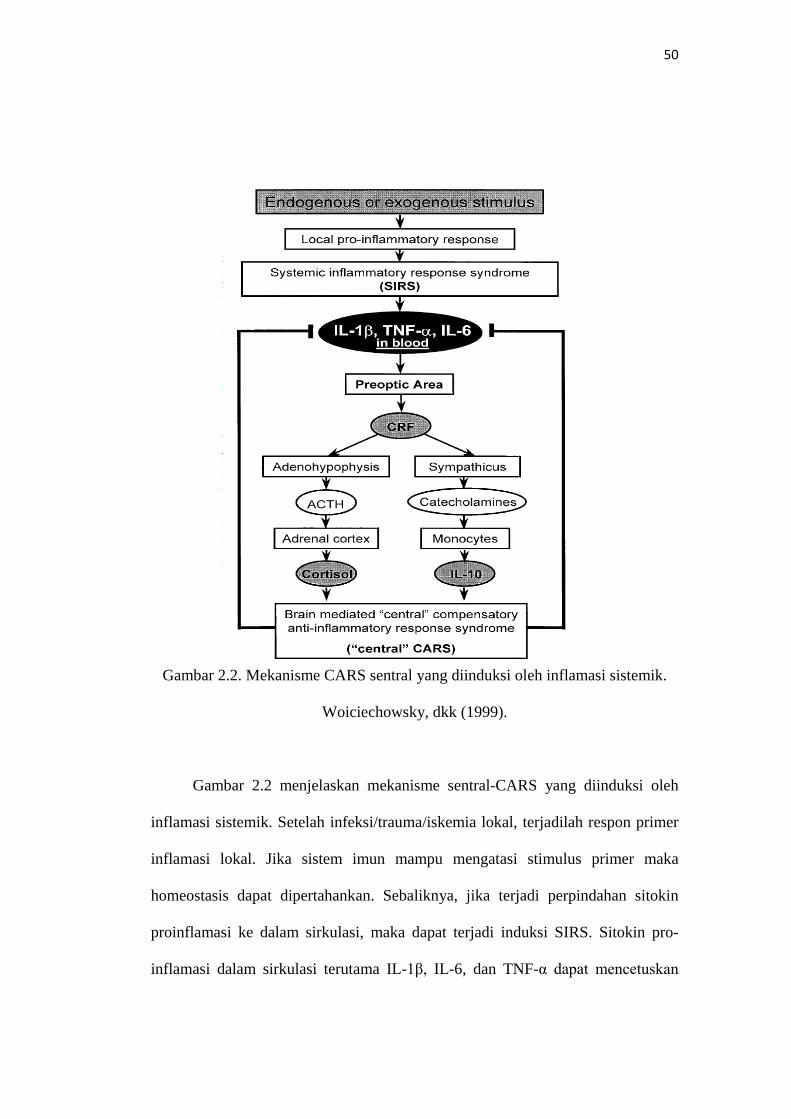
50
Gambar 2.2. Mekanisme CARS sentral yang diinduksi oleh inflamasi sistemik.
Woiciechowsky, dkk (1999).
Gambar 2.2 menjelaskan mekanisme sentral-CARS yang diinduksi oleh
inflamasi sistemik. Setelah infeksi/trauma/iskemia lokal, terjadilah respon primer
inflamasi lokal. Jika sistem imun mampu mengatasi stimulus primer maka
homeostasis dapat dipertahankan. Sebaliknya, jika terjadi perpindahan sitokin
proinflamasi ke dalam sirkulasi, maka dapat terjadi induksi SIRS. Sitokin pro-
inflamasi dalam sirkulasi terutama IL-1β, IL-6, dan TNF-α dapat mencetuskan

51
pengeluaran corticotropin –releasing factor (CRF) pada area preoptic, jika terjadi
kerusakan sawar darah otak. Hal ini dapat menstimulasi adenohipofisis/korteks
adrenal dan sympathetic nervous system (SNS) untuk menghasilkan ACTH/kortisol
dan katekolamin yng mencetuskan sekresi IL-10. Kedua mediator dapat mengawali
suatu CARS untuk meredam inflamasi sistem imun dan mempertahankan
homeostasis.
Gambar 2.3 Model hipotesis pro-inflammatory (SIRS) dan anti-inflammatory
response (CARS) terhadap trauma dan infeksi, yang dapat
mencetuskan kegagalan multiorgan dikutip dari: Binkowska, dkk
(2015).

52
2.14 Reflek inflamasi pada infeksi SSP
Inflamasi adalah respon lokal, protektif terhadap invasi mikroba atau
cedera. Apabila inflamasi menyebar ke dalam aliran darah, seperti yang terjadi
dalam sindrom syok septik, sepsis, meningitis dan trauma berat, respon inflamasi
dapat menjadi lebih berbahaya daripada stimulus pencetus aslinya. Homeostasis
dapat dikembalikan ketika proses inflamasi dibatasi oleh respon anti inflamasi
yang berlebihan, cepat, reversibel, terlokalisir, adaptif terhadap perubahan dalam
sistem saraf.
Pengetahuan terkini telah mengidentifikasi jalur saraf dasar yang secara
reflek memonitor dan menyesuaikan respon inflamasi. Stimulus inflamasi
mengaktifkan jalur sensoris, yang meneruskan informasi ke hipotalamus. Stimulus
inflamasi mengaktifkan respons anti inflamasi yang cepat dan otomatis untuk
mencegah kebocoran produk inflamasi ke dalam sirkulasi. Kontrol neural dari
inflamasi akut bersifat refleksif, saling berhubungan secara langsung dan
terkendali, terutama pada mekanisme anti inflamasi kolinergik yang menghambat
aktivasi makrofag dan pelepasan sitokin.
Mekanisme kontra-regulasi yang sangat terjaga pada kondisi normal, dapat
membatasi respon inflamasi akut dan mencegah penyebaran mediator inflamasi
ke dalam aliran darah. Sel imun yang kompeten teraktivasi melepaskan fragmen
reseptor yang mengikat TNF-α dan menetralkan aksi inflamasi yang berpotensi
toksik. Sitokin anti inflamasi, seperti IL-10 dan transforming growth factor β

53
(TGF-β), secara spesifik menghambat pelepasan TNF-α dan mediator
proinflamasi lainnya.
Glukokortikoid adrenal, adrenalin, α-melanocyte stimulating hormone (α-
MSH) dan hormon stres klasik lainnya menghambat sintesis sitokin dan transduksi
sinyal intraselular. Hilangnya mekanisme anti inflamasi endogen mengubah
respon inflamasi yang biasanya protektif dan mampu membatasi diri sendiri
menjadi respon berlebihan yang berpotensi merusak. Sebagai contoh, neuron di
SSP dapat mensintesis dan mengekspresikan TNF-α dan IL-1. Sitokin ini dapat
berpartisipasi dalam komunikasi neuronal. Komunikasi ini bersifat dua arah,
karena sitokin dapat mengaktifkan pelepasan glukokortikoid dari hipotalamus-
pituitari dan selanjutnya glukokortikoid menekan sintesis sitokin lebih lanjut.
Selain itu, sel sistem imun tubuh dapat memproduksi neuropeptida termasuk
endorfin, asetilkolin dan neurotransmiter lainnya.
Jalur anti inflamasi kolinergik merupakan mekanisme dasar yang
meregulasi inflamasi. Mekanisme saraf yang menghambat aktivasi makrofag
melalui arus keluar/outflow parasimpatis disebut jalur anti inflamasi kolinergik.
Asetilkolin (Ach) adalah neurotransmitter parasimpatik utama. Makrofag yang
terpapar pada asetilkolin dinonaktifkan secara efektif. Jalur anti inflamasi
kolinergik dapat menghambat inflamasi lokal secara spesifik.
Aktivitas saraf eferen pada nervus vagus berhubungan dengan pengeluaran
asetilkolin dalam sistem organ retikuloendotelial, yaitu hati, jantung, lien dan
traktus gastrointestinal. Asetilkolin berinteraksi dengan α-bungarotoksin-sensitive

54
nicotinic receptors (ACh receptor) pada jaringan makrofag yang menghambat
pengeluaran TNF, IL-1, high mobility group box 1 (HMGB1) dan sitokin lain.
2.15 Granul Protein dalam Netrofil sebagai Modulator Respon Imun.
Sel polimorfonuklear (PMN) merupakan tipe utama dari sel leukosit di
dalam darah tepi, sekitar 40-70% leukosit terdapat dalam kondisi normal. PMN
mempunyai peranan penting dalam pertahanan awal host melawan masuknya
mikroorganisme. Perekrutan sel PMN dari aliran darah ke tempat infeksi
melibatkan mediator mediator inflamasi, ikatan adhesi molekul pada endotel
pembuluh darah dan migrasinya melewati pertahanan endotel (Soehnlein, 2008).
Ketika sel PMN mencapai fokus infeksi, sel tersebut teraktivasi penuh dan
dapat melawan infeksi melalui perantara sekresi oksigen reaktif, peptida
antimikroba dan enzim degradatif (Soehnlein, 2008).
Netrofil merupakan keluarga granulosit dari leukosit termasuk juga
eosinofil dan basofil. Diberikan terminologi granulosit karena adanya sekelompok
kecil preformed protein yang menghiasi sitoplasma (granula) yang dapat
membantu dalam mendefinisikan morfologinya secara histologi. Beberapa tipe
granula yang berperan dalam netrofil adalah: granula sekretorik, granula primer,
granula sekunder dan granula tertier (Fox dkk., 2010).
Granula protein dari PMN merupakan kunci respon imun yang diinisiasi
oleh PMN (Chertov dkk., 1997). Subset PMN diklasifikasikan menjadi granula
primer, sekunder dan tersier sesuai dengan vesikel sekretorik. Hubungan di antara
kelompok granula dapat digunakan sebagai analisis marker protein (Sohnlein,

55
2008). Sekitar 300 protein yang berbeda tersimpan di dalam granula PMN
(Lominadze dkk., 2005), yang akan dikeluarkan ke sekitarnya, bekerjasama
dengan membran sel atau masih melekat pada membran sel selama mobilisasi
granula. Selama perjalanannya dari aliran darah ke lokasi inflamasi, PMN
mengeluarkan granulanya sesuai dengan hirarkinya. Hal ini dapat dilihat pada
gambar 2.5 (Soehnlein, 2008).
Ciri khas dari vesikel sekretorik adalah pengeluaran granulanya yang
sangat cepat jika terjadi kontak antara PMN dan sel endotel. Granula tersier
dimobilisasi jika PMN bertransmigrasi dan granula sekunder dan primer bebas
keluar pada tempat atau lokasi inflamasi. Protein di dalam granula merupakan
suatu yang bebas jika diperlukan PMN untuk membunuh organisme. Selanjutnya
protein-protein ini mengendap/terdeposit di dalam ruang ekstravaskuler dan
secara fungsional berperan sebagai sel inflamasi.
Gambar 2.4 Hirarki organisasi granul PMN yang dikeluarkan (Soehnlein, 2008)

56
Lebih dari 300 protein terkandung dalam granula yang terlibat dalam
proses netrofil meliputi aktivitas adhesi, migrasi dan antibakteri (Fox dkk., 2010).
Netrofil minimal memiliki 4 granula yang berbeda atau tipe vesikel yang berbeda
yaitu:
a. Secretory granule atau vesikel sekretorik merupakan suatu easily mobilizable
compartment, yang mengandung alkalin fosfatase dan protein plasma seperti
albumin serum. Vesikel sekretorik pada dasarnya merupakan vesikel endositik
yang pertama kali dimobilisasi untuk memfasilitasi rekruitmen dan migrasi
netrofil. Vesikel sekretorik mengandung reservoir dari protein membran yang
dimasukkan ke dalam protein membran luar dari netrofil mengikuti
pengeluaran vesikel untuk memulai respon inflamasi. β-integrin, complement
receptors I (CR1), CD14, CD16 dan formyl peptide receptors semuanya
diregulasi pada membran netrofil mengikuti aktivasi vesikel sekretorik.
Konsentrasi HBP yang bermakna dapat ditemukan mengikuti aktivasi vesikel
sekretorik dan diduga bekerja sebagai chemoattractant yang berimplikasi
penting dalam resolusi respon inflamasi (Fox dkk., 2010).
b. Primary atau azurophilic granule yang mengandung myeloperoxidase (MPO),
protein bakterisidal, dan proteinase. Primary atau azurophilic granule
terutama berespon terhadap sebagian besar dari efektor antimikroba, yang
mengandung alpha-defensin dan serine protease, elastase (Sohnlein, 2008;
Fox dkk., 2010).
c. Secondary atau specific granule yang menyimpan lactoferrin dan enzim-
enzim seperti kolagenase dan gelatinase (Sohnlein, 2008).

57
d. Tertiary atau gelatinase granule, menyerupai granula spesifik, yang
mengandung tissue degrading enzymes (Sohnlein, 2008; Fox dkk., 2010).
Keempat tipe granula dimobilisasi pada berbagai tahap proses inflamasi.
Secondary granule dan tertiary granule merupakan granul granul berikutnya yang
dimobilisasi, yang akhirnya diikuti oleh primary granule. Koordinasi dan waktu
dikeluarkannya dari berbagai subset granule yang berbeda memungkinkan respon
yang sesuai pada saat yang tepat dan meminimalkan potensi penghancuran netrofil
(Tapper dkk., 2002). Diantara protein protein yang tersimpan dalam granula
netrofil adalah HBP yang juga disebut azurocidin atau cationic antimicrobial
protein of 37 kDa (CAP37). Protein netrofil yang lain dalam famili ini adalah
capthesin G dan proteinase 3.
2.16 Heparin-Binding Protein (HBP)
Heparin Binding Protein (HBP) pertama kali diidentifikasi oleh Shafer
dkk., pada tahun 1984. HBP pada awal ditemukan sangat menarik karena HBP
memiliki aktivitas antimikroba terhadap bakteri gram positif (enterococcus
faecalis), bakteri gram negative (Escherichia coli) dan Candida albicans.
Akhirnya diketahui bahwa HBP memiliki peranan kuat dalam aktivitas
memodulasi respon imun. Kedua aktivitas HBP (antimikroba dan imunomodulasi)
juga dibantu oleh granula protein yang lain seperti α-defensin atau LL-37 yang
secara bersama sama disebut alarmins (Linder dkk., 2012).
2.16.1 Struktur dan fungsi HBP
Berdasarkan berat dan ukuran molekulnya Heparin-Binding Protein (HBP)
disebut juga dengan cationic antimicrobial protein of 37 kDa atau CAP37. Gabay

58
dkk., mengidentifikasi suatu PMN-derived bactericidal protein dari azurophilic
granule PMN manusia yang diberi nama azurocidin. Flodgard dkk., mengisolasi
suatu protein yang sama dari sel PMN manusia dan porcine yang mempunyai
kemampuan berikatan kuat pada heparin dan memberi nama Heparin-Binding
Protein. Sekuensing gen yang lengkap menunjukkan bahwa CAP37, azurocidin
dan HBP merupakan protein yang sama (Soehnlein, 2008).
HBP merupakan glikoprotein yang disintesis oleh netrofil. Secara
struktural HBP termasuk superfamili serine protease, walaupun sebanyak 45%
sekuen HBP identik dengan human neutrophil elastase, namun HBP merupakan
homolog inactive serine-protease yang memiliki berbagai fungsi (Tapper dkk.,
2002; Watorek, 2003).
Azurocidin atau HBP dihasilkan dari vesikel sekretorik atau granula
primer PMN. HBP disimpan dalam vesikel sekretorik bersama dengan granula
primer. Azurocidin atau HBP merupakan suatu protein yang dapat bergerak
dengan cepat dari leukosit polimorfonuklear (PMN) yang teraktivasi (Tapper dkk.,
2002).
Azurocidin/HBP memiliki aktivitas antimikroba yang sangat luas terutama
melawan bakteri gram negatif. Azurocidin juga dikenal sebagai mediator
inflamasi yang memiliki multifungsi, diantaranya berperan dalam meningkatkan
efek kontraksi pada sel endotel yang mengakibatkan peningkatan permeabilitas
pembuluh darah, meningkatkan kemampuan ikatan terhadap endotoksin dan
kemampuan menarik monosit ke tempat inflamasi (Watorek, 2003).

59
Pada awalnya HBP teridentifikasi memiliki aktivitas antimikroba, namun
saat ini HBP diduga juga berperan dalam beberapa proses inflamasi. HBP
dikeluarkan dari vesikel sekretorik netrofil yang teraktivasi pada saat kontak
dengan endotel. Ketika dikeluarkan, HBP menginduksi calcium–dependent
rearrangement dari sitosekeleton sel endotel. Hal ini mengakibatkan kontraksi sel
dan meningkatkan permeabilitas endotel. HBP juga diinternalisasi oleh sel endotel
untuk melindungi sel endotel dari apoptosis (Soehnlein, 2008).
HBP juga dihasilkan dari vesikel sekretorik ketika M-protein atau komplek
fibrinogen yang dibentuk oleh M-protein dikeluarkan dari permukaan sel bakteri,
yang berinteraksi dengan β2-integrin pada permukaan sel netrofil (Chertov dkk.,
1997; Lee dkk., 2002; Soehnlein, 2008).
Pada tempat infeksi, HBP juga disekresi oleh granula azurophilic selama
fagositosis. Hal ini menunjukkan aktivitas antimikroba HBP dan
responsibilitasnya terhadap rekrutmen dan aktivivasi monosit dan mediator
inflamasi yang lain. HBP juga diinternalisasi oleh monosit untuk pertahanan
jangka panjang dan meningkatkan produksi sitokin. Selanjutnya, HBP secara
langsung berperan dalam memelihara dan melanjutkan proses inflamasi
(Soehnlein, 2008; Soehnlein dan Lindbom, 2009; Linder dkk., 2010).
HBP berlokasi sangat dekat dengan membran plasma. HBP disimpan
dalam granula azurophilic dan vesikel sekretorik dan terdeteksi dalam jumlah
kecil sampai ke membran plasma. Adanya HBP dalam vesikel sekretorik
tergantung pada tahap diferensiasi sel. HBP lebih banyak terkandung dalam
netrofil matur dibandingkan dengan pada sel promielositik. HBP dapat dengan

60
mudah disintesis atau ditarget menjadi mobilized compartment pada stadium akhir
dari maturasi netrofil (Lindmark dkk., 1999; Tapper dkk., 2002).
Kemampuan netrofil untuk mensekresi HBP dari vesikel sekretorik
menjadi penting dalam fungsi proinflamasi seperti misalnya menimbulkan
perubahan permeabilitas pembuluh darah (Lindmark dkk., 1999; Tapper dkk.,
2002).
Diantara protein protein yang tersimpan dalam granula netrofil, HBP lebih
cendrung berperan dalam penyakit infeksi. Protein netrofil yang lain dalam
kelompok famili ini adalah capthesin G dan proteinase 3 (Tapper dkk., 2002).
Perbedaan HBP dengan proteinase 3 ini adalah HBP tidak memiliki aktivitas
enzimatik karena penggantian 2 asam amino esensial dalam trias katalisis. HBP
merupakan protein yang memiliki multifungsi, walaupun tidak mempunyai
aktivitas enzimatik (Tapper dkk., 2002).
Azurocidin atau HBP dapat berperan sebagai alarm dalam sistem imun
berbagai jalur yang berbeda dan kemudian membantu mediator penting selama
inisiasi respon imun. HBP berperan sebagai kemoatraktan dan aktivator monosit
dan makrofag. Secara fungsional berakibat meningkatkan pengeluaran sitokin dan
fagositosis bakteri, yang mendasari bacterial clearance yang efisien (Soehnlein
dkk., 2009).
Aktivasi leukosit oleh HBP/azurocidin dimediasi melalui β2 integrins dan
azurocidin-induced chemotaxis tergantung pada formyl-peptide receptors.
Sebagai tambahan, azurocidin mengaktivasi sel endotel yang menimbulkan
perembesan plasma (vascular leakage) dan pembentukan edema. Karena alasan

61
ini target pengeluaran azurocidin dan aksinya mungkin mempunyai potensi efek
terapi dalam kondisi inflamasi (Soehnlein dan Lindbom, 2009).
Sebagai rangkuman, beberapa fungsi HBP adalah (Lindmark dkk., 1999;
Watorek, 2003):
1. Memiliki aktivitas antimikroba yang kuat karena sifat kationisitasnya dan
sifat hidrofobisitasnya sehingga disebut sebagai komponen oxygen
independent host defense.
2. Dapat merekrut dan mengaktivasi monosit untuk memobilisasi sel T.
3. Menginduksi detachment dan agregasi homotipik dari sel endotel dan
fibroblast.
4. Endositosis HBP oleh monosit memacu produksi lipopolysaccharide-
induced tumor necrosis factor-α .
5. Internalisasi HBP mencegah apoptosis sel endotel.
6. HBP berperan dalam mediasi mengubah permeabilitas dinding pembuluh
darah yang dibangkitkan oleh aktivasi chemoattractant-induced PMN.
Beberapa fungsi ini menyiratkan bahwa HBP harus dimobilisasi lebih awal
dibandingkan dengan kandungan granula azurophilic yang lain dan mungkin
sebelum netrofil keluar dari pembuluh darah (Tapper dkk., 2002).
2.16.2 HBP sebagai pencetus peningkatan permeabilitas pembuluh darah
Sel endotel merupakan target pertama polymorphonuclear leucocyte-
derived HBP yang dilepaskan dari vesikel sekretori. Beberapa interaksi
menimbulkan aktivasi endotel dan sintesis de novo subsekuen dari cell adhesion
molekul (CAM) yang akhirnya meningkatkan penangkapan sel inflamasi.

62
Gangguan pada fungsi barier endotel menimbulkan perembesan plasma dan
pembentukan edema yang merupakan gambaran khas reaksi inflamasi (Linder,
dkk.,2009; 2012).
Studi sebelumnya menunjukkan bahwa emigrasi netrofil bersamaan
dengan efflux plasma dari dalam pembuluh darah dan sel netrofil ini berperan
sebagai pencetus perubahan permeabilitas sel pembuluh darah. Adesi netrofil dan
aktivasi melalui β2-integrin sangat penting pada netrofil dalam membangkitkan
peningkatan permeabilitas. Adesi netrofil pada sel endotel segera menginduksi
mobilisasi Ca2+ intraseluler dalam kedua tipe sel tersebut, yang memerintah granul
eksositosis dalam netrofil dan mengatur kembali sitoskeleton sel sel endotel. Blok
terhadap fungsi β2-integrin secara lengkap dapat membatalkan respon ini (Gautam
dkk., 2001; Sohnlein dan Lindbom, 2009).
Beberapa granul protein yang dikeluarkan diduga penting terlibat dalam
perubahan permeabilitas yang dimediasi neutrofil. Beberapa protein yang
merupakan anggota serprocidin, famili cathepsin G, elastase, proteinase-3 dan
inactive serine protease HBP. HBP memprovokasi dengan cepat peningkatan Ca2+
bebas sitosolik dalam sel endotel yang berdekatan, formasi actin stress fibers dan
peningkatan permeabilitas paraseluler. Respon stimulasi HBP identik dengan
yang dicapai oleh stimulasi kemoatraktan oleh netrofil dan imuno-netralisasi HBP
dalam neutrophil-derived secretion yang secara lengkap menghambat aktivitas
tersebut. Hal ini memperkuat peranan penting HBP dalam neutrophil-evoked
alteration dalam permeabilitas pembuluh darah. Walaupun demikian mekanisme
pasti aktivitas jalur sinyal HBP dalam sel endotel dan reorganisasi stimulasi

63
sitoskeletal dan kompleks junction masih sukar dipahami. Gambar 2.5. adalah
skema efek HBP pada infeksi bakteri (Linder dkk.,2010).
Gambar 2.5. Skema tentang efek HBP pada infeksi bakteri (Linder dkk., 2010).
2.16.3 Peran HBP dalam infeksi bakteri sistemik (sepsis)
Sepsis merupakan infeksi sistemik yang berat. Patofisiologi sepsis
merupakan suatu dinamika sindrom yang kompleks akibat ketidakseimbangan
antara mekanisme proinflamasi dan anti-inflamasi. Invasi mikroorganisme atau
kerusakan komponen jaringan dapat menstimulasi sel sel imun, yang
menimbulkan pelepasan sitokin proinflamasi, rective oxygen species (ROS) dan
enzim enzim. Sepsis didiagnosis berdasarkan tanda-tanda/gejala klinis dan tanda
laboratorium yang menunjukkan adanya inflamasi sistemik. Beberapa marker

64
yang diyakini sebagai biomarker awal sepsis adalah procalcitonin, interleukin-6,
laktat dan C-reactive protein (CRP). Walaupun demikian keakuratan diagnosis
maupun prognostik dari marker marker tersebut masih tidak konsisten dan
hasilnya bervariasi tergantung pada beratnya penyakit (Linder dkk., 2009; 2012).
Heparin binding protein (HBP) memiliki aktivitas imunomodulasi yang
kuat. Aktivitas antimikroba maupun aktivitas imunomodulasi HBP juga dimiliki
oleh protein netrofil granul yang lain diantaranya α-defensin atau LL-37 yang
secara bersama sama disebut dengan nama alarmins. HBP menunjukkan
mekanisme kerja yang unik diantara protein protein yang lain. Berbeda dengan
defensin atau LL-37 yang hanya keluar melalui eksositosis mengikuti aktivasi
netrofil, maka HBP dikeluarkan hampir secara lengkap ke lingkungan
ekstraseluler. HBP disimpan dalam 2 subset granul yang berbeda yaitu dalam
azurophilic/granul primer dan vesikel sekretorik. Hal ini membedakan HBP dari
granul protein yang lain. Granul protein HBP disintesis dalam netrofil. HBP
menginduksi rearrangement sel endotel sitoskeleton pada saat dikeluarkan dari
netrofil yang teraktivasi sehingga mengakibatkan peningkatan permeabilitas sel
endotelium. Pada tempat infeksi HBP berespon terhadap rekruitmen dan aktivasi
monosit dan mediator inflamasi lainnya. HBP juga diinternalisasi oleh monosit
sehingga bertahan lama dan memicu produksi sitokin. HBP selanjutnya berperan
dalam memelihara dan mengembangkan inflamasi (Lee dkk., 2002; Sohnlein,
2008; Linder dkk., 2012).
Pada penderita febris dengan suspek sepsis, HBP telah terbukti merupakan
prediktor terbaik dari sepsis yang berat dengan kurve ROC (Receiver-Operating

65
Characteristic) menunjukkan AUC (Area Under Curve) 0,95 yang lebih baik
dibandingkan dengan procalcitonin, IL-6 dan laktat (Linder dkk., 2009; 2012).
Telah dibuktikan bahwa pengukuran HBP dapat digunakan untuk
mengidentifikasi pasien yang berisiko berkembang menjadi sepsis dengan
kegagalan sirkulasi. HBP menunjukkan sensitivitas 87,1% dalam mendiagnosis
sepsis berat dengan atau tanpa syok dan memiliki spesifisitas 95,1%, Positive
Predictive Value (PPV) 88,4% dan Negative Predictive Value (NPV) 94,5%
dimana nilai nilai tersebut melebihi dari nilai untuk marker marker lain yang telah
diuji. Kurve receiver-operating characteristic (ROC) juga menunjukkan bahwa
HBP merupakan prediktor terbaik dari sepsis berat dengan nilai area under curve
(AUC) 0,949 (IK 0,917; 0,982) (Linder, 2012).
HBP merupakan induser kuat terjadinya perembesan plasma, yang
merupakan tanda khas pada sepsis yang berat mengakibatkan ekstravasasi cairan
plasma dan hipotensi. Median kadar HBP plasma lebih tinggi pada pasien yang
mengalami atau berkembang menjadi sepsis berat daripada pasien sepsis, infeksi
ringan atau SIRS tanpa infeksi. Hubungan yang kuat antara konsentrasi HBP yang
tinggi dan berkembangnya sepsis dengan kegagalan sirkulasi mendukung peranan
HBP sebagai mediator perembesan (leakage) kapiler pada infeksi. Untuk
perkiraan kadar HBP sebagai marker sepsis, pada studi sebelumnya dibandingkan
dengan biomarker lain seperti prokalsitonin, interleukin-6, CRP dan laktat. Semua
marker lain menunjukkan pola yang hampir sama dengan kadar yang lebih tinggi
pada pasien dengan sepsis yang berat. Kadar plasma HBP tampaknya menjadi
parameter paling kuat untuk membedakan pasien dengan sepsis berat dengan

66
kegagalan sirkulasi dibandingkan dengan infeksi yang kurang berat. Pada batas
kadar 15 ng/ml, sensitivitas HBP dalam mendiagnosis sepsis berat 87% dan
spesifisitasnya 95%. Kadar HBP pada pasien dengan sepsis berat meningkat
sebelum hipotensi yang bermakna terdeteksi. Pada pasien pasien tersebut kadar
plasma HBP meningkat 12 jam sebelum onset hipotensi. Jadi HBP merupakan
neutrophil-derived protein yang bekerja sebagai amplifier/penguat respon
inflamasi dan menginduksi kebocoran (leakage) kapiler. Selama sepsis terdapat
peningkatan kadar HBP dalam plasma, dan kadarnya berkorelasi dengan
terjadinya hipotensi dan kegagalan sirkulasi (Linder dkk., 2012).
2.16.4 Ekspresi HBP pada berbagai penyakit infeksi bakteri
HBP juga meningkat pada kulit selama terjadi infeksi jaringan lunak dan
mungkin berkontribusi terhadap berkembangnya komplikasi seperti edema. Telah
ditunjukkan juga bahwa M1/kompleks fibrinogen mengaktivasi netrofil untuk
degranulasi dan keluarnya HBP. Pada studi lain, biopsi jaringan dari penderita
dengan necrotizing fasciitis atau selulitis berat yang disebabkan oleh S. pyogenes
menunjukkan bahwa rekruitmen netrofil dan monosit/makrofag pada fokus lokasi
infeksi disertai keluarnya HBP. Studi ini mendukung bahwa vasoaktif HBP
memainkan peranan penting dalam terbentuknya edema pada infeksi kulit oleh
streptokokus (Linder dkk., 2010a).
HBP ditemukan lebih tinggi pada luka ulkus kronis dibandingkan dengan
luka ulkus yang akut. Pada kultur supernatan luka ditemukan kuman Proteus
mirabilis, E. faecalis dan Pseudomonas aeruginosa yang menginduksi
pengeluaran HBP yang bermakna dari total HBP dalam netrofil. HBP tidak

67
didegradasi oleh elastase P. aeruginosa. Elastase merupakan metalloproteinase
yang kuat yang mendegradasi peptida antimikroba seperti LL-37. Hasil temuan
ini menunjukkan bahwa HBP stabil dalam lingkungan proteolitik dan efek HBP
terhadap permeabilitas endotel dan rekruitmen netrofil mungkin mewakili tahap
patogenik awal dalam berkembangnya ulkus pada kulit (Lundqvist dkk., 2004).
Kadar plasma HBP berkorelasi dengan berkembangnya ARDS (acute
respiratory distress syndrome) pada pasien trauma dibandingkan dengan pasien
yang tidak mengalami ARDS. HBP berpotensi sebagai biomarker untuk
mendeteksi berkembangnya ARDS setelah trauma (Johansson dkk., 2013).
Penelitian mengenai kadar HBP dalam urin pada penderita infeksi saluran
kencing juga menunjukkan bahwa kadar Urine-HBP diatas 32 ng/ml memiliki
sensitivitas 93,3% dan spesifisitas 90,3% dalam mendeteksi infeksi saluran
kencing. Hasil penelitian ini mengindikasikan bahwa analisis U-HBP dapat
membantu dalam tatalaksana infeksi saluran kencing (Kjolvmark dkk., 2012).
2.16.5 Ekspresi HBP pada meningitis bakterialis
Gambaran klinis pasien meningitis bakterialis dan infeksi SSP oleh virus
dapat sangat mirip dan terkadang hasil analisis CSS tidak dapat digunakan untuk
mengambil kesimpulan diagnosis yang pasti. Penelitian pertama yang mencari
adanya neutrophil-derived HBP dalam CSS menunjukkan bahwa HBP dapat
membedakan antara infeksi bakteri dan virus pada SSP. Secara konvensional,
adanya pleositosis dalam CSS dapat digunakan sebagai dasar yang bermanfaat
dalam membedakan antara penyebab bakteri dan virus, dimana pleositosis ringan
atau limfositosis mengindikasikan penyebab non bakterial. Walaupun demikian

68
terdapat juga kasus infeksi meningitis bakterialis yang pada pemeriksaan analisis
CSS tidak ditemukan adanya pleositosis. Sensitivitas CSS dengan pleositosis >
1000 x106 sel/L hanya 61% dan spesifisitasnya hanya 68% sebagai marker infeksi
CSS pada pasien paska bedah saraf (Ross dkk., 1998). Sebesar 61% pasien dengan
meningitis bakterialis akut dan 3% pasien dengan infeksi virus pada SSP
menunjukkan pleositosis CSS> 1000 x 106 sel/L. Predominan neftrofil ditemukan
pada 73% pasien dengan meningitis bakterialis akut dan 7% pasien dengan infeksi
virus pada SSP (Linder dkk., 2011).
Pada pasien yang secara klinis suspek meningitis bakterialis, seringkali
antibiotika diberikan sebelum dilakukan punksi lumbal. Hal ini menurunkan
diagnosis secara mikrobiologi dan memperpanjang terapi antibiotika pada
beberapa kasus. Sekitar 63% pasien telah mendapat terapi antibiotika 48 jam
sebelum dilakukan punksi lumbal. Pada penelitian tersebut diperoleh hasil 24 dari
26 pasien yang telah mendapat antibiotika masih menunjukkan peningkatan kadar
HBP. Standar baku emas diagnosis meningitis bakterialis adalah kultur CSS yang
positif dan risiko kesalahan diagnosis sangat besar apabila hasil kultur negatif.
Ditemukan fakta semua pasien meningitis bakterialis yang hasil kulturnya positif
menunjukkan hasil kadar HBP >20 ng/ml. Hal inilah yang mendukung dugaan
bahwa HBP berpotensi sebagai biomarker untuk mendukung diagnosis meningitis
bakterialis terutama bila hasil kultur menunjukkan hasil negatif (Linder dkk.,
2011).
HBP menginduksi kebocoran plasma dan pembentukan edema. Edema
pada otak merupakan suatu kondisi yang serius dan seringkali dapat menimbulkan

69
komplikasi yang bersifat fatal pada meningitis bakterialis. Semua pasien
meningitis bakterialis yang meninggal memiliki kadar HBP sangat tinggi (diatas
385 ng/mL). Sesuai dengan efek HBP pada sel endotel yang mengakibatkan
kebocoran plasma, maka hal ini mengindikasikan bahwa HBP memegang peranan
dalam berkembangnya edema serebri pada meningitis bakterialis (Linder dkk.,
2011).
Masalah pada penelitian tersebut adalah pada metodologinya, mereka
menggunakan 2 kelompok sampel yang berbeda, dimana sebagian kelompok
sampel merupakan data prospektif dan sebagian kelompok merupakan data
retrospektif yang diambil 10 tahun sebelumnya. Mereka juga menggunakan HBP
untuk mendiagnosis meningitis bakterialis pada pasien yang tidak dapat
didiagnosis dengan pasti. Sensitivitas dan spesifisitas kadar HBP dalam CSS
dibandingkan dengan jumlah sel polimorfonuklear dan hitung leukosit dalam CSS
yang secara statistik identik (Koenig, 2011).
2.16.6 Pengukuran kadar HBP
Terdapat beberapa Assay Kit untuk pemeriksaan human Heparin Binding
Protein yang telah ada, diantaranya: Human Heparin Binding Protein (HBP)
ELISA kit dari My.Biosource.com; Heparin Binding Protein (AZU/HBP)
BioAssay ELISA Kit (human); Axis-Shield Heparin Binding Protein (HBP) ELISA
kit. Cusabio -Human Azurocidin/Heparin Binding Protein (AZU/HBP) ELISA Kit.
Kit Elisa untuk HBP yang dapat diperoleh di Indonesia adalah Heparin
Binding Protein (AZU/HBP) BioAssay ELISA Kit (human), merupakan teknik
Sandwich Enzyme Immunoassays kuantitatif. BioAssay-ELISA Kit merupakan 1,5

70
jam solid-phase Sandwich ELISA yang dirancang untuk menentukan HBP/AZU
secara kuantitatif dengan metode deteksi colorimetric.
2.17 Protein S100B
Protein S100 adalah salah satu calcium binding peptide yang digunakan
sebagai parameter aktivasi glia dan atau kematian pada sebagian besar gangguan
susunan saraf pusat (SSP). Marker otak ini memainkan peranan penting pada
perkembangan SSP secara fisiologis dan pemulihan otak setelah mengalami
cedera (Yardan dkk., 2011).
Protein S100B terutama diekspresikan oleh sel sel pada sistem saraf pusat
terutama astroglia dan sel neuron. Protein S100B terutama ditemukan pada
astroglia dan sel Schwann, namun juga dapat dihasilkan dari sumber-sumber
ekstra serebral. S100B memiliki peran parakrin dan autokrin terhadap neuron dan
glia. Sebagai salah satu marker neurokimia otak, S100B juga dapat digunakan
sebagai marker adanya kerusakan pada proses kegagalan sirkulasi, stroke, dan
cedera kepala (Bouvier dkk., 2011;Yardan dkk., 2011).
Protein S100B sering digunakan sebagai parameter dari aktivasi sel glia
dan atau parameter kematian sel dari beberapa kelainan SSP. Protein S100B
memiliki peran penting dalam perkembangan normal SSP dan pemulihan setelah
cedera. Protein S100B juga diduga memiliki hubungan dengan adanya kerusakan
neuronal dan stress oksidatif yang dihubungkan dengan ensefalopati sepsis atau
meningitis bakterialis pada anak-anak. Peningkatan kadar S100B berhubungan

71
dengan cedera patologis atau tingkat keparahan klinis pada berbagai kelainan SSP
(Hamed dkk., 2009; Yardan dkk., 2011).
Protein S100B dapat ditemukan dalam darah tepi, urin, tali pusat, cairan
serebrospinal (CSS), cairan amnion dan dalam konsentrasi yang lebih tinggi
ditemukan pada air susu. Konsentrasi S100B dalam serum lebih rendah
dibandingkan dengan dalam cairan serebrospinal (Yardan dkk., 2011).
2.17.1 Struktur famili protein S100B
Konsep brain-specific proteins pertama kali disebutkan oleh Moore, pada
tahun 1965. Ditemukan 3 jenis asam protein dengan berat molekul rendah, dengan
konsentrasi yang tinggi pada fraksi otak. Tetapi konsentrasinya rendah pada fraksi
hepar. Ketiga protein tersebut diberi nama: protein 14-3-2, protein 14-3-3 dan
protein S100. Diberi nama S100 karena protein tersebut larut 100% dalam
ammonium sulphate pada pH netral. Banyak protein yang disebut “brain-specific
protein” yang sesungguhnya tidak spesifik terhadap otak tetapi konsentrasinya
pada otak jauh lebih tinggi daripada jaringan lain, sehingga dapat digunakan
sebagai marker kerusakan otak (Sen dan Belli, 2007).
Sejak pertama ditemukan S100 disebut sebagai brain-specific proteins
secara berlebihan. Saat ini diketahui bahwa S100 merupakan superfamili dari low
molecular weight acidic calcium binding proteins. Pada awalnya protein ini
dianggap spesifik terhadap otak, namun penelitian selanjutnya menunjukkan
bahwa protein S100 ditemukan pada jaringan lain tetapi dalam konsentrasi yang
sangat rendah dibandingkan yang ditemukan dalam SSP (Sen dan Belli, 2007).

72
S100 merupakan protein unik yang berukuran 10-12 kDa berlokasi di
dalam kromosom 1q21. Protein S100 terdiri atas 2 kelompok protein yang mirip
dan berhubungan yaitu S100B dan S100A1. Berdasarkan analisis sekuen, protein
S100B dan S100A1 merupakan calcium-binding protein dari EF hand type (EF
hand merupakan rantai asam amino yang terlibat dalam ikatan), secara struktural
sangat berhubungan dengan EF hand calcium binding protein seperti troponin C,
calmodulin dan parvalbumin (Yardan dkk., 2011; Park dkk., 2013).
Protein S100 merupakan famili multigenik dari protein berukuran kecil,
khas ditandai oleh 2 ikatan kalsium dengan bentuk helix E-loop-helix F (“EF-hand
type”). Sampai saat ini telah ditemukan 25 anggota protein S100 yang hanya
diekspresikan dalam vertebrata. Protein S100 terdiri atas anggota sebagai berikut
(Steiner dkk., 2011):
a. Sebanyak 21 anggota yang ditemukan pada vertebrata yang
mempunyai kluster gen pada lokus kromosom 1q21 yaitu S100A1-
S100A18, trichohyalin, filaggrin dan repetin.
b. Anggota yang lain ditemukan pada manusia pada lokus kromosom
4p16 (S100P), 5q14 (S100Z), 21q22 (S100B) dan Xp22 (S100G).
Bagian utama dari anggota famili S100 adalah Ca2+ sensor proteins, yang salah
satunya diaktivasi oleh second messenger Ca2+, berinteraksi dengan target
protein intraseluler, selanjutnya mengatur aktivitasnya. Calcium merupakan
universal second messenger yang berperan dalam proses pengaturan konduksi dan
transmisi impuls saraf, kontraksi otot, motilitas sel, pertumbuhan dan diferensiasi
sel, ekspesi gen, komunikasi antara berbagai sistem enzim, apoptosis dan nekrosis.

73
Sel memiliki sekelompok protein intraseluler yang berikatan dengan kalsium yang
bekerja mengatur kadar kalsium sitosolik atau tranduksi sinyal kalsium
intraseluler.
Ekspresi protein S100 tampaknya konsisten antar ras, tidak berbeda antara
laki dan perempuan, tidak menunjukkan variasi sirkadian serta tidak tergantung
pada usia stabil dalam serum sampai minimal 7 bulan dan berbagai perannya
dibagi menjadi 2 kelompok yaitu aktivitas intraseluler dan ekstraseluler (Sen dan
Belli, 2007; Steiner dkk., 2011).
Protein S100B merupakan bagian dari suatu superfamili protein yang
memiliki efek sebagai calcium dependent-regulatory proteins yang memodulasi
aktivitas efektor protein atau sel (Sen dan Belli, 2007). Tidak seperti famili S100
yang lain, S100B terletak pada kromosom yang berbeda yaitu 21q22. Struktur
S100B homodimer dan tiap beta monomer memiliki berat 10,5 kDa stabil dalam
serum sampai minimal 7 bulan dan berbagai perannya dibagi menjadi 2 kelompok
yaitu aktivitas intraseluler dan ekstraseluler (Heizmann, 2004; Park dkk., 2013).
Protein S100B dihasilkan dalam SSP, terutama di dalam astroglia,
oligodendrosit dan sel Schwann yaitu pada sitosol yang melekat pada membran
sel. Sintesa dan sekresi terutama oleh astrosit pada otak (Yardan dkk., 2011; Kepa
dan Grzesik, 2013).
Beberapa sel di luar otak juga dapat menghasilkan protein ini, seperti sel
epitel ginjal, ependimosit, kondrosit, adiposit, limfosit, sumsum tulang dan
melanosit. Eliminasi S100B terutama terjadi melalui ginjal (Sun dkk., 2013).
2.17.2 Fungsi dan peranan protein S100B

74
Famili protein S100 terdiri atas 24 anggota yang secara fungsional dibagi
menjadi 3 subkelompok utama yaitu:
a. Hanya berperan dalam regulasi intraseluler.
b. Berperan dalam regulasi intraseluler dan ekstraseluler.
c. Berperan terutama dalam regulasi ekstraseluler.
Protein S100 di dalam sel terlibat dalam regulasi proliferasi, diferensiasi dan
apoptosis, Ca2+ homeostasis, metabolisme energi, inflamasi dan migrasi/invasi
melalui interaksi dengan berbagai target protein seperti enzim, subunit
sitoskeletal, reseptor, faktor transkripsi, dan asam nukleat. Beberapa protein S100
disekresi/dikeluarkan dan mengatur fungsi sel dalam suatu autokrin dan parakrin
melalui aktivasi surface receptors, G-protein couple receptors, scavenger
receptor, atau heparan sulfat proteoglicans dan N-glycan.
a. Fungsi intraseluler
Efek dari protein S100B tergantung dari konsentrasinya baik di dalam
maupun di luar sel. Pada konsentrasi nanomolar, S100B in vitro menstimulasi
pertumbuhan neurit pada neuron korteks serebri dan meningkatkan daya tahan
neuron dalam berbagai sistem selama tahap perkembangan. S100B secara in vitro
memiliki aktivitas neurotropik untuk sel saraf selama proses maturasi neuronal
dan proliferasi sel glia. S100B mengurangi kematian sel dan kehilangan fungsi
mitokondria dari kekurangan glukosa. S100B juga diduga memiliki peran dalam
perkembangan SSP dan pemulihan pasca cedera dengan efek neurotropik dan
gliotropik yang dimilikinya (Yardan dkk., 2011).

75
Protein S100B intraseluler berfungsi sebagai stimulasi proliferasi sel,
migrasi dan inhibitor apoptosis dan diferensiasi, yang memiliki peranan penting
dalam perkembangan dan perbaikan sel otak yang cedera, kartilago dan otot lurik,
aktivasi astrosit pada cedera otak dan proses neurodegenerasi dan remodeling
kardiomiosit setelah infark.
Protein S100B terekspresi dalam astrosit, sekelompok sel neuronal
tertentu, sel Schwann, melanosit, kondrosit, adiposit, myofiber otot skeletal dan
sel satelit yang berhubungan, sel dendritik tertentu dan limfosit dan beberapa tipe
sel lain. Aktivitasnya sebagai stimulator proliferasi sel dan migrasi sel dan
inhibitor apoptosis dan diferensiasi, yang menimbulkan implikasi penting selama
perkembangan dan regenerasi otak, kartilago, otot skeletal, aktivasi astrosit pada
kerusakan tertentu pada otak, proses neurodegeneratif dan remodeling
kardiomiosit setelah infark, demikian juga melanomagenesis, dan gliomagenesis
(Donato dkk., 2013).
Protein S100B memiliki beberapa peran penting dalam sistem saraf, salah
satunya adalah memodulasi proliferasi dan diferensiasi sel neuron dan glia.
Protein ini juga berperan dalam transduksi sinyal melalui proses inhibisi
fosforilasi protein dan juga regulasi aktivitas enzim dan homeostasis kalsium.
S100B juga berperan dalam regulasi morfologi sel dengan berinteraksi dengan
sitoskeleton sitoplasma (Aleksovska dkk., 2014).
b. Fungsi ekstraseluler
S100B diekskresikan oleh sel sel ekstraseluler terutama melalui aktivitas
cytokine-like dengan efek parakrin pada neuron. Informasi mengenai implikasi

76
fungsional sekresi S100B oleh astrosit ke dalam ruang ekstraseluler masih belum
jelas, tetapi terdapat bukti mendasar bahwa sekresi S100B oleh sel glia
mengekspresikan efek toksik atau tropik tergantung pada konsentrasinya.
Implikasi protein S100B yang luas ini menimbulkan para peneliti
menggambarkannya sebagai CRP dari otak (Sen dan Belli, 2007).
Konsentrasi S100B ekstraseluler dapat berefek tropik pada sel, tetapi
dalam konsentrasi patologis menyebabkan aktivasi glia dan apoptosis. Secara in
vitro, S100B ekstraseluler menstimulasi ekspresi sitokin proinflamasi dan
menghasilkan efek neurotoksik yang menginduksi apoptosis. Pada konsentrasi
mikromolar, S100B memiliki efek yang merugikan. Penelitian terkini
menunjukkan bahwa S100B pada konsentrasi mikromolar dapat menyebabkan
apoptosis dengan cara berinteraksi dengan Receptor for Advance Glycation End
Products (RAGE), yang dapat menyebabkan peningkatan Reactive Oxygen
Species (ROS), pelepasan sitokrom C dan aktivasi kaskade caspase (Donato,
2009; Yardan dkk., 2011).
S100B diduga dapat berkontribusi terhadap perubahan patologik dalam
proses neurodegenerasi maupun penyakit inflamasi otak dengan aktivasi
mikroglia. Saat terjadi cedera metabolik, misalnya keterbatasan oksigen dan
glukosa, proses yang paling dini terjadi sebelum adanya respon glia adalah
pelepasan S100B (Sen dan Belli, 2007; Yardan dkk., 2011).
Konsentrasi S100B yang tinggi akan menyebabkan kerusakan neuronal
melalui pelepasan nitrit oksida (NO) oleh astrosit. Waktu paruh biologis S100B
adalah sekitar 30 menit. Hal ini memungkinkan terjadinya kenaikan persisten

77
konsentrasi S100B dalam serum yang merefleksikan pelepasan S100B secara
kontinyu dari jaringan yang mengalami cedera tersebut (Yardan dkk., 2011).
Protein S100B sebagian besar diekspresi oleh astrosit, namun tidak semua
astrosit dapat mengekspresikan protein S100B ini. Protein S100B hanya
diekspresikan oleh subtipe astrosit yang matur yang melapisi pembuluh darah dan
memiliki sel ekspresi NG-2. Penelitian terkini menunjukkan informasi yang lebih
lengkap mengenai mekanisme S100B sebagai regulator intrasel dan sinyal
ekstrasel.
Fungsi S100B ekstraselular terlibat dalam mekanisme receptor for
advanced glycation end products (RAGE) dalam berbagai tipe sel yang berbeda,
baik yang memberikan efek yang menguntungkan atau merugikan, pro-proliferasi
atau pro-diferensiasi tergantung dari konsetrasi S100B yang dibentuk oleh tipe sel
dan lingkungan mikro (Donato dkk., 2009; Sun dkk., 2013).
Waktu paruh biologis dari S100B adalah 30 menit. Hal ini menunjukkan
bahwa peningkatan persisten dari kadar protein ini mencerminkan pengeluaran
yang terus menerus dari jaringan yang berkaitan. S100B dapat ditemukan di darah,
plasenta, urin, LCS, cairan amnion. Kadar S100B di serum lebih rendah daripada
di LCS (Yardan dkk., 2011).
S100B disekresi secara teratur oleh astrosit dan sekresinya diatur oleh
beberapa faktor. Faktor-faktor tersebut dapat dibagi menjadi faktor yang dapat
meningkatkan sekresi dari protein S100B dan faktor yang dapat menurunkan
sekresi S100B. Sel yang rusak atau nekrosis juga dapat melepaskan protein ini.
Keberadaan protein ini dalam cairan serebrospinal (CSS), serum dan cairan

78
amnion diatas ambang rata-rata dapat digunakan dalam kepentingan diagnostik
dan prognostik (Sorci dkk., 2010).
Gambar 2.6. menunjukkan skema representasi efek ekstraseluler protein
S100B pada neuron microglia, astrosit, myoblast, VSCM, kardiomyosit, dan saraf
perifer. (A) pada kondisi normal, secara fisiologis S100B yang disekresi oleh
astrosit menunjukkan efek tropik pada neuron dan memodulasi akivitas microglia
bersama dengan RAGE. (B) Jika S100B berada di ekstraseluler (di luar sel otak)
dalam konsentrasi tinggi, S100B mengaktivasi microglia dan astrosit sehingga
ikut berpartisipasi dalam respon inflamasi dan bersifat toksik terhadap neuron
dengan menstimulasi RAGE secara berlebihan. (C). Pada injuri saraf perifer akut,
S100B keluar daru sel Schwann, membantu makrofag dan sel Schwann bermigrasi
dan mengeluarkan trophic factor melalui RAGE. Dengan demikian berpartisipasi
dalam regenarasi saraf perifer.(D) Dalam konsentrasi sub-nanomolar sampai
nanomolar, S100B menstimulasi proliferasi myoblast dan menginhibisi
diferensiasi myoblast dengan cara meningkatkan sinyal bFGF/FGFR1. (E) Pada
dosis tinggi S100B menstimulasi proliferasi VSCM dan sekresi IL-6 dan MCP-1
dari VSCM melalui aktivitas RAGE sehingga potensial berpartisipasi dalam
atherogenesis. (F) Pada dosis tinggi S100B menyebabkan apoptosis kardiomiosit
melalui aktivitas RAGE (Donato dkk., 2013).
79
Gambar 2. 6. Skema representasi efek ekstraseluler protein S100B pada neuron microglia dan astrosit (Donato dkk., 2013)
2.17.3 Protein S100B sebagai marker pada penyakit neurologi
Sebagai suatu biomarker, S100B secara primer diproduksi oleh astrositosit
sehingga dapat mempresentasikan aktivasi astrosit. Sel sel penghasil S100B
terbanyak terdapat pada substansia grisea, sedangkan oligodendrogliosit
memproduksi S100B terbanyak di substatia alba (Yardan dkk., 2011)

80
S100B mRNA intraseluler maupun ekstraseluler dan kadar S100B tersebut
digunakan sebagai parameter aktivasi astrosit dan kematian sel pada beberapa
situasi cedera otak. Peningkatan konsentrasi pada serum atau CSS berhubungan
dengan berbagai gangguan SSP (Yardan dkk., 2011).
S100B merupakan biomarker untuk prediksi dini perkembangan tekanan
intrakranial yang meningkat. Sejak diketahui adanya peningkatan S100B dalam
serum dan CSS pasien cedera kepala, penelitian selanjutnya diarahkan pada
berbagai proses patologis intrakranial selain cedera kepala, misalnya pada kasus
kasus stroke iskemik, disfungsi otak karena hipoksia, proses degeneratif maupun
inflamasi (Yardan dkk., 2011).
Sebuah penelitian tentang hubungan protein S100B dan stress oksidatif
pada anak dengan meningitis sudah dilakukan oleh Hamed, dkk. Mekanisme
kerusakan SSP karena meningitis belum diidentifikasi secara jelas. Sepanjang
infeksi bakteri, netrofil dan makrofag bersama-sama bekerja pada daerah infeksi
untuk melawan mikroorganisme. Sel sel sistem imun dari darah dan parenkim otak
melepaskan spesies oksigen reaktif (ROS=reactive oxygen species) sebagai
bagian dari mekanisme pertahanan diri host melawan invasi bakteri. Pada proses
ini, sel sel mengkonsumsi molekul oksigen yang berubah menjadi anion
superoksida toksik, hidrogen peroksida dan radikal hidroksil. Dalam kondisi
normal, ROS dieliminasi oleh pertahanan enzim enzim seluler misalnya
superoksida dismutase dan katalase dan non enzimatik misalnya glutation dan
asam urat yang berfungsi sebagai mekanisme pertahanan antioksidan. Bila ROS
tidak dapat disingkirkan karena kelelahan kapasitas antioksidan, maka terjadi

81
ketidakseimbangan antara aktivitas oksidan dan antioksidan yang menghasilkan
stress oksidatif dan menyebabkan terjadinya peroksidasi membrane lipid,
kerusakan protein dan DNA. Proses ini memiliki peranan besar dalam berbagai
proses patologis pada meningitis bakterialis (Hamed dkk., 2009).
Protein S100B secara fisiologis diproduksi oleh astrosit dan berpengaruh
pada perkembangan dan pemeliharaan SSP. Walaupun mekanisme sekresi S100B
belum diketahui secara pasti, diperkirakan sekresinya berkaitan dengan stress
oksidatif. Kadar S100B yang tinggi (mikromolar) merupakan hasil kerusakan sel
sel glia atau reaksi astrositik terhadap cedera neuronal (reaktif astrogliosis) yang
selanjutnya meningkatkan kerusakan neuronal. S100B dengan kadar tinggi dapat
mengindikasikan kerusakan atau disfungsi SSP. S100B dalam konsentrasi yang
normal menjaga hipokampus dari toksisitas glutamate, namun pada konsentrasi
yang tinggi justru mengeksaserbasi neuroinflamasi, stress oksidatif dan apoptosis
neuronal. Peningkatan S100B menggambarkan kerusakan glia atau reaksi astrosit
terhadap adanya cedera neuronal. Hubungan antara S100B dengan stress oksidatif
atau mekanisme antioksidan pada pasien dengan meningitis bakterialis belum
banyak diteliti. Penelitian yang telah dilakukan oleh Hamed, dkk. (2009), menilai
kadar S100B pada serum dan CSS dari 40 anak dengan meningitis bakterialis
dibandingkan dengan 20 anak sehat tanpa meningitis. Pada penelitian tersebut
didapatkan bahwa kadar serum S100B pada anak dengan meningitis lebih tinggi
daripada anak sehat tanpa meningitis. Dari hasil penelitian tersebut diperoleh
asumsi sebagai berikut:

82
a. Terjadi disfungsi sawar darah otak pada pasien dengan meningitis
bakterialis.
b. Stress oksidatif memegang peranan penting pada pathogenesis
meningitis bakterialis.
c. Kerusakan otak yang terjadi pada meningitis ditandai dengan adanya
peningkatan sintesis S100B intratekal.
d. Terdapat hubungan antara kadar mediator stress oksidatif,
antiokasidan dan S100B yang kemungkinan berkorelasi dengan tingkat
keparahan penyakit dan timbulnya komplikasi neurologis.
Pada penelitian yang membandingkan antara meningitis bakterialis dan viral,
menunjukkan bahwa S100B lebih memiliki manfaat pada pasien dengan
meningitis bakterialis (Jung dkk., 2011).
2.17.4 Peran protein S100B pada infeksi SSP
Sebuah penelitian tentang hubungan protein S100B dan stress oksidatif
pada anak dengan meningitis sudah dilakukan oleh Hamed, dkk. Mekanisme
kerusakan SSP karena meningitis belum diidentifikasi secara jelas. Sepanjang
infeksi bakteri, netrofil dan makrofag bersama sama bekerja pada daerah infeksi
untuk melawan mikroorganisme. Sel sel sistem imun dari darah dan parenkim otak
melepaskan spesies oksigen reaktif (ROS=reactive oxygen species) sebagai
bagian dari mekanisme pertahanan diri host melawan invasi bakteri. Pada proses
ini, sel sel mengkonsumsi molekul oksigen yang berubah menjadi anion
superoksida toksik, hidrogen peroksida dan radikal hidroksil. Dalam kondisi
normal, ROS dieliminasi oleh pertahanan enzim enzim seluler misalnya

83
superoksida dismutase dan katalase dan non enzimatik misalnya glutation dan
asam urat yang berfungsi sebagai mekanisme pertahanan antioksidan. Bila ROS
tidak dapat disingkirkan karena kelelahan kapasitas antioksidan, maka terjadi
ketidakseimbangan antara aktivitas oksidan dan antioksidan yang menghasilkan
stress oksidatif dan menyebabkan terjadinya peroksidasi membrane lipid,
kerusakan protein dan DNA. Proses ini memiliki peranan besar dalam berbagai
proses patologis pada meningitis bakterialis (Hamed dkk., 2009).
Protein S100B secara fisiologis diproduksi oleh astrosit dan berpengaruh
pada perkembangan dan pemeliharaan SSP. Walaupun mekanisme sekresi S100B
belum diketahui secara pasti, diperkirakan sekresinya berkaitan dengan stress
oksidatif. Kadar S100B yang tinggi (mikromolar) merupakan hasil kerusakan sel
sel glia atau reaksi astrositik terhadap cedera neuronal (reaktif astrogliosis) yang
selanjutnya meningkatkan kerusakan neuronal. S100B dengan kadar tinggi dapat
mengindikasikan kerusakan atau disfungsi SSP. S100B dalam konsentrasi yang
normal menjaga hipokampus dari toksisitas glutamate, namun pada konsentrasi
yang tinggi justru mengeksaserbasi neuroinflamasi, stress oksidatif dan apoptosis
neuronal. Peningkatan S100B menggambarkan kerusakan glia atau reaksi astrosit
terhadap adanya cedera neuronal.
Hubungan antara S100B dengan stress oksidatif atau mekanisme
antioksidan pada pasien dengan meningitis bakterialis belum banyak diteliti.
Penelitian yang telah dilakukan oleh Hamed, menilai kadar S100B pada serum
dan CSS dari 40 anak dengan meningitis bakterialis dibandingkan dengan 20 anak
sehat tanpa meningitis. Pada penelitian tersebut didapatkan bahwa kadar serum

84
S100B pada anak dengan meningitis lebih tinggi daripada anak sehat tanpa
meningitis. Dari hasil penelitian tersebut diperoleh asumsi sebagai berikut (Hamed
dkk., 2009):
a. Terjadi disfungsi sawar darah otak pada pasien dengan meningitis
bakterialis.
b. Stress oksidatif memegang peranan penting pada pathogenesis
meningitis bakterialis
c. Kerusakan otak yang terjadi pada meningitis ditandai dengan adanya
peningkatan sintesis S100B intratekal.
d. Terdapat hubungan antara kadar mediator stress oksidatif,
antiokasidan dan S100B yang kemungkinan berkorelasi dengan tingkat
keparahan penyakit dan timbulnya komplikasi neurologis.
Pada penelitian yang membandingkan antara meningitis bakterialis dan viral,
menunjukkan bahwa S100B lebih memiliki manfaat pada pasien dengan
meningitis bakterialis (Jung dkk., 2011).
2.17.5 Pengukuran kadar protein S100B
S100B dapat diukur dengan beberapa metode antara lain:
Immunoradiometric Assay (IRMA), Immunoluminometric Assay (LIA), mass
spectroscopy, Western blot, Enzyme Linked Immunosorbent Assay (ELISA),
electrochemiluminescene dan Polymerase Chain Reaction (PCR) kuantitatif.
Masing masing metode berbeda dalam hal spesifisitas dan sensitivitas, aplikasi
sampel dan biaya.

85
Bouvier dkk. (2011), melaporkan kadar protein S100B serum selama 3
tahun pertama kehidupan. Pada anak usia 0-16 tahun, rerata kadar protein S100B
adalah 0,17 µg/L (SD 0,11; rentang 0,07-0,83) dan tidak terdapat perbedaan yang
bermakna antara lelaki dan perempuan. Rerata kadar S100B pada anak usia di atas
36 bulan 0,11 µg/L dan pada anak usia 0-3 tahun rerata kadar S100B adalah 0,18
µg/L. Terdapat 4 kelompok usia yang memiliki kadar S100B tidak berbeda
bermakna secara statistik. Rerata kadar S100B secara berurutan sebagai berikut:
usia 0-3 bulan 0,38 µg/L; 4-9 bulan 0,23 µg/L; pada usia 10-24 bulan 0,16 µg/L
dan usia 25-36 bulan 0,11 µg/L. Kadar S100B serum ditemukan menurun seiring
dengan meningkatnya usia dengan rasio korelasi r-=-0,60. Kadar S100Bcss juga
ditemukan berkorelasi dengan lingkar kepala (Bouvier dkk., 2011). Pada studi
studi sebelumnya, dilaporkan bahwa kadar protein S100Bcss meningkat seiring
umur pada individu yang sehat dan diduga tergantung juga pada jenis kelamin
karena peningkatannya lebih bermakna pada anak laki dibanding perempuan.
Portela dkk., 2002), melaporkan bahwa tidak terdapat perbedaan yang bermakna
pada kadar S100Bserum pada laki dan perempuan baik pada neonatus cukup bulan
(median 1,79 µg/L); anak usia 4-9 tahun (median 0,31 µg/L); anak usia 10-15
tahun (0,31 µg/L) dan dewasa (median 0,10 µg/L) (Wiesman dkk., 1998)
menunjukkan bahwa kadar S100B dalam plasma pada usia dewasa (18-65 tahun)
tidak tergantung pada jenis kelamin dan usia.
Kepa dan Grzesik (2013), melakukan studi evaluasi terhadap kadar protein
S100Bcss pada pasien dewasa dengan meningitis bakterialis (purulen). Rerata
kadar protein S100B selama 24 jam pertama dirawat pada pasien dengan kondisi

86
klinis sangat berat adalah 1215,63 pg/mL, sedangkan rerata kadarnya pada pasien
dengan kondisi klinis ringan-sedang adalah 419,56 pg/mL. Tidak terdapat korelasi
antara kadar S100B dengan parameter antiinflamasi yang lain. Hal ini
menunjukkan bahwa protein S100Bcss mungkin berguna untuk memperkirakan
beratnya kondisi klinis pasien (Kepa dan Grzesik, 2013).