karakterisasi fitoplasma penyebab penyakit layu...
TRANSCRIPT

Prasetyo et al. Karakterisasi Fitoplasma 105 J. HPT Tropika. ISSN 1411-7525Vol. 17, No. 2: 105 – 110, September 2017
KARAKTERISASI FITOPLASMA PENYEBAB PENYAKIT LAYU KELAPADI PULAU DERAWAN MENGGUNAKAN RFLP IN SILICO
Agus Eko Prasetyo1, Kikin Hamzah Mutaqin2, & Giyanto2
1Pusat Penelitian Kelapa SawitJl. Brigjend Katamso No 51 Medan 20158
2Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian BogorJl. Kamper Kampus IPB Dramaga, Bogor 16680
E-mail: [email protected]
ABSTRACT
Characterization of phytoplasmas associated with coconut wilt disease in Derawan Island using in silico RFLP. Coconutwilt disease has been reported in Derawan Island that resulted in eradication up to 10% of the total cultivated palms. Theobjective of this study was to detect and characterize phytoplasmas associated with coconut wilt disease in Derawan islandusing nested PCR technique and in silico RFLP based on 16S rRNA gene sequences. Detection of phytoplasmas was performedusing nested PCR method, cloning of nPCR products, sequencing, and analysis of sequencing results using in silico RFLP.The results revealed that phytoplasmas could not be detected by PCR using P1/P7 primer pairs however it could be amplifiedby nested PCR using R16F2n/R16R2 primer pairs resulting amplicon at about 1.25 kb. In silico RFLP analysis indicated thatphytoplasmas associated with coconut wilt disease in Derawan Island belong to 16SrII (witches broom phytoplasma). PCRproduct of the nPCR need to be sequenced because the R16F2n/R16R2 primer will also amplify the other bacterial species,mainly from Gram positive bacteria.
Key words: coconut wilt disease, phytoplasmas, nested PCR, sequencing
ABSTRAK
Karakterisasi fitoplasma penyebab penyakit layu kelapa di Pulau Derawan menggunakan RFLP in silico. Penyakit layukelapa telah ditemukan di Pulau Derawan, Kalimantan Timur yang mengakibatkan 10% tanaman dieradikasi. Penelitian inibertujuan untuk mendeteksi dan mengkarakterisasi fitoplasma yang berasosiasi dengan penyakit layu kelapa di Pulau Derawan.Kegiatan penelitian meliputi pengambilan sampel, ekstraksi DNA, deteksi fitoplasma menggunakan teknik nested-PCR (nPCR),kloning dan sekuensing, serta analisis hasil sekuensing menggunakan RFLP in silico. Hasil penelitian menunjukkan bahwafitoplasma tidak terdeteksi menggunakan PCR dengan primer P1/P7 tetapi menjadi terdeteksi dengan nPCR menggunakanprimer R16F2n/R16R2 yang memiliki amplikon DNA sekitar 1,25 kb. Hasil analisis sekuensing menggunakan RFLP in silicomembuktikan bahwa fitoplasma yang berasosiasi dengan penyakit layu kelapa di Pulau Derawan tergolong ke dalam kelompok16SrII (witches broom phytoplasma). Deteksi fitoplasma memerlukan sekuensing karena primer R16F2n/R16R2 masih dapatmengamplifikasi beberapa bakteri Gram Positif.
Kata kunci: penyakit layu kelapa, fitoplasma, nested-PCR, sekuensing
PENDAHULUAN
Keberadaan tanaman kelapa di Pulau Derawan,Kalimantan Timur, Indonesia mulai berkurang akibatadanya penyakit layu kelapa. Padahal, tanaman inimenjadi satu-satunya tanaman perkebunan yangdibudidayakan. Selain sebagai sumber pendapatantambahan, tanaman kelapa di pulau ini juga memiliki
fungsi estetika yakni menambah kecantikan pandangpantai di pulau yang menjadi maskot pariwisata diprovinsi ini. Terlebih lagi, tanaman kelapa di Indonesiamenjadi komoditi sosial kedua setelah padi dengan luasareal sekitar 3,7 juta ha atau setara dengan sepertigaluas kelapa dunia yang menjadikan Indonesia sebagainegara produsen kelapa terluas di dunia (Anonim, 2007).Kejadian penyakit layu terlihat cukup tinggi, tanaman

106 J. HPT Tropika Vol. 17, No. 2, 2017: 105 -110
mati hingga mencapai 10% sedangkan tanaman lainyang masih bergejala dapat menurunkan produksi buah.
Gejala penyakit layu kelapa di Pulau Derawanditandai dengan menguningnya daun, pelepah-pelepahmaupun daun kelapa yang muncul terlihat lebih pendekdaripada tanaman kelapa yang normal (sehat),mengeringnya daun (klorosis) yang dimulai dari bagiandaun yang tua (bawah), pangkal pelepah tua mudahpatah, terjadinya pengguguran buah kelapa yang masihmuda atau jika masih ada buah kelapa yang tersisasampai besar dan masak, biasanya hanya 1 atau 2 bijisaja. Gejala akhir penyakit ini adalah mengeringnyaseluruh pelepah dan daun kelapa rontok dan terlihathanya seperti tonggak batang kayu. Menurut Leon etal. (1996), tanaman yang sakit akan menghasilkan asamabsisat dan etilen yang sangat tinggi. Akibatnya terjadiketidak-seimbangan hormon yang akan memacupenuaan daun dan mengakibatkan patahnya pelepahserta gugurnya buah yang masih muda (Musetti, 2010).
Sebelumnya, di daerah Sampit, KalimantanTengah ditemukan penyakit layu Kalimantan(kalimantan wilt) yang kemudian berhasil diidentifikasioleh Warokka et al. (2006) disebabkan oleh ca.Phytoplasma oryzae (Warokka et al., 2006). DiMalaysia, ditemukan ca. Phytoplasma cynodontis yangmenjadi penyebab penyakit coconut lethal decline(Nejat et al., 2009). Strain fitoplasma yang telahdiketahui menyebabkan kerugian yang sangat besar padatanaman kelapa di daerah Amerika dan Afrika adalahca. Phytoplasma palmae, penyebab penyakit lethalyellowing (Tymon et al., 1998; Harrison et al., 2002;Myrie et al., 2006). Strain fitoplasma terakhir masukdalam kelas A1 Karantina Tumbuhan Indonesia karenaselain tanaman kelapa, strain ini juga diduga dapatmenyerang tanaman kelapa sawit yang saat ini menjadisumber devisa non migas utama di Indonesia. MenurutWeintraub & Wilson (2010), sampai saat ini, pengendalianpatogen ini pada tanaman kelapa belum ada yangmemuaskan.
Deteksi dan karakterisasi fitoplasma sebagaipenyebab penyakit layu kelapa di Pulau Derawan sangatpenting dilakukan untuk mencegah terjadinya ledakanpenyakit. Penelitian ini bertujuan untuk mengidentifikasidan mengklasifikasikan fitoplasma yang berasosiasidengan penyakit layu kelapa di Pulau Derawanberdasarkan sekuen gen 16S rRNA menggunakanmetode Restriction Fragment Length Polymorphism(RFLP) in silico. Penelitian ini sangat bermanfaatsebagai langkah awal dalam menyusun strategipengendalian penyakit.
METODE PENELITIAN
Tempat dan Waktu. Pengambilan sampel dilakukan diPulau Derawan, Kalimantan Timur kemudian dilanjutkandengan ekstraksi DNA di Laboratorium Bakteriologi,Departemen Proteksi Tanaman, Fakultas Pertanian,Institut Pertanian Bogor. Proses PCR maupun nested-PCR, kloning dan sekuensing dilakukandi Laboratorium Genomic of Bacterial CellFunction, Nara Institute of Science andTechnology (NAIST) Jepang. Analisis RFLP in silicodilakukan di Divisi Proteksi Tanaman, Pusat PenelitianKelapa Sawit, Medan. Penelitian ini dilakukan padaSeptember 2011 hingga September 2013.
Pengambilan Sampel. Pengambilan sampel dilakukandi Pulau Derawan, Kalimantan Timur. Sampel batangtanaman kelapa dibor menggunakan bor kayuberdiameter 1 cm pada ketinggian sekitar 1 meter daripermukaan tanah sedalam 15-20 cm (Oropeza et al.,2002). Kriteria tanaman sampel adalah tanaman dengangejala penyakit ringan (daun menguning dan mulaiklorosis), berat (daun klorosis, mengering dan tidakmemproduksi buah), dan tanaman yang tidak bergejalapenyakit masing-masing 6 tanaman dengan 3 sisipengeboran yang berbeda. Sebagai kontrol positifdigunakan sampel tanaman kelapa terserang penyakitlayu Kalimantan dari daerah Sampit dan beberapasampel tanaman kacang tanah dan kedelai yangterserang penyakit sapu setan dari daerah Bogor.
Ekstraksi DNA dan Amplifikasi menggunakanPCR. DNA fitoplasma dari serbuk kayu kelapadiekstraksi menggunakan metode Zhang et al. (1998).DNA hasil ekstraksi diamplifikasi berturut-turutmenggunakan dua pasang primer universal fitoplasma,P1 (5’-AAG AGT TTG ATC CTG GCT CAG GAT T-3’) (Deng & Hiruki 1991) / P7 (5’-CGT CCT TCATCG GCT CTT-3’) dan dilanjutkan dengan pasanganprimer R16F2n (5’-GAA ACG ACT GCT AAG ACTGG-3’) / R16R2 (5’-TGA CGG GCG GTG TGT ACAAAC CCC G-3’) (Gundersen & Lee, 1996). ReaksiPCR dilakukan pada volume 10 ml yang terdiri dari 1 mltemplate DNA, 1 ml dNTPs (2 mM); 0,6 ml MgSO4(25 mM); 1 ml primer forward (2 pmol); 1 ml primerreverse (2 pmol); 0,1 ml Taq KOD plus Neo; dan 4,3ml dH2O. Siklus PCR yang pertama dilakukan denganmenggunakan pasangan primer P1/P7 dimulai dengansatu siklus pemanasan awal pada suhu 94 oC selama 2menit; dilanjutkan dengan 35 siklus yang terdiri dari

Prasetyo et al. Karakterisasi Fitoplasma 107
denaturasi dengan suhu 94 oC selama 15 detik,penempelan primer dengan suhu 54 oC selama 30 detik,ekstensi dengan suhu 68 oC selama 1 menit 30 detik.Sebanyak 1 ml hasil PCR pertama ini digunakan sebagaitemplate (cetakan) untuk PCR kedua dengan pasanganprimer R16F2n/R16R2. Siklus pemanasan PCR keduasama dengan PCR yang pertama, hanya berbeda padasuhu penempelan primer yakni 57 oC.
Kloning Produk Nested PCR. Terdapat empattahapan kloning, meliputi preparasi sel bakteri kompeten,insersi produk nPCR ke dalam DNA plasmid,transformasi DNA plasmid ke sel bakteri kompeten, danisolasi plasmid. Sel bakteri yang digunakan sebagai selkompeten adalah Eschericia coli DH5α dengan metodemetode Inoue “ultra competent” cells (Sambrook &Russel, 2001). Produk nPCR diinsersikan ke dalamplasmid pDONRTM 201 (Invitrogen) dengan sistemgateway. Plasmid yang telah mengandung produk nPCRkemudian ditransformasikan ke sel bakteri kompetenDH5α dengan metode heat shock (Sambrook &Russell, 2001) dengan seleksi bakteri dilakukan padamedia LB + kanamycin 50 µg/ml. Tahap akhir yaitu isolasiplasmid dilakukan dengan metode alkalin lisis denganSDS (Sambrook & Russell, 2001).
Sekuensing. DNA plasmid yang digunakan sebagaitemplate, sebelumnya telah diencerkan 100–200 kali.Reaksi PCR dilakukan dengan komposisi 1 ml 5X bufersekuensing; 0,5 ml primer F atau R (2 pmol); 2 ml DNAplasmid; 1 ml big dye; dan 0,5 ml dH2O. Mesin PCRdiatur dengan satu siklus denaturasi awal dengan suhu
94 oC selama 2 menit; dilanjutkan dengan 25 siklus yangterdiri dari denaturasi dengan suhu 96 oC selama 10 detik,penempelan primer dengan suhu 50 oC selama 5 detik,ekstensi dengan suhu 60 oC selama 3 menit. Prosessekuensing dilakukan pada mesin sekuenser ABIPRISM 3100 Genetic Analyzer (Applied Biosystem).
Pemotongan Fragmen DNA dengan EnzimRestriksi secara In Silico. Sekuen DNA hasilsekuensing dimasukkan pada program pDRAW32 yangdikembangkan oleh AcaClone Software (http://www.acaclone.com). Masing-masing sekuen DNAkemudian dipotong-potong menggunakan 17 macamenzim restriksi yang baku digunakan untuk analisis RFLPgen 16S rRNA fitoplasma (Wei et al., 2007). Enzim-enzim restriksi tersebut adalah AluI, BamHI, BfaI, BstUI(ThaI), DraI, EcoRI, HaeIII, HhaI, HinfI, HpaI,HpaII, KpnI, Sau3AI (MboI), MseI, RsaI, SspI, danTaqI. Setelah dipotong, kemudian diplotkan denganelektroforesis gel agarose 3%.
HASIL DAN PEMBAHASAN
Deteksi Fitoplasma dengan Nested-PCR. Hasilamplifikasi DNA menggunakan primer universal P1/P7hanya menghasilkan amplikon DNA fitoplasma yangberasal dari kacang tanah dan kedelai pada gel agarose,yang digunakan sebagai kontrol positif. Produkamplifikasi yang dihasilkan adalah sekitar 1,8 kb(Gambar 1) yang di dalamnya terkandung gen yangdekat dengan awal 16S rRNA, daerah interspacer(ITS), dan bagian ujung 5’ gen 23S rRNA. Namun,
3472 bp
19329 bp
925 bp
1882 bp
6223 bp
Gambar 1. Amplifikasi sampel yang diduga terinfeksi fitoplasma menggunakan primer P1/P7 (kiri) dan R16F2n/R16R2 (kanan). Lajur M, l Eco T14I digest ladder; 1-2, sampel kelapa belum bergejala dari PulauDerawan; 3-6, sampel kelapa bergejala ringan dari Pulau Derawan; 7-10, sampel kelapa bergejalaberat dari Pulau Derawan; 11-13, sampel kelapa dari Sampit; 14, sampel kedelai; 15, sampel kacangtanah.

108 J. HPT Tropika Vol. 17, No. 2, 2017: 105 -110
nPCR dari hasil amplikon PCR pertama memperlihatkanadanya fragmen DNA dari sampel kelapa yangberukuran sekitar 1,25 kb yang mengandung internal gen16S rRNA (Gundersen & Lee, 1996).
Semua sampel dari tanaman kelapa bergejala layuringan dan berat (masing-masing 6 tanaman) dari PulauDerawan menunjukkan hasil positif dengan nPCR,sedangkan pada tanaman yang belum bergejala hanya3 tanaman yang positif (sebagian ditunjukkan padaGambar 1). Setelah disekuensing dan dilakukan ujiBLAST dari NCBI, diperoleh bahwa 77,78% (dari total15 tanaman bergejala berat, ringan dan belum bergejala)sekuen DNA produk nPCR sangat mirip denganbeberapa strain fitoplasma dan 23,22% bukan merupakanfitoplasma. Terdeteksinya fitoplasma pada tanamankelapa yang belum bergejala menunjukkan bahwapatogen tersebut telah menginfeksi tanaman namunbelum menimbulkan gejala penyakit. Hal inimembuktikan bahwa metode nPCR dapat digunakanuntuk mendeteksi fitoplasma pada tanaman yang sakitmaupun sehat secara visual. Meskipun demikian,identifikasi fitoplasma memerlukan sekuensing karenaterdeteksinya sekuen DNA bukan fitoplasma denganukuran yang sama sekitar 1,25 kb baik pada tanamansakit maupun sehat.
Sekuen DNA yang mirip dengan spesies bukanfitoplasma meliputi Bacillus megaterium, Bacillus sp.,Clostridium sp. , Friedmaniella lacustris, dan
Lagionella birminghamensis memiliki nilai homologi93-96%, yang termasuk ke dalam bakteri Gram Positif.Menurut Bai et al. (2006), fitoplasma erat berhubungandengan bakteri Gram Positif khususnya grup Bacillusdan Clostridium. Hal ini berarti banyak sekuen genfitoplasma dan bakteri Gram Positif yang akan memilikikesamaan. Terdeteksinya berbagai strain bakteri GramPositif ini juga mengindikasikan bahwa primer fitoplasmayang digunakan belum spesifik.
Hasil uji RFLP in silico dari sampel asal PulauDerawan menunjukkan adanya 6 pola potongan fragmenDNA yang berbeda (Gambar 2). Keenam pola RFLPini diperoleh dari 6 sekuen DNA yang berbeda hasil darisekuensing. Bila dibandingkan dengan hasil RFLP insilico terhadap berbagai grup fitoplasma yang telahdibuat oleh Wei et al. (2007), maka keenam sekuenDNA tersebut mempunyai nilai koefisiensi kesamaanyang tinggi dengan grup 16SrIIA (peanut witches broomphytoplasma) yakni berkisar antara 0,84-0,98 (Tabel1). Kelompok witches broom phytoplasma yangberasosiasi dengan penyakit layu kelapa ini baru pertamakali ditemukan di dunia. Umumnya gejala penyakittanaman yang disebabkan oleh kelompok witchesbroom phytoplasma meliputi daun yang menguning,virescence, memendek dan berbentuk seperti sapu(witches‘broom), kerdil dan bahkan kematian tanaman(Dickinson et al., 2013).
Gambar 2. Ploting hasil analisis RFLP in silico menggunakan 17 enzim restriksi (1-17, berturut-turut AluI, BamHI,BfaI, BstUI, DraI, EcoRI, HaeIII, HhaI, HinfI, HpaI, HpaII, KpnI, Sau3AI, MseI, RsaI, SspI, danTaqI) terhadap 6 sekuen DNA yang berbeda dari Pulau Derawan pada 3% gel agarose. M, markerjX174DNA-HaeIII digestion.

Prasetyo et al. Karakterisasi Fitoplasma 109
SIMPULAN
Deteksi fitoplasma dengan nPCR menggunakanprimer R16F2n/R16R2 menghasilkan fragmen DNAberukuran sekitar 1,25 kb yang diperoleh dari sampeltanaman kelapa bergejala layu berat, ringan, maupunyang belum bergejala. Analisis filogenetik dan RFLP insilico terhadap sekuen DNA dari Pulau Derawanmenggolongkan fitoplasma yang berasosiasi denganpenyakit layu kelapa memiliki hubungan kekerabatanyang tinggi dengan kelompok witches broomphytoplasma (grup 16SrII).
SANCAWANA
Kami mengucapkan terima kasih kepada DirekturPusat Penelitian Kelapa Sawit Medan atas danapenelitian, Kelapa Pusat Penelitian Biologi LIPI Cibinongatas kesempatan magang dan pelaksanaan sebagiankegiatan penelitian, dan Prof. Naotake Ogasawara yangtelah memberikan segala fasilitas penelitian diLaboratorium Genomic of Bacterial Cell Function,Nara Institute of Science and Technology (NAIST)Jepang melalui program JSPS (The Japan Society forPromoting Sciences).
DAFTAR PUSTAKA
Anonim. 2007. Prospek dan Arah PengembanganAgribisnis Kelapa. Ed ke-2. Jakarta: DepartemenPertanian.
Bai XD, Zhang JH, Ewing A, Miller SA, Radek AJ,Shevchenko DV, Tsukerman K, Walunas T,Lapidus A, Campbell JW, & Hogenhout SA. 2006.Living with genome instability, the adaptation ofphytoplasmas to diverse environments of theirinsect and plant hosts. J. Bacteriol. 188(10): 3682–3696.
Deng S & Hiruki C. 1991. Amplification of 16S rRNAgenes from culturable and non culturable mollicutes.J. Microbiol. Methods 14(1): 53–61.
Dickinson M, Tuffen M, & Hodgetts J. 2013. Thephytoplasma: an introduction. In: Dickinson M &Hodgetts J (Eds.) Phytoplasma Methods andProtocols. pp. 1–14. Humana Press, New York.
Gundersen DE & Lee IM. 1996. Ultrasensitive detectionof phytoplasmas by nested PCR assays using twouniversal primer pairs. Phytopathol. Mediterr.35(3): 144–151.
Sekuen fitoplasma 1 2 3 4 5 6 kw p s IIA XIA
Derawan 1 1,00 Derawan 2 0,88 1,00 Derawan 3 0,88 0,86 1,00 Derawan 4 0,92 0,94 0,98 1,00 Derawan 5 0,93 0,92 0,93 0,98 1,00 Derawan 6 0,74 0,72 0,79 0,75 0,76 1,00 Kalimantan wilt 0,76 0,81 0,76 0,75 0,77 0,95 1,00 Peanut 0,92 0,90 0,97 0,99 0,97 0,74 0,75 1,00 Soybean 0,92 0,90 0,97 0,99 0,97 0,74 0,75 1,00 1,00 16SrIIA 0,95 0,95 0,98 0,98 0,96 0,84 0,84 0,99 0,99 1,00 16SrXIA 0,71 0,70 0,72 0,73 0,73 0,74 0,74 0,68 0,68 0,61 1,00
Tabel 1. Koefisien kesamaan yang diperoleh dari analisis pola RFLP in silico gen 16S rRNA dari 6 sekuen DNA asal Pulau Derawan
No 1 – 6: Sekuen Derawan 1 – 6 dari kelapa Pulau Derawan, kw: sekuen kalimantan wilt dari kelapa Sampit, p:sekuen dari kacang tanah Bogor, s: sekuen dari kedelai Bogor, 16SrIIA – 16SrXIA (Wei et al., 2007). Koefisienkesamaannya (F) dihitung untuk masing-masing pasangan sekuen DNA fitoplasma berdasarkan rumus F = (N-x+ Ny)/2Nxy, dengan x dan y merupakan dua strain fitoplasma yang berbeda, Nx dan Ny adalah totaljumlah fragmen DNA yang dihasilkan dari pemotongan menggunakan 17 enzim restriksi, berturut-turut dari strainfitoplasma x dan y, serta Nxy ialah jumlah fragmen DNA dari kedua strain fitoplasma.

110 J. HPT Tropika Vol. 17, No. 2, 2017: 105 -110
Harrison NA, Myrie W, Jones P, Carpio ML, Castilo M,Doyle MM, & Oropeza C. 2002. 16S rRNAinteroperon sequence heterogeneity distinguishesstrain populations of palm lethal yellowingphytoplasma in the Carribean region. Ann. Appl.Biol. 141(2): 183–193.
Leon R, Santamaria JM, Alpizar L, Escamilla JA, &Oropeza C. 1996. Physiological and biochemicalchanges in shoots of coconut palms affected bylethal yellowing. New Phytol. 134(2): 227–234.
Musetti R. 2010. Biochemical changes in plant infectedby phytoplasma. In: Weintraub PG & Jones P(Eds.). Phytoplasmas: Genomes, Plant Hostsand Vectors. pp. 132–146. CAB International,Wallingford.
Myrie WA, Paulraj L, Dollet M, Wray D, & Been BO.2006. First report of lethal yellowing disease ofcoconut palms caused by phytoplasma on NevisIsland. Plant Dis. 90(6): 834.
Nejat N, Sijam K, Abdullah SNA, Vadamalai G, &Dickinson M. 2009. Molecular characterization ofa phytoplasma associated with coconut yellowdecline (CYD) in Malaysia. Am. J. Appl. Sci. 6(7):1331–1340.
Oropeza C, Cordova I, Narvaez M, & Harrison N. 2002.Palm Trunk Sampling for DNA Extraction andPhytoplasma Detection. University of Florida,Florida.
Sambrook J & Russel DW. 2001. Molecular Cloning,A Laboratory Manual. Ed ke-3. Cold SpingHarbor Laboratory Press, New York.
Tymon AM, Jones P, & Harrison NA. 1998. Phylogeneticrelationships of coconut phytoplasmas and thedevelopment of specific oligonucleotide PCRprimers. Ann. Appl. Biol. 132(3): 437–452.
Warokka JS, Jones P, & Dickson MJ. 2006. Detectionof phytoplasma associated with kalimantan wiltdisease of coconut by the polymerase chainreaction. J. Littri 12(4): 154–160.
Wei W, Davis RE, Lee IM, & Zhao Y. 2007. Computer-simulated RFLP analysis of 16S rRNA genes,identification of ten new phytoplasma groups. Int.J. Syst. Evol. Microbiol. 57: 1855–1867.
Weintraub PG & Wilson MR. 2010. Control ofphytoplasma disease and vectors. In: WeintraubPG & Jones P (Eds.). Phytoplasmas: Genomes,Plant Hosts and Vectors. pp. 233–266. CABInternational, Wallingford.
Zhang YP, Uyemoto JK, & Kirkpatrick BC. 1998. Asmall-scale procedure for extracting nucleic acidsfrom woody plants infected with var iousphytopathogens for PCR assay. J. Virol. Methods71(1): 45–50.

Rosyidi et al. Kompatibilitas Cendawan 111
KOMPATIBILITAS CENDAWAN METARHIZIUM ANISOPLIAE (METSCHN.)SOROKIN DENGAN NEMATODA ENTOMOPATOGEN STEINERNEMA SP.
Imron Rosyidi, Hari Purnomo, Nanang Tri Haryadi, & Mohammad Hoesain
Program Studi Agroteknologi, Fakultas Pertanian, Universitas JemberJl. Kalimantan 37, Kampus Tegal Boto, Jember 68121
E-mail: [email protected]
ABSTRACT
Compatibility Metarhizium anisopliae (Metschn.) Sorokin with entomopathogenic nematode Steinernema sp. Metarhiziumanisopliae (Metschn.) Sorokin and entomopathogenic nematode Steinernema sp. can be used as biological control of insectpests. The objective of this research was to identify compatibility between fungus M. anisopliae with entomopathogenicnematode Steinernema sp. This research was conducted using a completely randomized design (CRD) consisting of 5treatments and repeated 5 times. The treatment used was M. anisopliae isolates Jombang 1 with Steinernema sp. (M1), M.anisopliae Jombang 2 with Steinernema sp. (M2), M. anisopliae isolates Kediri with Steinernema sp. (M3), M. anisopliaeisolates Bondowoso with Steinernema sp. (M4), and M. anisopliae isolates Banyuwangi with Steinernema sp. (M5). Basedon this research result, compatibility between the fungus M. anisopliae with entomopathogenic nematode Steinernema sp.was antagonistic. This occurs due to the symbiotic bacteria Xenorhabdus sp. that produced compounds antimycotic andcapable to inhibit the growth of fungus M. anisopliae.
Key words: compatibility, M. anisopliae, Steinernema sp., Xenorhabdus sp.
ABSTRAK
Kompatibilitas cendawan Metarhizium anisopliae (Metschn.) Sorokin dengan nematoda entomopatogen Steinernema sp.Metarhizium anisopliae (Metschn.) Sorokin dan nematoda patogen serangga Steinernema sp. dapat digunakan sebagaiagens pengendali hayati serangga hama. Penelitian ini bertujuan untuk mengetahui kompatibilitas antara cendawan M.anisopliae dengan Steinernema sp. Penelitian ini menggunakan Rancangan Acak Lengkap lima perlakuan dan lima ulangan.Perlakuan yang digunakan yaitu M. anisopliae isolat Jombang 1 dengan Steinernema sp. (M1), M. anisopliae Jombang 2dengan Steinernema sp. (M2), M. anisopliae isolat Kediri dengan Steinernema sp. (M3), M. anisopliae isolat Bondowosodengan Steinernema sp. (M4), dan M. anisopliae isolat Banyuwangi dengan Steinernema sp. (M5). Hasil penelitian menunjukkankompatibilitas antara cendawan M. anisopliae dengan nematoda Steinernema sp. bersifat antagonis. Hal ini terjadi dikarenakanadanya bakteri simbion Xenorhabdus sp. yang menghasilkan senyawa antimikotik dan mampu menghambat pertumbuhancendawan M. anisopliae.
Kata kunci: kompatibilitas, M. anisopliae, Steinernema sp., Xenorhabdus sp.
PENDAHULUAN
Aplikasi kombinasi agens pengendali hayati(APH) ditujukan untuk meningkatkan efektivitaspengendalian, dan sebagai upaya untuk mengurangipenggunaan pestisida dengan ditemukannya metodepengendalian yang lebih ekonomis dan efisien (Xu etal., 2011). Ansari et al. (2008) mengombinasikanMetarhizium anisopliae dengan Steinernema sp.dalam mengendalikan hama Otiorhynchus sulcatusdengan memberikan mortalitas lebih dari 93% pada ujiskala laboratorium dan lebih dari 83% pada uji skalalapang.
Hartati (2013) menyebutkan bahwa penggunaanAPH seperti cendawan entomopatogen M. anisopliaedan nematoda entomopatogen Steinernema sp. banyakdilakukan untuk mengendalikan uret Lepidiota stigmapada tanaman tebu. Penelitian sebelumnya yangdilakukan oleh Alfarizi (2014) mengenai pengendalianuret menggunakan nematoda entomopatogenSteinernema sp. dengan cendawan entomopatogen M.anisopliae di laboratorium, menunjukkan perlakuanaplikasi kombinasi Steinernema sp. dengan M.anisopliae memberikan mortalitas uret sebesar 92%pada pengamatan tiga minggu setelah aplikasi kombinasiSteinernema sp. terlebih dahulu dan 48 jam berikutnya
J. HPT Tropika. ISSN 1411-7525Vol. 17, No. 2: 111 – 118, September 2017

112 J. HPT Tropika Vol. 17, No. 2, 2017: 111 - 118
aplikasi M. anisopliae, sedangkan padapengamatan tigaminggu setelah aplikasi kombinasi M. anisopliae terlebihdahulu dan 48 jam berikutnya aplikasi Steinernema sp.memberikan mortalitas uret sebesar 84%.
Menurut Alfarizi (2014), gejala kematian uret yangditunjukkan pada perlakuan kombinasi Steinernema sp.dengan M. anisopliae sama dengan gejala kematianuret yang disebabkan oleh aplikasi Steinernema sp.secara tunggal dan tidak ditemukan pertumbuhan miseliaM. anisopliae pada tubuh uret yang mati pada perlakuankombinasi Steinernema sp. dengan M. anisopliae. Olehkarena itu pada penelitian ini dilakukan kombinasicendawan entomopatogen M. anisopliae dengannematoda entomopatogen Steinernema sp. untukmengetahui kompatibilitas antara cendawanentomopatogen M. anisopliae dan nematodaentomopatogen Steinernema sp.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilaksanakan diLaboratorium Agroteknologi, Fakultas Pertanian,Universitas Jember pada bulan Januari sampai Agustus2016.
Peremajaan Isolat M. anisopliae. Peremajaandilakukan pada media PDA (Potato Dextrose Agar).Secara aseptis koloni cendawan diambil dan digoreskansecara strike plate pada media PDA dan diinkubasipada ruang gelap dengan suhu rata-rata 28 °C selamakurang lebih 15 hari sampai cendawan entomopatogenM. anisopliae tumbuh memenuhi media. Isolatcendawan M. anisopliae yang digunakan yaitu isolatJombang 1, Jombang 2, Kediri, Banyuwangi danBondowoso. Semua isolat M. anisopliae diisolasi dariserangga inang Lepidiota stigma.
Penyediaan Suspensi Nematoda EntomopatogenSteinernema sp. Nematoda entomopatogenSteinernema sp. diperbanyak dengan carain vivomenggunakan metode perangkap White (White trap)kemudian diinkubasi selama 7-15 hari. Tenebrio molitoryang mati terinfeksi Steinernema sp. dipanen danpopulasi nematoda entomopatogen dihitung padaMikroskop (Olympus SZ51, Tokyo Jepang) dalamCountingdish.
Pembuatan Media NBTA (Nutrient BromothymolBlue Agar). Sebanyak 23 g NA Nutrient Agar (DifroTM
Becton Dickinson and Company, USA) dan 0,025 gBromothymol Blue (ACS Merck, German) dicampur
dengan1 L air destilasi steril, dan dimasak 30 menit laludiautoklaf (Wisd Daihan, Korea) 15 menit suhu 121 oCbertekanan 1 atm, setelah steril didiamkan sampai dingindan ditambahkan 0,04 g TTC (Triphenyl TetrazoliumChloride) (Sigma- Aldrich, Austria USA) yang sudahdisterilisasi menggunakan microdisk (Kurabo, Cina).
Isolasi Bakteri Simbion Xenorhabdus sp. Bakterisimbion Xenorhabdus sp. diisolasi dari larva T. molitorterinfeksi Steinernema sp.. Larva terinfeksi disterilisasipermukaan dengan dicelupkan pada alkohol 70% selama10-15 detik, dibilas tiga kali dengan air destilasi steril,dan dikeringkan pada kertas saring (Whatman, UK).Tungkai T. molitor mati dipotong dan cairanhaemolympha yang keluar dari tungkai digoreskan padamedia NBTA, selanjutnya diinkubasi pada ruang gelapsuhu rata-rata 28 °C selama 24 jam sampai terdapatkoloni bakteri simbion Xenorhabdus sp. fase primeryang tumbuh dengan tanda warna koloni biru.
Uji Kompatibilitas secara Bioassay. Uji kompatibilitasM. anisopliae dengan Steinernema sp. pada larva T.molitor dilakukan dengan dua uji bioassay. Setiap ujibioassay terdapat lima perlakuan yang diulang lima kalidiantaranya M. anisopliae Jombang 1 (M1) denganSteinernema sp., M. anisopliae Jombang 2 (M2)dengan Steinernema sp., M. anisopliae Kediri (M3)dengan Steinernema sp., M. anisopliae Banyuwangi(M4) dengan Steinernema sp. dan M. anisopliae (M5)dengan Steinernema sp.
Pada bioassay pertama T. molitor (n=10 ekor)dicelupkan pada suspensi M. anisopliae (108 spora/ml)selama 10 detik. Selanjutnya diinkubasi selama 48 jampada cawan Petri (Anumbra, Ceko) yang berisi kertassaring (Whatman TM, UK) dan kapas basah untukmengatur kelembapan. Pada periode 48 jam berikutnya,T. molitor (n=10 ekor) diinfeksi dengan 1 ml suspensiSteinernema sp. per cawan Petri (± 200 Juvenil infektif),setelah itu diinkubasi pada ruangan gelap dengan rata-rata suhu ruang 28 °C sampai T. molitor mati terinfeksi,dengan gejala terinfeksi oleh M. anisopliae atauSteinernema sp. secara tunggal ataupun oleh keduanya.
Pada bioassay kedua, T. molitor (n=10 ekor)diinfeksi dengan satu ml suspensi Steinernema sp. (±200 Juvenil infektif) dan diinkubasi selama 48 jam.Setelah 48 jam, T. molitor (n=10 ekor) dicelupkan padasuspensi M. anisopliae (108 spora/ml) selama 10 detik,selanjutnya diinkubasi pada ruang gelap dengan suhuruang 28 °C sampai serangga uji mati terinfeksi.
Pengamatan dilakukan setiap hari, dimulai dari 1hari setelah infeksi (HSI) sampai terdapat mortalitas

Rosyidi et al. Kompatibilitas Cendawan 113
serangga mati 100% pada salah satu perlakuan. Padapengamatan terakhir semua serangga uji dibongkar dandiamati di bawah mikroskop untuk memastikan seranggamati terinfeksi oleh M. anisopliae atau nematodaSteinernema sp.
Interaksi Kompatibilitas Hasil Uji Bioassay.Pengamatan reaksi kompatibilitas pada uji bioassayditentukan berdasarkan rasio mortalitas kenyataandengan mortalitas harapan T. molitor yang dihitungsetelah terdapat mortalitas 100% pada salah satuperlakuan. Data mortalitas kenyataan merupakanmortalitas hasil pengamatan langsung, sedangkanmortalitas harapan merupakan nilai mortalitas yangdiharapkan berdasarkan respon interaksi. Interaksikompatibilitas berdasarkan mortalitas T. molitor ditentukan dengan rumus (Farenhorst et al., 2010):
dengan:Mo = Mortalitas kenyataan;Mi = Mortalitas kontrol infeksi APH pertama secara
tunggal;Mn = Mortalitas kontrol infeksi APH kedua secara
tunggal;Me = Mortalitas harapan.
Jika (Mo–Me) bernilai positif dan nilai X2 > 3,84maka kombinasi antara M. anisopliae denganSteinernema sp. dikatakan sinergis. Jika (Mo -Me)
bernilai negatif dan nilai X2 < 3,84 maka kombinasi antaraM. anisopliae dengan Steinernema sp. dikatakanantagonis (Ansari et al., 2008).
Gejala dan Perkembangan Steinernema sp. padaTubuh T. molitor Mati Terinfeksi. Pengamatandiperoleh dari pengamatan tujuh hari setelah infeksi.Gejala terinfeksi oleh M. anisopliae dijelaskan olehTrizelia at al. (2010) yaitu tubuh larva yang mengerasdan diselimuti oleh hifa cendawan berwarna putih dankemudian berubah menjadi hijau pucat. Sedangkan gejalaserangga terinfeksi Steinernema sp. yaitu perubahanwarna pada T. molitor menjadi cokelat muda sampaicokelat gelap (Chaerani et al., 2007). Pengamatanperkembangan Steinernema sp. dalam tubuh T. molitoryang terinfeksi Steinernema sp. dilakukan denganmembedah tubuh T. molitor untuk memastikan terdapatnematoda yang hidup dan berkembang dalam tubuh T.molitor yang mati terinfeksi.
Uji Kompatibilitas secara In Vitro. Uji kompatibilitassecara in vitro dilakukan antara M. anisopliae denganbakteri simbion Xenorhabdus sp. pada media NBTAsesuai dengan metode Ansari et al. (2005). Ujikompatibilitas secara in vitro menggunakan limaperlakuan dan diulang lima kali. Kelima perlakuan yaituM. anisopliae Jombang 1 (M1) dengan Steinernemasp. , M. anisopliae Jombang 2 (M2) denganSteinernema sp., M. anisopliae Kediri (M3) denganSteinernema sp., M. anisopliae Banyuwangi (M4)dengan Steinernema sp. dan M. anisopliae (M5)dengan Steinernema sp. Potongan subkultur isolat M.anisopliae (D= 0,5 cm) diletakkan pada bagian mediaNBTA pada jarak 3 cm dari tepi cawan Petri dan 3 cm
Gambar 1. Metode uji kompatibilitas secara in vitro; A. sketsa penumbuhan M. anisopliae secara tunggal;B. sketsa penumbuhan M. anisopliae secara tunggal.
)100Mn
-(1 MiMnMe
Me
2)eM-o(M2X
A B
Xenorhabdus sp.
M. anisopliaeM. anisopliae

114 J. HPT Tropika Vol. 17, No. 2, 2017: 111 - 118
dari titik inokulasi bakteri Xenorhabdus sp. BakteriXenorhabdus sp. diambil dengan ose steril dandigoreskan secara garis pada jarak 3 cm dari tepi cawanPetri. Sebagai pembanding dilakukan inokulasi tunggalsemua isolat M. anisopliae tanpa inokulasi bakterisimbion Xenorhabdus sp. pada jarak 3 cm dari tepicawan Petri (Gambar 1). Pengamatan dilakukan interval3 hari setelah investasi (HSI) dan dihentikan setelahmiselium M. anisopliae yang ditumbuhkan secaratunggal menyentuh bagian tepi cawan Petri.
Persentase Hambatan Pertumbuhan M. anisopliae.Persentase hambatan dihitung dari panjang diameter M.anisopliae pada pengamatan terakhir menggunakanrumus persentase hambatan sebagai berikut:
dengan:H% = persentase daya hambat;d1 = diameter pertumbuhan M. anisopliae kontrol;d2 = diameter pertumbuhan M. anisopliae perlakuan.
Indeks Kompatibilitas. Variabel pengamatanmenunjukkan reaksi kompatibilitas antara M. anisopliaedan Xenorhabdus sp. yang ditumbuhkan bersama dalamsatu cawan Petri, apakah terjadi penghambatanpertumbuhan M. anisopliae oleh Xenorhabdus sp. atautidak. Nilai indeks kompatibilitas (IK) dihitungberdasarkan rumus yang diadopsi dan dimodifikasi dariHamilton & Attia (1997).
Jika IK > 1= uji tersebut tidak kompatibel(antagonis) dan jika IK 1= uji tersebut kompatibel(sinergis).
Analisis Data. Hasil pengamatan uji reaksikompatibilitas secara bioasssay dan hambatanpertumbuhan M. anisopliae pada uji kompatibilitassecara in vitro dianalisis ragam menggunakan ANOVAdengan bantuan software StatView versi 5.0.1 SAS(1998) dan data yang menunjukkan berbeda nyata diujilanjut dengan uji Tukey pada taraf 5%.
HASIL DAN PEMBAHASAN
Reaksi Kompatibilitas pada Uji Bioassay. Hasilanalisis ragam kedua uji Bioassay menunjukkan bahwaperlakuan kombinasi antara M. anisopliae denganSteinernema sp. terhadap mortalitas kenyataan T.molitor tidak nyata dengan P-Value= 0,0769 untuk ujibioassay pertama, dan P-Value= 0,3116 untuk uji
bioassay kedua. Berdasarkan rasio nilai mortalitaskenyataan dengan mortalitas harapan pada uji Bioassaypertama pada 4 hari setelah infeksi M. anisopliae yaitumenunjukkan semua perlakuan terjadi interaksiantagonis (Tabel 1), demikian juga pada uji bioassaykedua pada 4 hari setelah infeksi Steinernema sp.(Tabel 2).
Hasil pengataman gejala infeksi T. molitor yangmati baik pada uji bioassay pertama maupun pada ujibioassay kedua menunjukkan tidak ada M. anisopliaeyang berhasil tumbuh pada tubuh T. molitor sampaipengamatan 7 hari setelah perlakuan infeksi pertama.Gejala infeksi yang ditemukan hanya disebabkan olehnematoda entomopatogen Steinernema sp. Hasilpenelitian ini sesuai dengan hasil penelitian Alfarizi(2014), yaitu gejala Lepidiota stigma yang matiterinfeksi pada perlakuan kombinasi M. anisopliaedengan Steinernema sp. hanya disebabkan olehnematoda entomopatogen Steinernema sp. Gejalainfeksi nematoda Steinernema sp. yaitu terjadinyaperubahan warna pada tubuh menjadi cokelat gelap.Chaerani et al. (2007) menyebutkan bahwa T. molitoryang terinfeksi Steinernema sp. menunjukkan gejalakhas yaitu tubuh menjadi cokelat muda sampai cokelatgelap. Hal tersebut mengindikasikan bahwa M.anisopliae dengan Steinernema sp. bersifat antagonissehingga M. anisopliae tidak mampu tumbuh pada tubuhT. molitor setelah nematoda menginfeksi tubuh T.molitor. Setelah tubuh serangga uji T. molitor dibedah,menunjukkan di dalam tubuh T. molitor terdapatnematoda entomopatogen Steinernema sp.. Hal inimenandakan Steinernema sp. berkembang dan hidupdalam tubuh T. molitor yang mati (Gambar 2). Shapiro-Ilan et al. (2004) menyatakan bahwa interaksi antagonispada kombinasi nematoda entomopatogen dengancendawan entomopatogen disebabkan oleh bakterisimbion yang dilepaskan oleh nematoda dalam tubuhserangga yang terinfeksi.
Bakteri Simbion Xenorhabdus sp. Hasil Isolasi.Isolat bakteri Xenorhabdus sp. yang digunakan padauji kompatibilitas secara in vitro merupakan isolat yangdiperoleh dari serangga terinfeksi nematodaentomopatogen Steinernema sp. Koloni bakteriXenorhabdus sp. akan tumbuh seteleh diisolasi darihaemolymph serangga terinfeksi dan diinkubasi selama24 jam pada ruang gelap dengan tanda koloni bakterisimbion Xenorhabdus sp. fase primer berwarna birupada media NBTA (Gambar 3). Menurut Lengyel etal. (2005), bakteri X. budapestensis fase primer padamedia indikator NBTA koloninya berwarna biru.
%100d
d-dH%1
21

Rosyidi et al. Kompatibilitas Cendawan 115
JI= Juvenil Infektif; Mo= Mortalitas kenyataan; Me= Mortalitas harapan yang dihitung berdasarkan rumus: Mn-Mi(1-Mn/100). Untuk interaksi kompatibilitas dilihat berdasarkan rasio mortalitas kenyataan dengan mortalitas harapanpada X2.
Tabel 1. Reaksi kompatibilitas infeksi M. anisopliae dan Steinernema sp. 48 jam berikutnya pada T. molitor padauji bioassay pertama
Isolat M. anisopliae Spora/ ml Nematoda JI/ ml
Interval aplikasi (hari)
Mo (%)
Me (%) X2 Interaksi
Jombang 1 108 Steinernema sp. 200 2 92 95,7 0,14 Antagonis Jombang 2 108 Steinernema sp. 200 2 98 99,3 0,02 Antagonis Kediri 108 Steinernema sp. 200 2 94 96,6 0,22 Antagonis Banyuwangi 108 Steinernema sp. 200 2 96 97,6 0,03 Antagonis Bondowoso 108 Steinernema sp. 200 2 88 92,8 0,25 Antagonis
Tabel 2. Reaksi kompatibilitas infeksi Steinernema sp.dan M. anisopliae 48 jam berikutnya pada T. molitor padauji bioassay kedua
JI= Juvenil Infektif; Mo= Mortalitas kenyataan; Me= Mortalitas harapan yang dihitung berdasarkan rumus: Mn-Mi(1-Mn/100). Untuk interaksi kompatibilitas dilihat berdasarkan rasio mortalitas kenyataan dengan mortalitas harapanpada X2.
Isolat M. anisopliae Spora/ ml Nematoda JI/ ml
Interval aplikasi (hari)
Mo (%)
Me (%) X2 Interaksi
Jombang 1 108 Steinernema sp. 200 2 98 100 0,04 Antagonis Jombang 2 108 Steinernema sp. 200 2 98 100 0,04 Antagonis Kediri 108 Steinernema sp. 200 2 94 100 0,36 Antagonis Banyuwangi 108 Steinernema sp. 200 2 98 100 0,04 Antagonis Bondowoso 108 Steinernema sp. 200 2 96 100 0,16 Antagonis
Gambar 2. Hasil identifikasi T. molitor mati terinfeksi; A. T. molitor terinfeksi nematoda Steinernema sp. dengangejala tubuh serangga terinfeksi berwarna cokelat muda sampai cokelat tua, B. dan C. Pengamatannematoda di bawah mikroskop dari tubuh T. molitor yang dibedah

116 J. HPT Tropika Vol. 17, No. 2, 2017: 111 - 118
Gambar 3. Koloni bakteri simbion Xenorhabdus sp. hasil Isolasi; A. Koloni bakteri simbion Xenorhabdus sp. hasilisolasi, B. Koloni tunggal bakteri simbion Xenorhabdus sp. fase primer, C. Isolat bakteri Xeborhabdussp. murni
Tabel 3. Persentase hambatan isolat M. anisopliae oleh bakteri simbion Xenorhabdus sp.
Perlakuan Rata-rata persentase daya hambat (%)
Met. Jombang 1 dan Xenorhabdus 45,38 Met. Jombang 2 dan Xenorhabdus 38,12 Met. Kediri dan Xenorhabdus 41,54 Met. Banyuwangi dan Xenorhabdus 41,54 Met. Bondowoso dan Xenorhabdus 40,64
Persentase Hambatan Pertumbuhan M. anisopliae.Uji kompatibilitas antara isolat cendawan M. anisopliaedan bakteri simbion Xenorhabdus sp. ini dilakukan untukmengetahui kemampuan bakteri Xenorhabdus sp.menghambat pertumbuhan cendawan M. anisopliae.Hasil analisis ragam menunjukkan bahwa perlakuan asalisolat M. anisopliae tidak nyata mempengaruhi dayahambat bakteri simbion Xenorhabdus sp. dengan P-Value = 0,0758 (Tabel 3). Daya hambat bakteriXenorhabdus sp. terhadap cendawan M. anisopliaepada uji kompatibilitas ini berkisar antara 38,1-45,5%.
Pada pengamatan 9 hari setelah infestasi padauji kompatibilitas secara in vitro diketahui bahwa bakterisimbion Xenorhabdus sp. mampu menghambatpertumbuhan semua isolat M. anisopliae (Gambar 4).Ansari et al. (2005) menyatakan bahwa bakteri simbionXenorhabdus sp. mampu menghambat pertumbuhancendawan entomopatogen. Menurut Dowds (1998),bakteri simbion Xenorhabdus sp. menghasilkansenyawa metabolit sekunder yang bersifat antimikotik.Dijelaskan oleh Li et al. (1995) bahwa senyawaantimikotik yang dihasilkan oleh bakteri simbionXenorhabdus sp. yaitu indoles dan dithiolopyrrolones
(xenorhabdins, xenomins, dan xenorxides). Senyawaantimikotik lainnya yang juga dihasilkan oleh bakteriXenorhabdus sp. yaitu xenocoumacins,hydroxystilbenes, dan nucleosides (Webster et al.,2002).
Indeks Kompatibilitas Isolat M. anisopliae denganBakteri Simbion Xenorhabdus sp. Berdasarkanpengamatan indeks kompatibilitas antara M. anisopliaedengan bakteri simbion Xenorhabdus sp. menunjukkanbahwa semua isolat M. anisopliae yang diuji tidakkompatibel dengan bakteri simbion Xenorhabdus sp.Kriteria indeks kompatibilitas dikatakan tidak kompatibelatau antagonis jika nilai indeks kompatibilitas > 1 dandikatakan kompatibel atau sinergis jika nilai indekskompatibilitas < 1 (Hamilton & Attia, 1997; Hanudin etal., 2012). Nilai indeks kompatibilitas perlakuan darisemua isolat M. anisopliae uji dengan bakteri simbionXenorhabdus sp. berturut-turut 1,9; 1,6; 1,7; 1,7 dan1,7 sehingga dapat dikatakan kompatibilitas antara isolatM. anisopliae dengan bakteri simbion Xenorhabdussp. bersifat antagonis (Tabel 4).

Rosyidi et al. Kompatibilitas Cendawan 117
Tabel 4. Indeks kompatibilitas isolat M. anisopliae dengan Xenorhabdus sp. secara in vitro
SIMPULAN
Berdasarkan hasil penelitian dapat disimpulkanbahwa kompatibilitas antara cendawan M. anisopliaedengan nematoda Steinernema sp. bersifat antagonis,dan bakteri simbion nematoda entomopatogenXenorhabdus sp. mampu menghambat pertumbuhancendawan M. anisopliae.
SANWACANA
Penulis mengucapkan terima kasih kepadaLembaga Penelitian Universitas Jember dengan SkimUsaha Strategi Nasional atas dana hibah penelitian yangdiberikan tahun anggaran 2016 Nomor: 187B/UN25.3.1/LT/2016.
IK > 1= Uji tersebut tidak kompatibel (antagonis); IK < 1= Uji tersebut kompatibel (sinergis).
Perlakuan Diameter Koloni (cm) Indeks Kompatibilitas (IK) Keterangan
Met Jombang 1 6,5 - - Met Jombang 1 × Xenorhabdus 3,54 1,9 Antagonis Met Jombang 2 6,4 - - Met Jombang 2 × Xenorhabdus 3,96 1,6 Antagonis Met Kediri 6,5 - - Met Kediri × Xenorhabdus 3,8 1,7 Antagonis Met Banyuwangi 6,5 - - Met Banyuwangi × Xenorhabdus 3,8 1,7 Antagonis Met Bondowoso 6,3 - - Met Bondowoso × Xenorhabdus 3,74 1,7 Antagonis
Gambar 4. Hasil uji kompatibilitas secara in vitro pada media NBTA; A. Jombang 1 kontrol, B. Jombang 1 denganXenorhabdus sp., C Jombang 2 kontrol, D. Jombang 2 dengan Xenorhabdus sp., E. Kediri kontrol, F.Kediri dengan Xenorhabdus sp., G. Banyuwangi kontrol, H. Banyuwangi dengan Xenorhabdus sp.;I. Bondowoso kontrol, J. Bandowoso dengan Xenorhabdus sp
A B C D E
F G H I J

118 J. HPT Tropika Vol. 17, No. 2, 2017: 111 - 118
DAFTAR PUSTAKA
Alfarizi S. 2014. Pengendalian hayati uret menggunakannematoda patogen serangga (nps) danMetarhizium sp. di laboratorium. Skripsi.Universitas Jember. Jember.
Ansari MA, Shah FA, & Butt TM. 2008. Combined useof entomopathogenic nematodes andMetarhizium anisopliae as a new approach forblack vine weevil, Otiorhynchus sulcatus,control. Entomol. Exp. Appl. 129(3): 340–347.
Ansari MA, Tirry L, & Moens M. 2005. Antagonismbetween entomopathogenic fungi and bacterialsymbionts of entomopathogenic nematodes.Biocontrol 50(3): 465–475.
Chaerani, Suryadi Y, Priyatno TP, Koswanudin D,Rahmat U, Sujatmo, Yusuf, & Griffin CT. 2007.Isolasi nematoda patogen serangga Steinernemadan Heterorhabditis. J. HPT Tropika 7(1): 1–9.
Dowds B. 1998. Bacterial virulence mechanism. In:Simões N, Boemare N, & Ehlers RU (Eds).Entomopathogenic Nematodes. Pp. 9–19.European Commission, Luxembourg.
Farenhorst M, Knols BGJ, Thomas MB, Howard AFV,Takken W, Rowland M, & N’Guessan R. 2010.Synergy in efficacy of fungal entomopathogensand permethrin against west African insecticide-resistant Anopheles gambiae mosquitoes. PlosOne 5(8): 1–10.
Hamilton JT & Attia FL. 1997. Effect of mixture ofBacillus thuringiensis and pesticide on Plutellaxylostella and the parasite Thyraella collaris.J. Econ. Entomol. 70(1): 146–148.
Hanudin B, Marwanto, Hersanti, & Muharam A. 2012.Kompatibilitas Bacillus subtilis, Pseudomonasfluorecens, dan Trichoderma harzianum untukmengendalikan Ralstonia solanacearum padatanaman kentang. J. Hort. 22(2): 172–179.
Hartati S. 2013. Pengendalian Hama Uret padaTanaman Tebu. http://www.puslitgula10.com/2013/05/pengendalian-hama-uret-pada-tanaman-tebu_16.html. Diakses tanggal 10Januari 2017.
Lengyel K, Elke L, Andra´s F, Emilia S, Peter S, & ErkoS. 2005. Description of four novel species ofXenorhabdus, family Enterobacteriaceae:Xenorhabdus budapestensis sp. nov.,Xenorhabdus ehlersii sp. nov., Xenorhabdusinnexi sp. nov., and Xenorhabdus szentirmaiisp. nov. Syst. Appl. Microbiol. 28(2): 115–122.
Li J, Chen G, Webster JM, & Czyzewska E. 1995.Antimicrobial metabolites from a bacterialsymbiont. J. Nat. Prod. 58(7): 1081–1086.
SAS 1998. Using Statview. SAS Institute Inc., USA.
Shapiro-Ilan DI, Jackson M, Reilly CC, & HotchkissMW. 2004. Effects of combining onentomopathogenic fungi or bacterium withentomopathogenic nematodes on mortality ofCurculio caryae (Coleoptera: Curculionidae). J.Biol. Control 30(1): 119–126.
Trizelia, Syam U, & Herawaty Y. 2010. Virulensi isolatMetarhizium sp. yang berasal dari beberaparizosfer tanaman terhadap Crocidolomiapavonana Fabricus (Lepidoptera: Pyralidae).Manggrove 10(2): 51–59.
Webster JM, Chen G, Kaiji-Hu, & Jianxiong-li. 2002.Bacterial metabolites. In: Gaugler R (Ed.).Entomopathogenic Nematology. Pp. 99–114.Cabi Publishing, New York USA.
Xu XM, Jeffries P, Pautasso M, & Jeger MJ. 2011.Combined use of biocontrol agents to manage plantdiseases in theory and practice. J.Phytopathology 101(9): 1024–1031.

Rizkie et al. Kerapatan dan Viabilitas Konidia 119J. HPT Tropika. ISSN 1411-7525Vol. 17, No. 2: 119 – 127, September 2017
KERAPATAN DAN VIABILITAS KONIDIA BEAUVERIA BASSIANA DANMETARHIZIUM ANISOPLIAE PADA MEDIA IN VITRO PH RENDAH
Lilian Rizkie1, Siti Herlinda 1, 2, 3, Suwandi1, 2, 3, Chandra Irsan1, 2, 3,Susilawati1, 3, 4, & Benyamin Lakitan1, 3, 4
1Program Studi Magister Ilmu Tanaman, Fakultas Pertanian, Universitas Sriwijaya, Palembang2Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Sriwijaya, Indralaya
3Pusat Unggulan Riset Pengembangan Lahan Suboptimal (PUR-PLSO), Universitas Sriwijaya, Palembang4)Jurusan Budidaya Pertanian, Fakultas Pertanian, Universitas Sriwijaya, Indralaya
E-mail: [email protected]
ABSTRACT
Conidial density and viability of Beauveria bassiana and Metarhizium anisopliae grown on the low-pH in vitro medium.Liquid bioinsecticide with active ingredient from conidial entomopathogenic fungus has major constraints, namely short shelflife due to declining conidial viability and density is caused by low pH in the bioinsecticide carrier. This experiment aimed tomeasure the loss of conidial viability and density of Beauveria bassiana and Metarhizium anisopliae isolates grown on invitro medium with low pH. Entomopathogenic fungus isolates were used as much as 28 isolates grown on in vitro medium atlow pH, namely pH 5, 4, 3, and 2. The results showed that the fungus isolate that had the highest conidial density on in vitromedium at pH 5 was found on isolates of B. bassiana with code BPcMs (2.583 x 109 conidia mL-1), while the lowest one wasfound on isolates of B. bassiana with code of BWS Pantura (0.825x109 conidia mL-1). All isolate conidial density from in vitromedium with pH 2 decreased regularly. Conidial density of BPcMs isolate decreased to 2.483 x 109conidia mL-1, as well as BWSPantura isolate also decreased to 0.425x109 conidia mL-1. The highest conidial viability at pH 5 was found on isolates of B.bassiana with code of BPcMs (51.572%), while the lowest conidial viability was found on isolate of B. bassiana with BTmPccode (15.040%). At pH 2, almost isolates tested had low conidial viability. The conidial viability of isolates BPcMs decreasedto 47.037%%, while the isolates BTmPc also decreased to 12.778%. Therefore, the lower of the pH of the in vitro medium was,the lower of conidial viability and density of B. bassiana and M. anisopliae was.
Key words: isolate, bioinsecticide, shelf life
ABSTRAK
Kerapatan dan viabilitas konidia Beauveria bassiana dan Metarhizium anisopliae pada media in vitro pH rendah.Bioinsektisida cair berbahan aktif konidia jamur entomopatogen memiliki kendala utama, yaitu singkatnya umur simpan yangdiakibatkan menurunnya viabilitas dan kerapatan konidia yang salah satu penyebabnya adalah penurunan pH media pembawa.Penelitian ini bertujuan untuk mengukur penurunan viabilitas dan kerapatan konidia isolat-isolat Beauveria bassiana danMetarhizium anisopliae yang ditumbuhkan pada media in vitro pada pH rendah. Isolat yang digunakan sebanyak 28 isolatyang ditumbuhkan pada media in vitro pada pH rendah, yaitu pH 5, 4, 3, dan 2. Hasil penelitian menunjukkan pada media invitro pada pH 5 isolat yang memiliki kerapatan tertinggi ditemukan pada isolat B. bassiana kode BPcMs (2,583x109 konidia mL-
1), sedangkan terendah pada isolat B. bassiana kode Bws Pantura (0,825x109 konidia mL-1). Semua isolat pada media in vitropH 2 menurun teratur kerapatan konidianya. Isolat BPcMs menurun kerapatannya menjadi 2,483 x109 konidia mL-1, begitu jugaisolat Bws Pantura turun menjadi 0,425x109 konidia mL-1. Viabilitas konidia pada pH 5 tertinggi ditemukan pada isolat B.bassiana kode BPcMs (51,572%), sedangkan terendah pada isolat B. bassiana kode BTmPc (15,040%). Pada pH 2,hampir semua isolat yang dicobakan mengalami penurunan viabilitas konidia. Viabilitas konidia isolat BPcMs menurunmenjadi 47,037%, sedangkan isolat BTmPc menurun menjadi 12,778%. Dengan demikian, semakin rendah pH media invitro, maka semakin turun viabilitas dan kerapatan konidia isolat B. bassiana dan M. anisopliae.
Kata kunci: isolat, bioinsektisida, umur simpan

120 J. HPT Tropika Vol. 17 No. 2, 2017: 119 - 127
PENDAHULUAN
Jamur entomopatogen, seperti Beauveriabassiana dan Metarhizium anisopliae telah terbuktiefektif (Prayogo, 2014) dan dapat membunuh berbagaispesies serangga hama. Misalnya, wereng cokelat(Nilaparvata lugens) (Herlinda et al., 2008a), werengpunggung putih (Sogatella furcifera) (Herlinda et al.,2008b), Spodoptera litura (Trizelia et al., 2011), kutuputih (Paracoccus marginatus) (Herlinda et al., 2012),penggerek batang padi kuning (Scirpophaga incertulas)(Thalib et al., 2013), Plutella xylostella (Nunilahwati etal., 2013), kutu daun (Aphis gossypii) (Riyanto et al.,2013), jangkrik (Gryllus sp.) (Ardiyati et al., 2015).
Aplikasi bioinsektisida berbahan aktif konidiajamur entomopatogen ini juga tidak berpengaruh burukterhadap arthropoda bukan sasaran (non-targetarthropods). Aplikasi bioinsektisida berbahan aktifkonidia B. bassiana terbukti tidak menurunkankelimpahan laba-laba (Herlinda et al., 2015a).Bioinsektisida tersebut juga tidak menurunkankelimpahan serangga predator di ekosistem sawah(Herlinda et al., 2015b). Hasil penelitian terhadapbioinsektisida berbahan aktif konidia jamurentomopatogen dengan bahan pembawa kompos cairternyata juga menghasilkan manfaat lebih, yaitu selainmengendalikan serangga hama sekaligus dapatmeningkatkan pertumbuhan dan perkembangan tanaman(Herlinda et al., 2015b).
Dari kelebihan-kelebihan di atas, bioinsektisidajuga memiliki banyak kendala dalam penerapannya.Bioinsektisida cair berbahan aktif konidia jamurentomopatogen memiliki kendala utama, yaitu singkatnyaumur simpan yang diakibatkan menurunnya viabilitasdan kerapatan konidianya (Prayogo, 2006; Prayogo &Santoso, 2013). Kemampuan bertahan hidup jamurentomopatogen dapat dipengaruhi suhu, kelambaban,dan faktor lingkungan fisik lainnya (Thalib et al., 2013;Suprayogi et al., 2015). Selain itu, penurunan viabilitasdan kerapatan konidia dipengaruhi oleh penurunan pHmedia pembawa (carrier) (Hanudin et al., 2010).Penurunan pH media pembawa sulit dihindari, semakinlama waktu penyimpanan maka akan semakin turun pH-nya (Indarmawan et al., 2016). Jamur membutuhkanpH optimum berkisar 4-7 untuk pertumbuhannya(Srikandace et al., 2007). Untuk mengatasi permasalahantersebut diperlukan konidia dari isolat yang mampu hidupdi pH rendah.
Seleksi isolat guna mendapatkan isolat yangmampu bertahan di pH rendah perlu dilakukan. Olehkarena itu, penelitian ini bertujuan untuk mengukurpenurunan viabilitas dan kerapatan konidia isolat-isolat
Beauveria bassiana dan Metarhizium anisopliae yangditumbuhkan pada media in vitro pada pH rendah.
METODE PENELITIAN
Tempat dan Waktu. Penelitian ini dilaksanakan diLaboratorium Entomologi, Jurusan Hama dan PenyakitTumbuhan, Fakultas Pertanian, Universitas Sriwijaya.Waktu penelitian dimulai pada bulan Januari sampaiAgustus 2016. Suhu dan kelembapan rata-rata selamapercobaan di laboratorium adalah 29,91 oC dan 90,20%.
Pesiapan Jamur Entomopatogen. Isolat jamurentomopatogen yang digunakan pada penelitian ini adalahisolat B. bassiana dan M. anisopliae yang merupakanisolat koleksi pribadi Siti Herlinda yang dihasilkan daripenelitian-penelitian sebelumnya yang didanai olehKementerian Riset, Teknologi, dan Pendidikan Tinggi.Isolat-isolat tersebut dieksplorasi dari tanah rawa diSumatera Selatan dan dari serangga-serangga yangditemukan di agroekosistem di Sumatera Selatan. Isolat-isolat tersebut selanjutnya ditumbuhkan pada media invitro mengikuti metode Pham et al. (2009) dari mediaagar (Glucose Yeast Agar, GYA) selanjutnya diperbanyakdi dalam media cair, Glucose Yeast Broth (GYB) yangmetode pembuatannya mengikuti Herlinda et al. (2008b),selanjutnya biakan jamur entomopatogen tersebutdigoyang di atas shaker pada kecepatan 120 rpm permenit selama 3x24 jam pada suhu kamar (29,91 oC)dan ruangan terang.
Untuk menghasilkan media in vitro denganberbagai perlakuan pH yang diinginkan pada percobaanini, maka digunakan metode Pham et al. (2009). Untukmendapat variasi pH media in vitro sebesar pH 5, 4, 3,dan 2, maka pada media GYB tersebut diteteskan larutanHCl berkisar 1-4 tetes mikropipet ukuran 10 µL, laludiukur dengan pH Indikator. Setelah, media in vitro berpHsesuai perlakuan selanjutnya isolat-isolat jamurentomopatogen ditumbuhkan pada media tersebut.
Pengamatan Kerapatan dan Viabilitas JamurEntomopatogen. Isolat yang digunakan sebanyak 28isolat yang ditumbuhkan pada media in vitro pada pHrendah, yaitu pH 5, 4, 3, dan 2. Media in vitro yangber-pH rendah ini selanjutnya diinkubasikan selama 3x24jam. Biakan jamur tersebut lalu diamati kerapatankonidianya. Sebanyak 1 mL biakan jamur diambil darimasing-masing isolat ditambah 9 mL air steril, dan prosesini selanjutnya diencerkan hingga 3 kali, seperti yangdilakukan Herlinda (2010). Setelah diencerkan suspensijamur tersebut diamati di bawah mikroskop yangdilengkapi dengan haemocytometer . Metode

Rizkie et al. Kerapatan dan Viabilitas Konidia 121
penghitungan kerapatan konidia mengikuti Gabriel &Riyatno (1989).
Untuk pengamatan viabilitas konidia dilakukanmengikuti cara Herlinda et al. (2010), yaitu suspensi daribiakan jamur tadi yang diencerkan sebanyak 3 kalipengenceran, lalu diambil menggunakan mikropipetukuran 10 µL sebanyak satu tetes yang diteteskan kekaca preparat, lalu ditutup dengan cover glass. Untukmenjaga agar slide preparat tetap basah, maka di pinggircover glass dioleskan kuteks bening. Setelah itu, slidepreparat masing-masing isolat diamati perkecambahankonidianya setiap 24 jam sekali, yaitu pada 24, 48, dan72 jam. Metode penghitungan viabilitas konidia mengikutiGabriel & Riyatno (1989).
Analisis Data. Perbedaan data kerapatan dan viabilitaskonidia antar isolat dianalisis menggunakan AnalisisKeragaman dan perlakuan disusun menggunakanrancangan acak lengkap. Bila terjadi perbedaan antarperlakuan, dilanjutkan dengan uji lanjut menggunakanBeda Nyata Terkecil (BNT) pada taraf nyata 5% yangpenghitungannya dibantu oleh program SAS-STAT padaSAS 6.12.
HASIL DAN PEMBAHASAN
Kerapatan Konidia Jamur Entomoptogen padaMedium in vitro pH Rendah. Hasil penelitianmenunjukkan isolat yang ditumbuhkan pada media invitro pada pH 5 memiliki kerapatan konidia tertinggiditemukan pada isolat B. bassiana berkode BPcMs(2,583 x 109 konidia mL-1), dan tidak berbeda nyatadengan kerapatan konidia isolat BTmTs. Kerapatankonidia terendah ditemukan pada isolat B. bassianaberkode Bws Pantura (0,825 x 109 konidia mL-1) dantidak berbeda nyata dengan kerapatan konidia isolat B.bassiana berkode BTmTf (Tabel 1).
Pada media in vitro pH 4, 3, dan 2, semua isolatmengalami penurunan kerapatan konidia. Penurunankerapatan konidia paling rendah terjadi pada media invitro pH 2 dibandingkan pH lainnya. Pada media in vitropH 2, isolat BPcMs masih mampu memiliki kerapatankonidia sebesar 2,483 x 109 konidia mL-1, dan berbedanyata dengan kerapatan konidia isolat lainnya. Isolat BwsPantura pada media in vitro pH 2 memiliki kerapatankonidia terendah hanya 0,425 x 109 konidia mL-1 danberbeda tidak nyata dengan kerapatan konidia B.bassiana (isolat BTmPe dan BTmTf) dan M. anisopliae(MaMg dan MTmKt).
Umumnya semua isolat pada media in vitromengalami penurunan kerapatan konidia seiringmenurunnya pH media in vitro. Isolat BPcMs menurun
kerapatannya dari 2,583 x 109 konidia mL-1 pada pH 5menjadi 2,542 x 109 konidia mL-1 pada pH 3, dan 2,483x 109 konidia mL-1 pada pH 2. Namun pada pH 4,kerapatan konidia isolat BPcMs menunjukkankecenderungan berbeda, yaitu terjadi peningkatankerapatan konidianya menjadi 2,617 x 109konidia mL-1.Pada isolat Bws Pantura, kerapatan konidianya menurundari 0,825 x 109 konidia mL-1 pada pH 5 menurunmenjadi 0,775 x 109 konidia mL-1 pada pH 4, 0,608 x109 konidia mL-1 pada pH 3, dan 0,425 x 109 konidiamL-1 pada pH 2.
Walau hasil penelitian ini menunjukkan terjadipenurunan kerapatan konidia jamur entomopatogen bilapH medium in vitro diturunkan, namun beberapa isolatmasih tumbuh dan berkembang baik dan menghasilkankerapatan konidia tinggi pada pH rendah. pH yangdigunakan pada penelitian ini merupakan pH di bawahambang toleransi jamur entomopatogen. Jamurentomopatogen tumbuh dan berkembang baik pada pHnetral (pH 6-8) (Karthikeyan et al., 2008; Fan et al.,2011). Dengan ditemukan isolat yang masih dapatmenghasilkan kerapatan konidia yang tinggi pada pH 2pada penelitian ini merupakan harapan untukmenghasilkan bioinsektisida yang memiliki umur simpanyang lebih lama.
Isolat jamur entomopatogen yang mampubertahan hidup dan menghasilkan kerapatan konidia tinggipada pH rendah memiliki keunggulan lebih bila dijadikanbahan aktif bioinsektisida cair. Pertama, bioinsektisidayang mengandung bahan aktif konidia isolat jamurentomopatogen tersebut dapat memiliki umur simpanyang lebih lama sehingga perubahan pH pada bahanpembawa tidak mempengaruhi stabilitas konidia. Kedua,konidia isolat jamur entomopatogen tersebut yang mampubertahan pada bahan pembawa pH rendah dapatmeningkatkan kebugaran konidianya karena pada pHrendah isolat jamur entomopatogen dapat meningkataktivitas enzim kitinasenya. Suryadi et al. (2013)menyatakan bila isolat jamur entomopatogen mampubertahan pada pH rendah, maka isolat jamurentomopatogen tersebut dapat meningkatkanefektifitasnya karena isolat seperti itu memiliki aktivitasenzim kitinase yang tinggi. Jamur entomopatogen yangmampu bertahan pada pH 4 mengalami aktivitas enzimkitinase tertinggi (0,0053 unit mL-1) dibandingkan pH 3.Semakin tinggi pH maka semakin menurun aktivitasenzim tersebut, namun pada pH 9 dan 10 aktivitas enzimkembali meningkat (Suryadi et al., 2013). Semakin tinggiaktivitas enzim kitinase tersebut, maka semakin tinggipula kemampuan isolat jamur entomopatogen tersebutuntuk mematikan serangga hama (Rachmawaty, 2009).

122 J. HPT Tropika Vol. 17 No. 2, 2017: 119 - 127
Viabilitas Konidia Jamur Entomoptogen padaMedium in Vitro pH Rendah. Viabilitas konidia isolatjamur entomopatogen umumnya memilikikecenderungan yang sama pada berbagai pH media invitro. Pada 24 dan 48 jam setelah inkubasi,kecenderungan viabilitas konidia jamur entomopatogen(Tabel 2 dan 3) belum menunjukkan perbedaan yangsignifikan antar isolat, sedangkan setelah diinkubasiselama 72 jam mulai menunjukkan perbedaan viabilitasyang nyata antar isolat (Tabel 4). Pada 72 jam setelahinkubasi, viabilitas konidia jamur entomopatogen padapH 5 ditemukan tertinggi pada isolat B. bassiana berkodeBPcMs (51,572%). Viabilitas konidia terendah pada pH
5 ditemukan pada isolat B. bassiana berkode BTmPc(15,040%). Konsistensi data menunjukkan viabilitaskonidia tertinggi ditemukan pada isolat BPcMs pada pH4 (46,065%), isolat BPcMs pada pH 3 (36,218%), danisolat BPcMs pada pH 2 (47,037%). Namun, viabilitaskonidia terendah ditemukan pada isolat yang berbeda-beda, misalnya viabilitas konidia terendah ditemukanpada isolat B. bassiana berkode BTmPc (15,040%) padapH 5, isolat B. bassiana berkode BTmSm pada pH 4(14,825%), isolat B. bassiana berkode BTmTr pada pH3 (8,333%), dan isolat B. bassiana berkode BTmTr padapH 2 (8,889%) akan tetapi ketiga isolat ini tidak berbedanyata dengan isolat BTmPc (Tabel 4).
Tabel 1. Kerapatan konidia jamur entomopatogen pada berbagai pH media in vitro
Angka diikuti oleh huruf yang sama pada lajur yang sama berbeda tidak nyata, data di dalam kurung merupakandata hasil tranformasi arcsine.
IsolatKerapatan Konidia (1 x 109 konidia/mL)
pH 5 pH 4 pH 3 pH 2
BBY 1,483 (9,170) ef 1,258 (9,097) efg 1,025 (9,010) efgh 0,808 (8,891) defgBLePd 1,233 (9,091) cd 0,992 (8,996) bc 1,042 (9,012) fghi 0,933 (8,969) ghijBPcMs 2,583 (9,411) m 2,617 (9,418) l 2,542 (9,404) p 2,483 (9,395) qBPcPd 1,042 (9,016) b 0,850 (8,928) ab 0,683 (8,830) ab 0,508 (8,700) abBPluS 2,300 (9,361) lm 2,000 (9,300) j 1,633 (9,213) lmn 1,317 (9,111) klmBTmGa 2,183 (9,339) klm 2,025 (9,305) jk 2,017 (9,305) nop 1,750 (9,243) opBTmMa 1,192 (9,072) bc 1,167 (9,062) cde 0,775 (8,887) abcd 0,733 (8,859) deBTmMj 1,683 (9,226) fghi 1,483 (9,169) h 1,475 (9,168) ijklm 0,942 (8,966) fghijBTmPc 1,708 (9,226) fghi 1,558 (9,192) hi 1,483 (9,163) ijklm 1,417 (9,149) klmnoBTmPd 2,100 (9,322) jkl 1,850 (9,267) j 1,717 (9,233) mno 1,500 (9,176) lmnoBTmPe 1,417 (9,151) de 1,000 (9,000) c 0,800 (8,901) bcde 0,533 (8,725) abcBTmRa 1,475 (9,166) ef 1,392 (9,143) fgh 1,100 (9,039) fghi 0,933 (8,970) ghijBTmSm 2,108 (9,324) jkl 1,975 (9,294) j 1,817 (9,259) mno 1,658 (9,219) nopBTmSo 1,475 (9,168) ef 1,208 (9,080) def 0,692 (8,839) ab 0,658 (8,818) cdBTmSr 2,125 (9,327) jkl 1,983 (9,297) j 1,917 (9,282) no 1,567 (9,195) mnoBTmTf 1,083 (9,033) bc 1,042 (9,017) cd 0,725 (8,855) abc 0,483 (8,678) abBTmTk 1,600 (9,198) efgh 1,375 (9,136) fgh 1,025 (9,009) efgh 0,733 (8,861) defBTmTr 1,467 (9,165) ef 1,342 (9,127) efgh 0,900 (8,951) cdef 0,792 (8,891) defgBTmTs 2,433 (9,386) lm 2,358 (9,373) kl 2,158 (9,334) op 2,092 (9,320) pqBws Pantura 0,825 (8,912) a 0,775 (8,883) a 0,608 (8,783) a 0,425 (8,625) aMa 1,883 (9,275) ijk 1,800 (9,254) ij 1,583 (9,196) klmn 1,358 (9,129) klmnMagIn 1,833 (9,263) ghij 1,758 (9,245) ij 1,467 (9,165) ijklm 1,158 (9,064) ijkMAgPd 1,692 (9,227) fghi 1,567 (9,194) hi 1,217 (9,084) hij 0,900 (8,950) efghMaMg 1,433 (9,154) def 1,358 (9,131) efgh 1,017 (8,991) defgh 0,483 (8,682) abMTmJr 1,650 (9,217) efghi 1,258 (9,098) efg 1,192 (9,075) ghij 1,008 (9,003) hijMTmKt 1,533 (9,182) ef 1,400 (9,144) fgh 0,942 (8,965) cdefg 0,567 (8,752) bcMTmMs 1,558 (9,192) efg 1,475 (9,167) gh 1,317 (9,101) hijk 0,925 (8,963) efghiMTmTr 1,858 (9,269) hijk 1,825 (9,261) ij 1,325 (9,122) ijkl 1,183 (9,072) jkl
F Hitung 20,622* 27,696* 18,409* 30,703*P Value 1,78 x 10-26 5,3 x 10-31 8,7 x 10-25 1,2 x 10-32
BNT 5% 0,073 0,070 0,110 0,106
Rizkie et al. Kerapatan dan Viabilitas Konidia 123
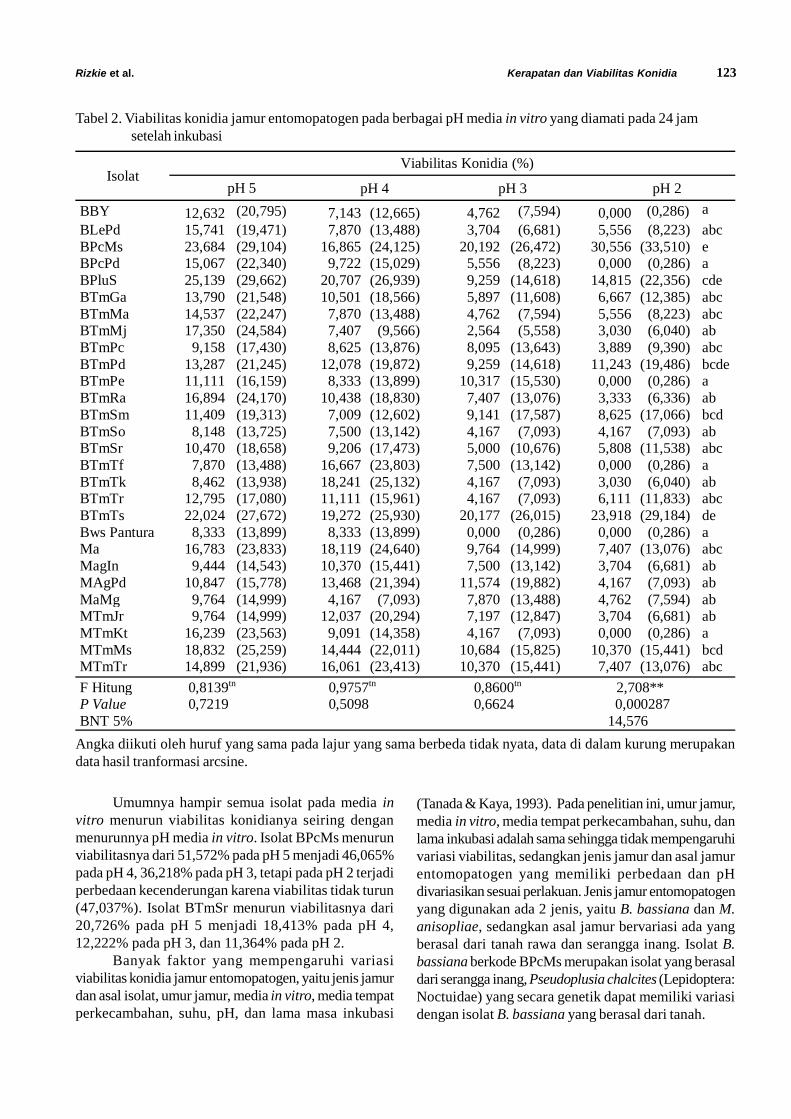
Tabel 2. Viabilitas konidia jamur entomopatogen pada berbagai pH media in vitro yang diamati pada 24 jamsetelah inkubasi
Angka diikuti oleh huruf yang sama pada lajur yang sama berbeda tidak nyata, data di dalam kurung merupakandata hasil tranformasi arcsine.
Umumnya hampir semua isolat pada media invitro menurun viabilitas konidianya seiring denganmenurunnya pH media in vitro. Isolat BPcMs menurunviabilitasnya dari 51,572% pada pH 5 menjadi 46,065%pada pH 4, 36,218% pada pH 3, tetapi pada pH 2 terjadiperbedaan kecenderungan karena viabilitas tidak turun(47,037%). Isolat BTmSr menurun viabilitasnya dari20,726% pada pH 5 menjadi 18,413% pada pH 4,12,222% pada pH 3, dan 11,364% pada pH 2.
Banyak faktor yang mempengaruhi variasiviabilitas konidia jamur entomopatogen, yaitu jenis jamurdan asal isolat, umur jamur, media in vitro, media tempatperkecambahan, suhu, pH, dan lama masa inkubasi
IsolatViabilitas Konidia (%)
pH 5 pH 4 pH 3 pH 2
BBY 12,632 (20,795) 7,143 (12,665) 4,762 (7,594) 0,000 (0,286) a
BLePd 15,741 (19,471) 7,870 (13,488) 3,704 (6,681) 5,556 (8,223) abcBPcMs 23,684 (29,104) 16,865 (24,125) 20,192 (26,472) 30,556 (33,510) eBPcPd 15,067 (22,340) 9,722 (15,029) 5,556 (8,223) 0,000 (0,286) aBPluS 25,139 (29,662) 20,707 (26,939) 9,259 (14,618) 14,815 (22,356) cdeBTmGa 13,790 (21,548) 10,501 (18,566) 5,897 (11,608) 6,667 (12,385) abcBTmMa 14,537 (22,247) 7,870 (13,488) 4,762 (7,594) 5,556 (8,223) abcBTmMj 17,350 (24,584) 7,407 (9,566) 2,564 (5,558) 3,030 (6,040) abBTmPc 9,158 (17,430) 8,625 (13,876) 8,095 (13,643) 3,889 (9,390) abcBTmPd 13,287 (21,245) 12,078 (19,872) 9,259 (14,618) 11,243 (19,486) bcdeBTmPe 11,111 (16,159) 8,333 (13,899) 10,317 (15,530) 0,000 (0,286) aBTmRa 16,894 (24,170) 10,438 (18,830) 7,407 (13,076) 3,333 (6,336) abBTmSm 11,409 (19,313) 7,009 (12,602) 9,141 (17,587) 8,625 (17,066) bcdBTmSo 8,148 (13,725) 7,500 (13,142) 4,167 (7,093) 4,167 (7,093) abBTmSr 10,470 (18,658) 9,206 (17,473) 5,000 (10,676) 5,808 (11,538) abcBTmTf 7,870 (13,488) 16,667 (23,803) 7,500 (13,142) 0,000 (0,286) aBTmTk 8,462 (13,938) 18,241 (25,132) 4,167 (7,093) 3,030 (6,040) abBTmTr 12,795 (17,080) 11,111 (15,961) 4,167 (7,093) 6,111 (11,833) abcBTmTs 22,024 (27,672) 19,272 (25,930) 20,177 (26,015) 23,918 (29,184) deBws Pantura 8,333 (13,899) 8,333 (13,899) 0,000 (0,286) 0,000 (0,286) aMa 16,783 (23,833) 18,119 (24,640) 9,764 (14,999) 7,407 (13,076) abcMagIn 9,444 (14,543) 10,370 (15,441) 7,500 (13,142) 3,704 (6,681) abMAgPd 10,847 (15,778) 13,468 (21,394) 11,574 (19,882) 4,167 (7,093) abMaMg 9,764 (14,999) 4,167 (7,093) 7,870 (13,488) 4,762 (7,594) abMTmJr 9,764 (14,999) 12,037 (20,294) 7,197 (12,847) 3,704 (6,681) abMTmKt 16,239 (23,563) 9,091 (14,358) 4,167 (7,093) 0,000 (0,286) aMTmMs 18,832 (25,259) 14,444 (22,011) 10,684 (15,825) 10,370 (15,441) bcdMTmTr 14,899 (21,936) 16,061 (23,413) 10,370 (15,441) 7,407 (13,076) abc
F Hitung 0,8139tn 0,9757tn 0,8600tn 2,708**P Value 0,7219 0,5098 0,6624 0,000287BNT 5% 14,576
(Tanada & Kaya, 1993). Pada penelitian ini, umur jamur,media in vitro, media tempat perkecambahan, suhu, danlama inkubasi adalah sama sehingga tidak mempengaruhivariasi viabilitas, sedangkan jenis jamur dan asal jamurentomopatogen yang memiliki perbedaan dan pHdivariasikan sesuai perlakuan. Jenis jamur entomopatogenyang digunakan ada 2 jenis, yaitu B. bassiana dan M.anisopliae, sedangkan asal jamur bervariasi ada yangberasal dari tanah rawa dan serangga inang. Isolat B.bassiana berkode BPcMs merupakan isolat yang berasaldari serangga inang, Pseudoplusia chalcites (Lepidoptera:Noctuidae) yang secara genetik dapat memiliki variasidengan isolat B. bassiana yang berasal dari tanah.
124 J. HPT Tropika Vol. 17 No. 2, 2017: 119 - 127
IsolatViabilitas Konidia (%)
pH 5 pH 4 pH 3 pH 2
BBY 18,717 (25,468) 14,286 (25,468) abcd 11,429 (25,468) 4,167 (7,093)BLePd 18,519 (21,225) 12,037 (21,225) ab 7,870 (21,225) 9,722 (15,029)BPcMs 38,670 (38,445) 33,433 (38,445) i 29,006 (38,445) 41,852 (40,300)BPcPd 18,771 (25,225) 15,278 (25,225) abcde 11,111 (25,225) 5,556 (8,223)BPluS 36,944 (37,419) 32,323 (37,419) hi 18,519 (37,419) 18,519 (25,241)BTmGa 25,794 (30,265) 25,946 (30,265) ghi 17,692 (30,265) 13,704 (21,490)BTmMa 21,204 (26,469) 15,741 (26,469) abcde 9,524 (26,469) 11,111 (16,159)BTmMj 19,915 (26,457) 15,278 (26,457) abcde 8,159 (26,457) 9,764 (14,999)BTmPc 11,119 (19,438) 14,219 (19,438) abcd 13,810 (19,438) 9,444 (14,830)BTmPd 18,415 (24,777) 19,870 (24,777) bcdefg 12,963 (24,777) 16,402 (23,714)BTmPe 17,222 (24,177) 16,667 (24,177) abcdef 15,079 (24,177) 4,762 (7,594)BTmRa 22,955 (28,119) 13,468 (28,119) abc 14,815 (28,119) 10,833 (15,852)BTmSm 13,492 (20,760) 12,261 (20,760) a 11,919 (20,760) 11,189 (19,397)BTmSo 16,296 (23,344) 14,537 (23,344) abcd 11,574 (23,344) 8,333 (10,191)BTmSr 15,598 (22,861) 13,810 (22,861) abc 10,000 (22,861) 8,586 (13,977)BTmTf 18,098 (24,904) 20,833 (24,904) cdefg 10,833 (24,904) 5,556 (8,223)BTmTk 17,293 (23,865) 21,574 (23,865) cdefg 7,870 (23,865) 6,061 (8,604)BTmTr 22,559 (23,601) 15,278 (23,601) abcde 8,333 (23,601) 8,889 (14,272)BTmTs 37,202 (37,547) 32,400 (37,547) hi 27,717 (37,547) 35,137 (36,350)Bws Pantura 18,056 (24,933) 13,889 (24,933) abc 3,704 (24,933) 4,762 (7,594)Ma 34,033 (35,633) 22,980 (35,633) efgh 19,461 (35,633) 15,278 (22,767)MagIn 16,111 (22,808) 17,778 (22,808) abcdefg 10,833 (22,808) 7,407 (13,076)MAgPd 17,328 (24,160) 17,172 (24,160) abcdefg 15,278 (24,160) 8,333 (10,191)MaMg 13,468 (21,394) 12,037 (21,394) ab 11,574 (21,394) 9,524 (10,961)MTmJr 16,498 (23,475) 15,741 (23,475) abcde 10,227 (23,475) 7,407 (13,076)MTmKt 18,803 (25,436) 15,455 (25,436) abcde 12,500 (25,436) 5,556 (8,223)MTmMs 34,330 (35,605) 24,815 (35,605) fghi 16,952 (35,605) 17,407 (24,051)MTmTr 17,929 (24,500) 22,424 (24,500) defg 17,407 (24,500) 11,111 (15,961)
F Hitung 1,3396tn 4,0409** 0,682tn 1,478tn
P Value 0,1577 5,08 x 10-7 0,868 0,0914BNT 5% - 6,274 - -
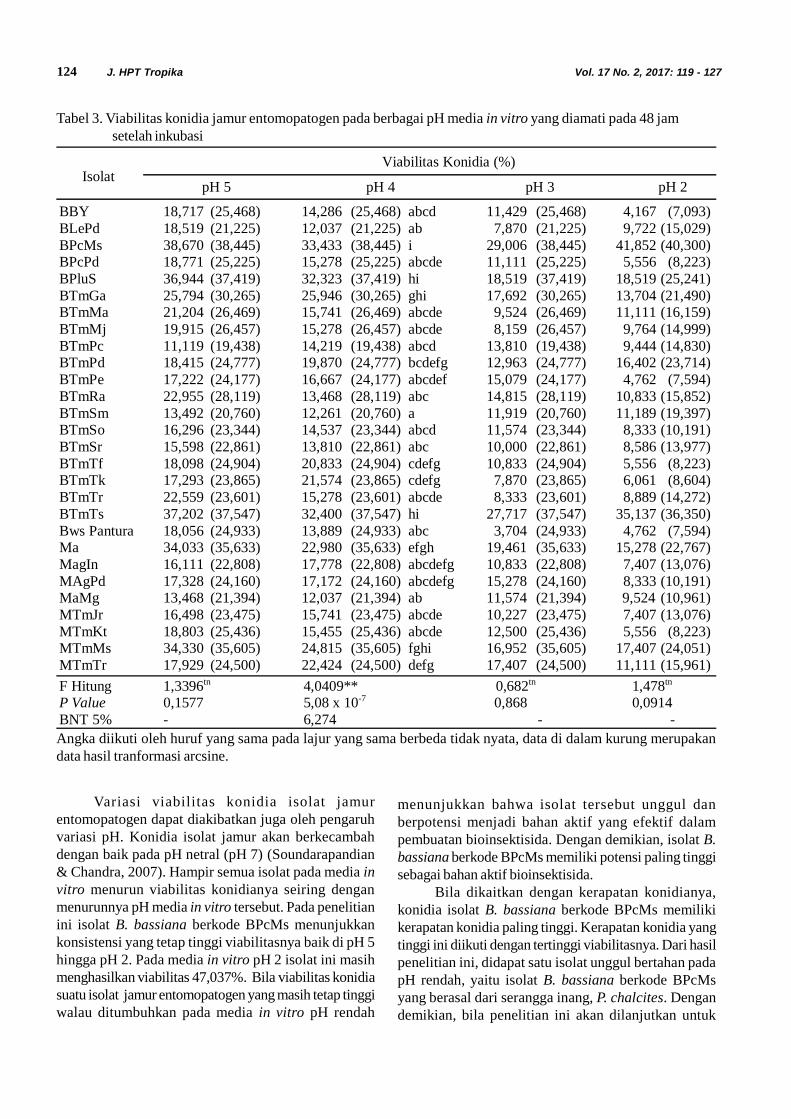
Tabel 3. Viabilitas konidia jamur entomopatogen pada berbagai pH media in vitro yang diamati pada 48 jamsetelah inkubasi
Angka diikuti oleh huruf yang sama pada lajur yang sama berbeda tidak nyata, data di dalam kurung merupakandata hasil tranformasi arcsine.
Variasi viabilitas konidia isolat jamurentomopatogen dapat diakibatkan juga oleh pengaruhvariasi pH. Konidia isolat jamur akan berkecambahdengan baik pada pH netral (pH 7) (Soundarapandian& Chandra, 2007). Hampir semua isolat pada media invitro menurun viabilitas konidianya seiring denganmenurunnya pH media in vitro tersebut. Pada penelitianini isolat B. bassiana berkode BPcMs menunjukkankonsistensi yang tetap tinggi viabilitasnya baik di pH 5hingga pH 2. Pada media in vitro pH 2 isolat ini masihmenghasilkan viabilitas 47,037%. Bila viabilitas konidiasuatu isolat jamur entomopatogen yang masih tetap tinggiwalau ditumbuhkan pada media in vitro pH rendah
menunjukkan bahwa isolat tersebut unggul danberpotensi menjadi bahan aktif yang efektif dalampembuatan bioinsektisida. Dengan demikian, isolat B.bassiana berkode BPcMs memiliki potensi paling tinggisebagai bahan aktif bioinsektisida.
Bila dikaitkan dengan kerapatan konidianya,konidia isolat B. bassiana berkode BPcMs memilikikerapatan konidia paling tinggi. Kerapatan konidia yangtinggi ini diikuti dengan tertinggi viabilitasnya. Dari hasilpenelitian ini, didapat satu isolat unggul bertahan padapH rendah, yaitu isolat B. bassiana berkode BPcMsyang berasal dari serangga inang, P. chalcites. Dengandemikian, bila penelitian ini akan dilanjutkan untuk

Rizkie et al. Kerapatan dan Viabilitas Konidia 125
memformulasikan bioinsektisidanya, maka isolat BPcMsdapat dijadikan calon bahan aktif bioinsektisida tersebut.
SIMPULAN
Pada media in vitro dengan pH 5 isolat yangmemiliki kerapatan tertinggi ditemukan pada isolat B.bassiana kode BPcMs (2,583 x 109 konidia mL-1),sedangkan terendah pada isolat B. bassiana kode BwsPantura (0,825 x 109 konidia mL-1). Semua isolat padamedia in vitro pH 2 menurun secara teratur kerapatankonidianya. Isolat BPcMs menurun kerapatannya
menjadi 2,483 x 109 konidia mL-1, begitu juga isolat BwsPantura turun menjadi 0,425 x 109 konidia mL-1.Viabilitas konidia pada pH 5 tertinggi ditemukan padaisolat B. bassiana kode BPcMs (51,572%), sedangkanterendah pada isolat B. bassiana kode BTmPc(15,040%). Pada pH 2, hampir semua isolat yangdicobakan mengalami penurunan viabilitas konidia.Viabilitas konidia isolat BPcMs menurun menjadi47,037%, sedangkan isolat BTmPc menurun menjadi12,778%. Dengan demikian, semakin rendah pH mediain vitro, maka semakin turun viabilitas dan kerapatankonidia isolat B. bassiana dan M. anisopliae.
Tabel 4. Viabilitas konidia jamur entomopatogen pada berbagai pH media in vitro yang diamati pada 72 jamsetelah inkubasi
IsolatViabilitas Konidia (%)
pH 5 pH 4 pH 3 pH 2
BBY 22,421 (27,848)abcd 21,429 (27,365)abcd 19,524 (21,936) ab 8,929 (14,400)abBLePd 21,296 (22,872)a 19,907 (26,277)abcd 15,741 (19,471) ab 13,889 (18,127)abcBPcMs 51,572 (45,905)e 46,065 (42,740)g 36,218 (36,852) b 47,037 (43,300)cBPcPd 22,475 (27,604)abc 19,444 (26,063)abcd 16,667 (19,882) ab 11,111 (11,946)aBPluS 36,944 (37,419)abcde 35,354 (36,479)efg 25,926 (30,505) ab 22,222 (27,620)abcBTmGa 37,500 (37,587)abcde 43,773 (41,407) fg 26,923 (31,155) ab 17,037 (23,706)abcBTmMa 28,704 (31,490)abcde 19,907 (26,277)abcd 9,524 (10,961) a 16,667 (19,882)abcBTmMj 25,256 (30,085)abcde 19,444 (25,866)abc 14,426 (21,910) ab 12,795 (17,080)abBTmPc 15,040 (22,423)a 16,783 (23,833)ab 19,221 (25,475) ab 12,778 (17,234)abBTmPd 23,543 (28,196)abcd 25,281 (30,120)abcde 15,741 (22,981) ab 21,164 (26,755)abcBTmPe 26,667 (30,857)abcde 20,833 (26,902)abcd 19,841 (21,758) ab 9,524 (10,961)aBTmRa 30,152 (32,986)abcde 17,172 (24,279)abc 19,577 (26,153) ab 17,500 (20,074)abcBTmSm 18,353 (24,528)a 14,825 (22,402)a 11,667 (16,240) ab 13,753 (21,269)abcBTmSo 20,741 (26,058)abc 18,241 (25,132)abc 19,444 (21,850) ab 12,500 (12,778)abBTmSr 20,726 (26,067)abc 18,413 (24,722)abc 12,222 (16,982) ab 11,364 (15,945)abBTmTf 24,158 (28,853)abcd 25,000 (29,489)abcde 14,167 (18,067) ab 11,111 (11,946)aBTmTk 19,858 (25,412)ab 25,741 (30,445)abcde 12,037 (16,586) ab 9,091 (10,685)aBTmTr 25,589 (25,340)ab 18,981 (25,652)abc 8,333 (10,191) a 8,889 (14,272)abBTmTs 48,214 (43,956)de 45,528 (42,432)g 35,623 (36,498) b 37,518 (37,748)bcBws Pantura 23,611 (28,656)abcd 18,056 (24,933)abc 11,111 (15,961) ab 9,524 (10,961)aMa 45,221 (42,228)cde 27,841 (31,823)bcde 26,195 (30,592) ab 22,685 (28,032)abcMagIn 18,889 (25,247)a 21,111 (26,936)abcd 14,167 (18,067) ab 11,111 (15,961)abMAgPd 17,328 (24,160)a 20,202 (26,360)abcd 15,278 (22,767) ab 12,500 (12,778)abMaMg 24,579 (29,543)abcd 15,741 (23,178)a 15,278 (18,752) ab 14,286 (13,822)abMTmJr 22,559 (27,111)abc 19,444 (25,558)abc 17,424 (20,590) ab 11,111 (15,961)abMTmKt 23,932 (28,642)abcd 21,818 (27,762)abcd 16,667 (19,584) ab 11,111 (11,946)aMTmMs 43,932 (41,454)bcde 31,852 (34,207)def 24,858 (29,267) ab 24,444 (28,943)abcMTmTr 23,485 (28,907)abcd 28,788 (32,288)cde 20,741 (22,548) ab 14,815 (18,846)abc
F Hitung 1,298tn 3,920** 0,667tn 0,837tn
P Value 0,184 8,8 x 10-7 0,883 0,692BNT 5% 16,192 8,172 23,014 25,393
Angka diikuti oleh huruf yang sama pada lajur yang sama berbeda tidak nyata, data di dalam kurung merupakandata hasil tranformasi arcsine.

126 J. HPT Tropika Vol. 17 No. 2, 2017: 119 - 127
SANWACANA
Penelitian ini dibiayai oleh Program HibahKompetensi (HIKOM) Tahun Anggaran 2016 sesuaiSurat Direktur Riset dan Pengabdian Masyarakat,Direktorat Riset dan Pengabdian Masyarakat (DRPM),Direktorat Jenderal Penguatan Riset dan Pengembangan,Kementerian Riset, Teknologi, dan Pendidikan Tinggi,Nomor: 0299/E3/2016, tanggal 27 Januari 2016 dengankontrak penelitian Nomor: 023/SP2H/LT/DRPM/II/2016tanggal, 17 Februari 2016 yang penelitiannya diketuaioleh Siti Herlinda.
DAFTAR PUSTAKA
Ardiyati AT, Mudjiono G, & Himawan T. 2015. Ujipatogenisitas jamur entomopatogen Beauveriabassiana (Balsamo) Vuillemin pada jangkrik(Gryllus sp.) (Orthoptera: Gryllidae). Jurnal HPT3(3): 43–51.
Fan Y, Zhang S, Kruer N, & Keyhani NO. 2011. High-throughput insertion mutagenesis and functionalscreening in the entomopathogenic fungusBeauveria bassiana. J. Invertebr. Pathol.106(2): 274–279.
Gabriel BP & Riyatno. 1989. Metarhizium anisopliae(Metch) Sor: Taksonomi, Patologi, Produksi danAplikasinya. Jakarta: Direktorat PerlindunganTanaman Perkebunan, Departemen Pertanian.
Hanudin, Nuryani W, Silvia E, Djatnika I, & MarwotoB. 2010. Formulasi biopestisida berbahan aktifBacillus subtilis, Pseudomonas fluorescens, danCorynebacterium sp. nonpatogenik untukmengendalikan penyakit karat pada krisan. J. Hort.20(3): 247–261.
Herlinda S. 2010. Spore density and viability ofentomopathogenic fungal isolates from Indonesia,and their virulence against Aphis gossypii Glover(Homoptera: Aphididae). Trop. Life Sci. Res.21(1): 11–19.
Herlinda S, Darmawan KA, Firmansyah, Adam T, IrsanC, & Thalib R. 2012. Bioesai bioinsektisidaBeauveria bassiana dari Sumatera Selatanterhadap kutu putih pepaya, Paracoccusmarginatus Williams & Granara De Willink(Hemiptera: Pseudococcidae). J. Entomol.Indones. 9(2): 81–87.
Herlinda S, Mulyati SI, & Suwandi. 2008a. Jamurentomopatogen berformulasi cair sebagaibioinsektisida untuk pengendali wereng coklat.Agritrop 27(3): 119–126.
Herlinda S, SI Mulyati, & Suwandi. 2008b. Selection ofisolates of entomopathogenic fungi and thebioefficacy of their liquid production againstLeptocorisa oratorius nymphs. Microbiol.Indones. 2(3): 141–146.
Herlinda S, Irsan C, Mayasari R, & Septariani S. 2010.Identification and selection of entomopathogenicfungi as biocontrol agents for Aphis gossypii fromSouth Sumatra. Microbiol. Indones. 4(3): 137–142.
Herlinda S, Dewi R, Adam T, Suwandi, & Wijaya A.2015a. Struktur komunitas laba-laba di ekosistempadi ratun: pengaruh aplikasi Beauveria bassiana(Balsamo). J. Entomol. Indones. 12(2): 91–99.
Herlinda S, Kusuma A, Suwandi, & Wijaya A. 2015b.Perbandingan efek pemberian bioinsektisida danekstrak kompos terhadap produksi padi ratun danpopulasi serangga hama. J. Agron. Indones.43(1): 23–29.
Indarmawan T, Mustopa AZ, Budiarto BR, & TarmanK. 2016. Antibacterial activity of extracellularprotease isolated from an algicolous fungusXylaria psidii KT30 against gram-positivebacteria. HAYATI Journal of Biosciences 23: 73–78.
Karthikeyan A, Shanthi V, & Nagasathya A. 2008.Effect of different media and pH on the growthof Beauveria bassiana and its parasitism on leafeating caterpillars. Res. J. Agric. Biol. Sci.4(2):117–119.
Nunilahwati H, Herlinda S, Irsan C, Pujiastuti Y,Khodijah, & Meidelima D. 2013. Uji efikasibioinsektisida jamur entomopatogen berformulasicair terhadap Plutella xylostella (L.) dilaboratorium. J. HPT Tropika 13(1): 52–60.
Pham TA, Kim JJ, Kim SG, & Kim K. 2009. Productionof blastospore of entomopathogenic Beauveriabassiana in a submerged batch culture.Mycobiology 37(3): 218–224.
Prayogo Y. 2006. Upaya mempertahankan keefektifancendawan entomopatogen untuk mengendalikanhama tanaman pangan. J. Litbang Pertanian25(2): 47–54.

Rizkie et al. Kerapatan dan Viabilitas Konidia 127
Prayogo Y. 2014. Efikasi cendawan entomopatogenLecanicillium lecanii terhadap Bemisia tabaci(Hemiptera: Aleyrodidae) pada kedelai. J. HPTTropika 14(2): 187–200.
Prayogo Y & Santoso T. 2013. Viabilitas dan infektivitasformulasi cendawan entomopatogenLecanicillium lecanii sebagai biopestisidapengendalian telur kepik coklat Riptortus linearis.Penelitian Pertanian Tanaman Pangan 32(1): 57–66.
Rachmawaty. 2009. Komparasi enzim kitinase dariBeauveria bassiana galur lokal Sulawesi Selatanterhadap mortalitas ulat grayak (Spodopteralitura). Bionature 10(2): 60–64.
Riyanto, Herlinda S, Irsan C, & Umayah A. 2013.Spesies-spesies jamur entomopatogen yangmenginfeksi Aphis gossypii (Glover) Hemoptera:Aphididae) di agroekosistem sayur dataran rendahdan dataran tinggi Sumatera Selatan. JurnalSainmatika 10(2): 1–9.
Soundarapandian P & Chandra R. 2007. Massproduction of endomopathogenic fungusMetarhizium anisopliae (Deuteromycota:Hyphomycetes) in the laboratory. Res. J.Microbiol. 2(9): 690–705.
Srikandace Y, Hapsari Y, & Simanjuntak P. 2007.Selection of endophytic microbes of Curcumazedoaria in producing antimicrobial compounds.J. Ilmu Kefarmasian Indonesia. 5: 77–84.
Suprayogi, Marheni, & Oemry S. 2015. Uji efektifitasjamur entomopatogen Beauveria bassiana danMetarhizium anisopliae terhadap kepik hijau(Nezara viridula L.) (Hemiptera: Pentatomidae)pada tanaman kedelai (Glycine max L.) di rumahkasa. J. Online Agroekoteknologi 3(1): 320–327.
Suryadi Y, Priyatno TP, Samudra IM, Susilowati DN,Lawati N, & Kustaman E. 2013. Pemurnianparsial dan karakterisasi kitinase asal jamurentomopatogen Beauveria bassiana isolatBB200109. Jurnal AgroBiogen 9(2): 77–84.
Tanada Y & Kaya HK. 1993. Insect Pathology.Academic Press, Inc., San Diego.
Thalib R, Fernando R, Khodijah, Meidalima D, &Herlinda S. 2013. Patogenisitas isolat Beauveriabassiana dan Metarhizium anisopliae asal tanahlebak dan pasang surut Sumatera Selatan untukagens hayati Scirpophaga incertulas. J. HPTTropika 13(1): 10–18.
Trizelia, Syahrawati MY, & Mardiah A. 2011.Patogenisitas beberapa isolat cendawanentomopatogen Metarhizium spp. terhadap telurSpodoptera litura Fabricius (Lepidoptera:Noctuidae). J. Entomol. Indones. 8(1): 45–54.

128 J. HPT Tropika Vol. 17 No. 2, 2017: 128 -136 J. HPT Tropika. ISSN 1411-7525
Vol. 17, No. 2: 128–136 , September 2017
RESPON GENOTIPE KEDELAI BIJI BESAR DAN UMUR GENJAHTERHADAP KOMPLEKS HAMA PENGISAP POLONG
Marida Santi Yudha Ika Bayu, Ayda Krisnawati, & M. Muchlish Adie
Balai Penelitian Tanaman Aneka Kacang dan UmbiJl. Raya Kendalpayak KM. 8 PO BOX 66 Malang, Indonesia
E-mail: [email protected]
ABSTRACT
Response of large seed size and early maturing soybean genotypes against pod sucking bugs complex. Pod sucking bugs(Hemiptera: Pentatomidae) is the most detrimental pest on soybean in Indonesia. These pest attacks caused the decreasing ofquality and quantity of yield as well as decreased seed germination. The use of resistant varieties is an alternative controlwhich is environmentally friendly. The objective of this study was to determine the response of various soybean genotypesagainst pod sucking complex in the field. The experiment was conducted on June to September 2015 in Pasuruan, East Java.The treatment was arranged in randomized block design, 16 treatments with three replicates. The treatment consists of 14soybean promising lines (G 511 H/Anj//Anj///Anj-11-2, G 511 H/Anjasmoro-1-6, G 511 H/Anj//Anj///Anj-7-1, G 511 H/Anjasmoro//Anjasmoro-5-1, G 511 H/Argom//Argom-2-1, G 511 H/Anjasmoro-1-4, G 511 H/Anjasmoro-1-2, G 511 H/Anjasmoro//Anjasmoro-2-8, G 511 H/Arg//Arg///Arg///Arg-12-15, G 511 H/Anj// Anj///Anj-6-3, G 511 H/Arg//Arg///Arg///Arg-19-7, G 511 H/Anjasmoro-1-7, G 511 H/Anj//Anj///Anj////Anjs-6-7, G 511 H/Anjasmoro-1-4-2), and two varieties (Anjasmoro and Grobogan). The resultsshowed that the differences in soybean genotypes significantly influence the intensity of damaged pod and seed caused bypod sucking bugs. The intensity of damaged pod was 29.8–64.0% per plant. The intensity of damaged seed was 13.2-63.9%per plant. The number of puncture was 1.2-1.6 punctures per pod and 0.6-2.7 punctures per seed. Based on the intensity ofdamaged pod and seed, genotipe G 511 H/Anjasmoro//Anjasmoro-2-8, G 511 H/Anjasmoro-1-2, and G 511 H/Anjasmoro-1-6were indicated consistently as moderately resistance against pod sucking complex. Those three genotypes could be use as asource of genes to improve the resistance of soybean against pod sucking bug.
Key words: damage intensity, morphological characters, pod sucking bug, soybean
ABSTRAK
Respon genotipe kedelai biji besar dan umur genjah terhadap kompleks hama pengisap polong. Pengisap polong merupakanhama yang berpotensi merugikan pada pertanaman kedelai di Indonesia. Serangan hama ini mengakibatkan penurunan kualitasdan kuantitas hasil panen, serta menurunnya daya kecambah biji. Penggunaan varietas tahan merupakan alternatif pengendalianyang ramah lingkungan. Tujuan penelitian ini adalah untuk mengetahui respon berbagai genotipe kedelai terhadap serangankompleks pengisap polong di lapang. Penelitian dilaksanakan pada bulan Juni-September 2015 di Kabupaten Pasuruan, JawaTimur. Rancangan percobaan yang digunakan adalah acak kelompok dengan 16 perlakuan dan diulang tiga kali. Perlakuanterdiri dari 14 galur harapan kedelai (G 511 H/Anj//Anj///Anj-11-2, G 511 H/Anjasmoro-1-6, G 511 H/Anj//Anj///Anj-7-1, G 511H/Anjasmoro//Anjasmoro-5-1, G 511 H/Argom//Argom-2-1, G 511 H/Anjasmoro-1-4, G 511 H/Anjasmoro-1-2, G 511 H/Anjasmoro//Anjasmoro-2-8, G 511 H/Arg//Arg///Arg///Arg-12-15, G 511 H/Anj// Anj///Anj-6-3, G 511 H/Arg//Arg///Arg///Arg-19-7, G 511 H/Anjasmoro-1-7, G 511 H/Anj//Anj///Anj////Anjs-6-7, G 511 H/Anjasmoro-1-4-2), dan dua varietas (Anjasmorodan Grobogan). Hasil analisis ragam menunjukkan bahwa perbedaan genotipe kedelai berpengaruh signifikan terhadapintensitas serangan pengisap polong. Intensitas kerusakan polong akibat pengisap berkisar antara 29,8-64,0% per tanaman.Intensitas kerusakan biji akibat pengisap berkisar antara 13,2-63,9% per tanaman. Jumlah tusukan pengisap polong berkisarantara 1,2 – 1,6 buah per polong dan 0,6 – 2,7 buah per biji. Berdasarkan intensitas kerusakan polong dan biji, Genotipe G 511H/Anjasmoro//Anjasmoro-2-8, G 511 H/Anjasmoro-1-2, dan G 511 H/Anjasmoro-1-6 terindikasi konsisten dengan kriteria agaktahan terhadap serangan pengisap polong. Ketiga genotipe tersebut dapat digunakan sebagai sumber gen untuk meningkatkanketahanan varietas kedelai terhadap pengisap polong.
Kata kunci: intensitas kerusakan, karakter morfologi, kedelai, pengisap polong

Bayu et al. Respon Genotipe Kedelai Biji Besar 129
PENDAHULUAN
Pengisap polong (Hemiptera: Alydidae)merupakan hama polyphagus yang menyerang banyakjenis tanaman inang, diantaranya kelompok tanamanbuah, tanaman dari famili Leguminoseae, dan tanamanserealia (Bae et al., 2009; Seo et al., 2011; Panizzi,2013; Mainali et al., 2014). Pada tanaman kedelai,pengisap polong ini menyerang dengan cara menusukdan mengisap bagian polong dan biji kedelai secaralangsung dan tingkat serangannya tinggi pada fasepemasakan polong (Mainali & Lim, 2012; Bae et al.,2014). Dampak yang ditimbulkan sangat serius yaituberupa penurunan kualitas dan kuantitas hasil panen,serta menurunnya daya kecambah biji. Bae et al. (2014)melaporkan bahwa serangan pengisap polong Riptortuspedestris yang terjadi pada fase pengisian polong (R5)hingga panen dapat menyebabkan penurunan hasilhingga 70%. Di Indonesia, spesies pengisap polongkedelai yang paling banyak ditemukan dan sangatmerusak yaitu Riptortus linearis Fabricius, Nezaraviridula Linnaeus, dan Piezodorus hybneri Gmelin(Bayu et al., 2011; Hendrival et al., 2013).
Imago pengisap polong adalah stadia yang palingmerusak tanaman kedelai. Pengisap polong menyerangkedelai dengan cara menusukkan stiletnya pada polongdan mengisap cairan nutrisi yang terkandung pada biji.Serangan yang terjadi pada fase pembentukan polongakan menyebabkan polong kering dan gugur. Pada fasepertumbuhan polong dan perkembangan biji, pengisappolong menyebabkan polong dan biji kempis. Sedangkan,serangan yang terjadi pada fase pengisian biji ataupemasakan polong akan menimbulkan bercak hitamkecoklatan dan penampilan keriput pada biji. MenurutDepieri & Panizzi (2011), waktu penyerangan pengisappolong berkorelasi positif dengan dampak yangditimbulkan. Selain itu, meningkatnya pelukaan pada bijimenyebabkan tertundanya umur masak tanaman(Leonard et al., 2011; Bae et al., 2014). Oleh karenaitu, pengendalian pengisap polong sangat pentingdilakukan hingga menjelang panen untukmempertahankan hasil panen.
Insektisida kimia menjadi pilihan utama bagi petanikedelai pada umumnya untuk menyelamatkanpertanamannya dari serangan hama dan insektisidatersebut sering kali diaplikasikan secara berlebihan(Razaq et al., 2013; Panizzi, 2013). Meskipun telahdiaplikasikan secara intensif, tingkat serangan pengisappolong pada kedelai sering masih tinggi, bahkan petanimasih sering mengalami gagal panen. Aplikasi insektisidasecara terus menerus dapat meningkatkan resistensi
hama target dan membunuh serangga berguna (Cordeiroet al., 2014; Afifah et al., 2015; Haddi et al., 2015;Guedes et al., 2016).
Penanaman varietas tahan hama dalam rangkamengurangi penggunaan insektisida penting untukdipertimbangkan. Penggunaan varietas tahanmerupakan alternatif pengendalian yang ramahlingkungan (Smith & Clement, 2012). Di Indonesia,beberapa genotipe kedelai, diantaranya IAC 100 danG100H, dilaporkan tahan terhadap pengisap polong.Genotipe G100H yang merupakan hasil persilanganantara varietas IAC 100 (introduksi dari Brazilia yangtahan pengisap polong) dengan Himeshirazu (introduksidari Jepang, merupakan salah satu genotipe yangterindikasi tahan pengisap polong) (Bayu & Tengkano,2014). IAC 100 diketahui merupakan hasil persilanganantara varietas IAC-12 and IAC 78-2318, yang tahanterhadap serangga pemakan dan pengisap (Veiga et al.,1999). IAC 100 memiliki pertahanan yang kuat terhadapcekaman yang disebabkan oleh faktor biologi sepertipelukaan oleh hama (Timbo et al., 2014). IAC 100 jugaterindikasi tahan terhadap pengisap polong (McPersonet al., 2007; Suharsono & Sulistyowati, 2012; Bayu &Tengkano, 2014). Selain itu, latar belakang genetiktanaman dan jenis penyebab cekaman mempengaruhirespon pertahanan tanaman terhadap suatu hama(Courtois et al., 2009). Berdasarkan hal-hal tersebut diatas, evaluasi terhadap genotipe kedelai terkait denganketahanannya terhadap pengisap polong sangat pentingdilakukan. Tujuan penelitian ini adalah untuk mengetahuirespon berbagai genotipe kedelai terhadap serangankompleks pengisap polong di lapang.
METODE PENELITIAN
Tempat dan Waktu. Lokasi penelitian berada padaketinggian 124 m di atas permukaan laut (dpl) dengantipe tanah mediteran coklat dan tipe iklim E (agak kering)di Kabupaten Pasuruan, Jawa Timur. Penelitiandilaksanakan pada bulan Juni sampai September 2015.
Rancangan percobaan. Rancangan percobaan yangdigunakan adalah acak kelompok dengan 16 perlakuandan diulang tiga kali. Perlakuan terdiri dari 14 galurharapan kedelai (G 511 H/Anj//Anj///Anj-11-2, G 511H/Anjasmoro-1-6, G 511 H/Anj//Anj///Anj-7-1, G 511H/Anjasmoro//Anjasmoro-5-1, G 511 H/Argom//Argom-2-1, G 511 H/Anjasmoro-1-4, G 511 H/Anjasmoro-1-2,G 511 H/Anjasmoro//Anjasmoro-2-8, G 511 H/Arg//Arg///Arg///Arg-12-15, G 511 H/Anj// Anj///Anj-6-3, G 511H/Arg//Arg///Arg///Arg-19-7, G 511 H/Anjasmoro-1-7,

130 J. HPT Tropika Vol. 17 No. 2, 2017: 128 -136
G 511 H/Anj//Anj///Anj////Anjs-6-7, G 511 H/Anjasmoro-1-4-2) dan dua varietas yaitu Anjasmoro dan Grobogan.
Lokasi percobaan merupakan lahan sawah yangsebelumnya ditanami padi dan sebelum tanam kedelaitidak dilakukan pengolahan lahan. Saluran drainasedibuat sebelum tanam dan dilakukan aplikasi herbisidapada permukaan tanah. Ukuran plot percobaan adalah2,4 x 4,5 m. Penanaman dilakukan dengan jarak tanam40 x 15 cm, dua tanaman per rumpun. Pupuk yangdigunakan adalah 250 kg Phonska ditambah 100 kg SP36 dan satu ton pupuk organik per ha yang diberikanseluruhnya pada saat tanam. Perawatan benih (seedtreatment) dilakukan dengan menggunakan insektisidaberbahan aktif tiametoksam sesuai dosis yangdirekomendasikan. Pengendalian gulma dilakukan padatanaman umur 2 dan 4 minggu setelah tanam (MST).Pengendalian lalat kacang dilakukan denganmenggunakan insektisida berbahan aktif sipermetrin pada8 hari setelah tanam (HST) dengan dosis 1 ml/l,dilanjutkan dengan aplikasi insektisida berbahan aktifsihalotrin pada 14, 21, dan 28 HST untuk mengendalikanhama pemakan daun dengan dosis 2 ml/l. Setelahtanaman berumur 28 hari, tidak dilakukan aplikasiinsektisida dengan tujuan agar populasi pengisap polongdatang ke pertanaman untuk menyerang.
Pengamatan. Pengamatan dilakukan terhadapbanyaknya polong dan biji terserang pengisap polongpada saat panen. Pengambilan sampel dilakukan sebelumpanen sebanyak 5 tanaman per plot secara acak.Pengamatan polong dan biji terserang dilakukan denganmenggunakan mikroskop binokuler di LaboratoriumEntomologi Balai Penelitian Tanaman Aneka Kacangdan Umbi (Balitkabi). Penghitungan persentase intensitaspolong dan biji terserang menggunakan rumus:
Jumlah polong terserang x 100% Jumlah polong diamati
Jumlah biji terserang x 100% Jumlah biji diamati
Untuk menentukan tingkat ketahanan tanamanmenggunakan metode seperti yang dilakukan oleh Asadi(2009) sebagai berikut:Skor 1, serangan 0-20% = tahanSkor 2, serangan 21-40% = agak tahanSkor 3, serangan 41-60% = agak rentanSkor 4, serangan 61-80% = rentanSkor 5, serangan >80% = sangat rentan
Tingkat serangan pengisap polong dianalisismenggunakan ANOVA (SPSS versi 22) dilanjutkandengan uji beda nyata dengan Tukey pada taraf nyata5%.
HASIL DAN PEMBAHASAN
Intensitas polong dan biji terserang pengisappolong. Hasil analisis ragam menunjukkan bahwaperbedaan genotipe kedelai berpengaruh sangat nyataterhadap intensitas serangan pengisap polong padapolong dan biji kedelai (p<0.000) (Tabel 1). Intensitasserangan pengisap pada polong berkisar antara 29,8-64,0% per tanaman. Genotipe kedelai dengan intensitaspolong terserang tertinggi adalah G 511 H/Arg//Arg///Arg///Arg-12-15 yaitu mencapai 64,0% per tanaman.Sedangkan, genotipe dengan intensitas polong terserangterendah adalah G 511 H/Anjasmoro//Anjasmoro-2-8yaitu 29,8% per tanaman.
Berdasarkan intensitas polong terserang yangdikelompokkan menggunakan kategori Asadi (2009),terdapat tiga genotipe kedelai yang terindikasi agak tahanterhadap serangan pengisap polong yaitu G 511 H/Anjasmoro//Anjasmoro-2-8, G 511 H/Anjasmoro-1-6,dan G 511 H/Anjasmoro-1-2 dengan intensitas kerusakanpolong berturut-turut 29,8%, 33,5%, dan 40,1% pertanaman. Selain ketiga genotipe tersebut diatas, genotipe-genotipe yang ditanam di Pasuruan pada MK 2 terindikasiagak rentan dan rentan terhadap pengisap polong.
Intensitas serangan pengisap polong pada biji dari16 genotipe kedelai berkisar antara 13,2-63,9% pertanaman (Tabel 1). Intensitas serangan pengisap padabiji yang terendah terdapat pada varietas Grobogan, yaitu13,2% per tanaman. Sedangkan, intensitas seranganpengisap pada biji yang tertinggi terdapat pada genotipeG 511 H/Anj//Anj///Anj////Anjs-6-7 yaitu mencapai63,9%. Berdasarkan intensitas biji terserang, terdapattiga genotipe kedelai yang terindikasi tahan terhadapserangan pengisap polong yaitu Grobogan, G 511 H/Arg//Arg///Arg///Arg-19-7, dan G 511 H/Anjasmoro//Anjasmoro-2-8 dengan persentase biji terserangberturut-turut 13,2%; 17,9%; dan 19,2% per tanaman.Selain itu terdapat lima genotipe kedelai yang terindikasiagak tahan yaitu genotipe G 511 H/Anjasmoro//Anjasmoro-5-1 (30%/tanaman), G 511 H/Anjasmoro-1-2 (31,8% per tanaman), G 511 H/Anjasmoro-1-6(31,9% per tanaman), G 511 H/Anj//Anj///Anj-11-2(37,1% per tanaman), dan Anjasmoro (39,4% pertanaman).
Pada penelitian ini, intensitas serangan pengisappolong pada polong lebih tinggi bila dibandingkan denganintensitas serangannya pada biji. Hasil penelitian ini
Polong terserang =
Biji terserang =

Bayu et al. Respon Genotipe Kedelai Biji Besar 131
sejalan dengan hasil penelitian Asadi (2009), yaitu bahwapersentase polong terserang dari 17 genotipe yang diujilebih tinggi bila dibandingkan dengan persentase bijiterserang oleh pengisap polong. Alasan yangmemungkinkan untuk menjelaskan fenomena ini adalahkekerasan kulit polong. Stilet pengisap diduga hanyamampu menjangkau kulit polong dan tidak mengenaipermukaan biji karena kulit polong keras dan adanyarongga diantara kulit polong dengan biji. Suharsono &Sulistyowati (2012) melaporkan bahwa ketebalan dankekerasan kulit polong sangat nyata menurunkan tingkatserangan pengisap polong.
Perbedaan tingkat serangan pengisap pada polongdan biji beberapa genotipe kedelai pada penelitian inijuga diduga karena adanya keragaman stadia pengisappolong yang menyerang. Selain kepadatan populasi hamadan stadia pertumbuhan tanaman, keragaman tingatserangan pengisap polong dipengaruhi oleh stadiaperkembangan hama (Lourencao et al., 2010; Depieri& Panizzi, 2011). Menurut Arifin & Tengkano (2010),tingkat kerusakan polong dan biji akibat serangan P.
hybneri mulai terlihat oleh stadia nimfa instar 3, 4, dan5, serta imago pada 63 hari setelah tanam (HST). Depieri& Panizzi (2011) juga melaporkan bahwa waktupenyerangan pengisap polong berkorelasi positif dengandampak yang ditimbulkan. Nimfa P. hybneri instar 1tidak bergerak dan nimfa instar 2 memiliki perilakukurang gesit sehingga tingkat kerusakan yangditimbulkan masih sangat rendah (Arifin & Tengkano,2010). Hal tersebut menguatkan dugaan bahwa nimfahanya mampu mengisap kulit polong dan tidak melukaibiji. Untuk dapat menyimpulkan secara lebih akurat,pengamatan populasi hama dan stadia yang menyerangperlu dilakukan pada penelitian berikutnya.
Penentuan tingkat ketahanan genotipe kedelaiterhadap pengisap polong pada penelitian ini didasarkanpada kekonsistensian dari intensitas serangannya padapolong dan biji kedelai. Genotipe G 511 H/Anjasmoro//Anjasmoro-2-8, G 511 H/Anjasmoro-1-2, dan G 511 H/Anjasmoro-1-6 adalah genotipe-genotipe yangmenunjukkan konsistensi dengan kategori agak tahanterhadap serangan pengisap polong berdasarkan polong
Tabel 1. Tingkat serangan pengisap polong pada 16 genotipe kedelai di Kabupaten Pasuruan
Angka selajur yang diikuti oleh huruf yang sama menyatakan tidak berbeda nyata (Tukey’s HSD test, p < 0,05)
Genotipe Polong
terserang (%/tanaman)
Kriteria ketahanan
Biji terserang (%/tanaman)
Kriteria ketahanan
G 511 H/Anj//Anj///Anj-11-2 40,5 abc AR 37,1 abcdef AT
G 511 H/Anjasmoro-1-6 33,5 bc AT 31,9 bcdef AT
G 511 H/Anj//Anj///Anj-7-1 54,7 abc AR 51,7 abc AR
G 511 H/Anjasmoro//Anjasmoro-5-1 44,8 abc AR 30,0 cdef AT
G 511 H/Argom//Argom-2-1 57,8 ab AR 53,7 abc AR
G 511 H/Anjasmoro-1-4 59,2 ab AR 57,1 abc AR
G 511 H/Anjasmoro-1-2 40,1 abc AT 31,8 bcdef AT
G 511 H/Anjasmoro//Anjasmoro-2-8 29,8 c AT 19,2 def T
G 511 H/Arg//Arg///Arg///Arg-12-15 64,0 a R 62,0 ab R
G 511 H/Anj//Anj///Anj-6-3 52,5 abc AR 48,1 abcde AR
G 511 H/Arg//Arg///Arg///Arg-19-7 46,4 abc AR 17,9 ef T
G 511 H/Anjasmoro-1-7 58,6 ab AR 52,6 abc AR
G 511 H/Anj//Anj///Anj////Anjs-6-7 62,8 a R 63,9 a R
G 511 H/Anjasmoro-1-4 52,9 abc AR 49,9 abcd AR
Anjasmoro 44,8 abc AR 39,4 abcdef AT
Grobogan 62,0 a R 13,2 f T
F 4,243 7,034
df 15, 32 15, 32
p p < 0,000 p < 0,000

132 J. HPT Tropika Vol. 17 No. 2, 2017: 128 -136
dan biji terserang. Kekonsistensian kategori ketahananberdasarkan polong dan biji terserang dinilai lebih kuatdalam pemilihan galur tahan terhadap serangan hamadibandingkan dengan pemilihan yang hanya didasarkanpada biji terserang saja. Biji terserang secara langsunglebih berpengaruh terhadap hasil, baik kualitas maupunkuantitas. Menurut Leonard et al. (2011), pelukaan padabiji akibat serangan hama menyebabkan umur masaktanaman menjadi tertunda. Namun, polong terserangyang ditandai dengan adanya bercak hitam pada kulitpolong akan mempengaruhi ketertarikan konsumendalam memilih kedelai segar.
Intensitas polong dan biji terserang pada genotipeG 511 H/Anjasmoro//Anjasmoro-2-8, G 511 H/Anjasmoro-1-2, dan G 511 H/Anjasmoro-1-6 lebihrendah dibandingkan dengan genotipe lainnya. Ketigagenotipe ini kurang terpilih oleh nimfa dan imago yangmerupakan stadia pengisap polong yang paling merusakkedelai untuk mengisap cairan atau nutrisi yang terdapatpada polong dan biji kedelai tersebut. Sifat ketahananyang dimiliki oleh ketiga genotipe tersebut adalahketahanan antixenosis. Antixenosis adalah mekanismeketahanan yang ditunjukkan oleh tanaman inang, yangmenghalangi serangga untuk meletakkan telur, makan,mencari tempat berlindung, dan kolonisasi (Afzal et al.,2009; Morando et al., 2015). Ketahanan antixenosistergolong sangat efektif karena dapat menyebabkanserangga kelaparan dan mati karena tidak mendapatsumber makanan.
Furstenberg-Hagg et al. (2013) dan War et al.(2012) melaporkan bahwa pertahanan tanaman terhadapserangan serangga herbivora terdiri dari pertahananlangsung yang meliputi trikoma (bulu pada tanaman),produksi metabolit primer (protein inhibitor dan enzimantioksidan), dan produksi metabolit sekunder (asamfenolik, flavonoid, dan isoflavon). Trikoma serta ketebalankulit polong menjadi faktor penghalang utama bagipengisap polong untuk menusukkan stiletnya (Souza etal., 2012). Selain itu, tanaman juga memiliki pertahanantidak langsung melalui produksi senyawa organik yangmudah menguap seperti metil salisilat dan cis-jasmoneyang dapat menarik musuh alami untuk datang sehinggaserangga akan menjauhi tanaman tersebut (Courtois etal., 2009; Michereff et al., 2011; Colazza & Wajnberg,2013).
Pada Gambar 1 dapat dilihat adanya keeratanhubungan antara polong terserang dengan biji terserangpengisap polong dengan nilai r = 0,666, artinyabanyaknya polong yang terserang akan diikuti denganbanyaknya jumlah biji yang terserang. Namun, padapenelitian lapang ini, hubungan antara keduanya tidakbegitu erat. Karakteristik polong memiliki peran pentingdalam menghalangi kerusakan biji. Pada kasus yangdemikian, stilet dari pengisap polong banyak yang hanyamelukai kulit polong dan tidak menjangkau bagian biji.
Jumlah tusukan pengisap polong pada polong danbiji. Hasil analisis ragam menunjukkan bahwaperbedaan genotipe kedelai tidak berpengaruh nyata
Gambar 1. Keeratan hubungan antara polong terserang (%) dengan biji terserang (%) pengisap polong pada 16genotipe kedelai
r = 0,666P = 0,004

Bayu et al. Respon Genotipe Kedelai Biji Besar 133
terhadap jumlah tusukan pengisap polong pada polong(p<0,507) tetapi berbeda nyata terhadap jumlah tusukanpada biji (p<0,036). Jumlah tusukan pengisap polongberkisar antara 1,2-1,6 buah per polong dan 0,6-2,7 buatper biji. Jumlah tusukan pada polong yang tertinggiterdapat pada varietas Grobogan (1,6 buah) dan yangterendah terdapat pada genotipe G 511 H/Argom//Argom-2-1 dan G 511 H/Anjasmoro-1-4 masing-masinghanya 1,2 buah. Jumlah tusukan pada biji yang tertinggiterdapat pada genotipe G 511 H/Anjasmoro-1-6 dan G511 H/Anj//Anj///Anj////Anjs-6-7 yaitu masing-masingsebanyak 2,7 buah. Sedangkan jumlah tusukan pada bijiyang terendah terdapat pada varietas Grobogan yaituhanya 0,6 buah.
Varietas Anjasmoro dan Grobogan diketahuisebagai varietas yang banyak diminati oleh petani karenahasil panennya yang tinggi dan sesuai untuk bahan bakutahu dan tempe (Krisnawati et al., 2016). Berdasarkanpengelompokan kriteria ketahanan yang mengacu padaAsadi (2009), varietas Anjasmoro dan Groboganmenunjukkan ketidakkonsistenan kriteria ketahananantara polong terserang dan biji terserang di lapang.Dilihat dari intensitas polong terserang, kedua varietastersebut tergolong rentan. Sedangkan, berdasarkanintensitas biji terserang keduanya tergolong tahanterhadap pengisap polong. Pada varietas Grobogan,jumlah tusukan yang ditemukan pada polong tercatatpaling banyak namun tusukan yang ditemukan pada bijitercatat paling sedikit. Hal ini terjadi karena stiletpengisap polong hanya melukai bagian permukaan polongvarietas Grobogan dan banyak dari tusukan tersebut yangtidak mencapai permukaan biji. Bagi petani yangmengambil hasil biji, tentunya keadaan tingginyaintensitas polong terserang ini bukan menjadipermasalahan jika didukung dengan intensitas kerusakanyang rendah pada biji. Meskipun demikian, penelitianlebih lanjut diperlukan untuk mengkonfirmasi hasilpenelitian ini terutama pada kondisi dipaksakan ataupengujian dengan tanpa pilihan inang.
Pengisap polong merusak tanaman kedelai dengancara menusuk dan mengisap polong serta biji kedelai.Polong dan biji yang terserang pengisap polong akanmenunjukkan gejala berupa bercak atau titik hitam bekastusukan stilet. Bercak hitam pada polong tersebutnampak jelas bila dilihat dari bagian permukaan dalamkulit polong. Serangan yang berat akan menyebabkantusukan stilet pada polong dapat menembus hingga kepermukaan biji. Serangan terjadi pada saat pengisianbiji, dapat menyebabkan biji tidak berkembang karenajaringan/sel yang mati berhenti berkembang.
Karakter Agronomi. Preferensi pengguna terhadapvarietas kedelai masa kini, selain berdaya hasil tinggi,juga berumur masak genjah (< 80 hari), berukuran bijibesar (> 14 g/100 biji) dan juga relatif toleran terhadapsalah satu cekaman biotik maupun abiotik. Pengisappolong memiliki sebaran cukup luas di Indonesia danmenimbulkan kerusakan pada pertanaman kedelai padaberbagai musim tanam. Tersedianya varietas kedelaiyang mampu memenuhi preferensi pengguna dan relatiftahan terhadap pengisap polong penting untukpengembangan kedelai di daerah tropis Indonesia.
Sidik ragam terhadap umur masak, bobot 100 bijidan hasil biji dari 16 genotipe kedelai, diperoleh pengaruhperlakuan yang nyata untuk ketiga karakter (Tabel 2)menunjukkan terdapat perbedaan karakter agronomi dari16 genotipe kedelai. Rentang umur masak adalah 75 –81 hari (rata-rata 76 hari), bobot 100 biji antara 15,22 –19,73 g (rata-rata 16,62 g/100 biji) dan rentang hasil bijiantara 2,01-2,55 ton/ha (rata-rata 2,30 ton/ha) (Tabel3).
Genotipe yang menunjukkan konsisten agak tahanterhadap hama pengisap polong berdasarkan kerusakanpada polong maupun biji yaitu Genotipe G 511 H/Anjasmoro//Anjasmoro-2-8, memiliki bobot 100 biji 16,87g dengan potensi hasil 2,24 ton/ha dan umur masak 74hari. Selain itu, terdapat genotipe G 511 H/Anjasmoro-1-2 memiliki umur masak 76 hari, bobot 100 biji 15,26 g
Tabel 2. Sidik ragam umur masak, bobot 100 biji dan hasil biji dari 16 genotipe kedelai. MK 2, 2015
tn = tidak nyata, ** = nyata pada p = 0,01
Karakter Kuadrat tengah
Ulangan Perlakuan
Umur masak (hr) 0,5208 tn 8,9958 ** Bobot 100 biji (g) 6,2335 ** 4,9187 ** Hasil (t/ha) 0,0237 tn 0,1691 **

134 J. HPT Tropika Vol. 17 No. 2, 2017: 128 -136
dengan potensi hasil 2,14 ton/ha, dan G 511 H/Anjasmoro-1-6 memiliki umur masak 75 hari, bobot 100biji 17,08 g dengan potensi hasil 2,46 ton/ha (Tabel 3).Ketiga genotipe tersebut memiliki harapan untukmemenuhi preferensi pengguna kedelai saat ini diIndonesia. Ketiga genotipe tersebut dapat digunakansebagai sumber gen untuk meningkatkan ketahananvarietas kedelai terhadap pengisap polong.
SIMPULAN
Genotipe G 511 H/Anjasmoro//Anjasmoro-2-8, G511 H/Anjasmoro-1-2, dan G 511 H/Anjasmoro-1-6menunjukkan tingkat serangan yang lebih rendah biladibandingkan dengan genotipe lainnya dan terindikasikonsisten agak tahan pengisap polong kompleksberdasarkan intensitas serangan pada polong dan biji dilapangan. Karakter polong pada kedelai memiliki peranpenting dalam mengendalikan kerusakan biji oleh hamapengisap polong. Oleh karena itu, strategi peningkatanketahanan kedelai terhadap pengisap polong dapatdilakukan dengan memperbaiki karakter kulit polongmelalui rekayasa genetika.
SANWACANA
Tabel 3. Karakter umur masak, bobot 100 biji dan hasil biji dari 16 genotipe kedelai. MK2, 2015
Terima kasih kami sampaikan kepada BadanPenelitian dan Pengembangan Pertanian yang telahmemberikan dana melalui DIPA untuk terlaksananyapenelitian ini hingga selesai serta kepada Arifin yangtelah membantu pelaksanaan penelitian di lapang hinggaselesai.
DAFTAR PUSTAKA
Afifah L, Hidayat P, Buchori D, Marwoto, & RahardjoBT. 2015. Pengaruh perbedaan pengelolaanagroekosistem tanaman terhadap strukturkomunitas serangga pada pertanaman kedelai diNgale, Kabupaten Ngawi, Jawa Timur. J. HPTTropika 15(1): 53–64.
Afzal M, Nazir Z, Bashir MH, & Khan BS. 2009.Analysis of host plant resistance in some genotypesof maize against Chilo partellus (Swinhoe)(Pyralidae: Lepidoptera). Pak. J. Bot. 41(1): 421–428.
Arifin M & Tengkano W. 2010. Tingkat kerusakanekonomi hama kepik punggung bergaris,Piezodorus hybneri pada kedelai. J. PenelitianPertanian Tanaman Pangan 29(1): 42–49.
No Genotipe Umur masak
(hari) Bobot 100 biji
(g) Hasil
(ton/ha)
1 G 511 H/Anj//Anj///Anj-11-2 77 15,48 2,26 2 G 511 H/Anjasmoro-1-6 75 17,08 2,46 3 G 511 H/Anj//Anj///Anj-7-1 75 17,39 2,12 4 G 511 H/Anjasmoro//Anjasmoro-5-1 77 17,61 2,01 5 G 511 H/Argom//Argom-2-1 77 15,22 2,02 6 G 511 H/Anjasmoro-1-4 77 16,78 2,55 7 G 511 H/Anjasmoro-1-2 76 15,26 2,14 8 G 511 H/Anjasmoro//Anjasmoro-2-8 74 16,87 2,24 9 G 511 H/Arg//Arg///Arg///Arg-12-15 76 16,39 2,60
10 G 511 H/Anj//Anj///Anj-6-3 77 16,83 2,32 11 G 511 H/Arg//Arg///Arg///Arg-19-7 76 17,03 2,06 12 G 511 H/Anjasmoro-1-7 77 16,27 2,05 13 G 511 H/Anj//Anj///Anj////Anjs-6-7 77 16,48 2,03 14 G 511 H/Anjasmoro-1-4-2 77 15,93 2,44 15 Anjasmoro 81 15,67 2,20 16 Grobogan 75 19,73 2,52
Rata-rata 76 16,62 2,30

Bayu et al. Respon Genotipe Kedelai Biji Besar 135
Asadi. 2009. Identifikasi ketahanan sumber daya genetikkedelai terhadap hama pengisap polong. BuletinPlasma Nutfah 15(1): 27–31.
Bae SD, Kim HJ, Yoon YN, Park ST, Choi BR, & JungJK. 2009. Effects of mungbean cultivar,Jangannogdu, on nymphal development, adultlongevity, and oviposition of soybean stink bug.Korean J. Appl. Entomol. 48(3): 311–318.
Bae SD, Kim HJ, & Mainali BP. 2014. Infestation ofRiptortus pedestris (Fabricius) decreases thenutritional quality and germination potential ofsoybean seeds. J. Asia-Pac. Entomol. 17(3):477–481.
Bayu MSYI, Christanto, & Tengkano W. 2011.Komposisi genus dan spesies pengisap polongkedelai pada pertanaman kedelai. Dalam: WidjonoA, Hermanto, Nugrahaeni N, Rahmianna AA,Suharsono, Rozi F, Ginting E, Taufiq A, HarsonoA, Prayogo Y, & Yusnawan E (Eds.) ProsidingSeminar Hasil Penelitian Tanaman AnekaKacang dan Umbi Tahun 2012. pp. 271–278.Tanggal 15 Juli 2012. Balai Penelitian Kacang danUmbi. Malang.
Bayu MSYI & Tengkano W. 2014. Evaluasi ketahanangalur-galur harapan kedelai toleran lahan masamdan kekeringan terhadap kepik coklat. Dalam:Prosiding Seminar Nasional 3 in 1,Hortikultura, Agronomi, dan PemuliaanTanaman. pp. 322–327. Tanggal 21 Agustus 2013.Fakultas Pertanian. Universitas Brawijaya.Malang.
Courtois EA, Paine CE, Blandinieres PA, Stien D, &Bessiere JM, Hovel E, Baraloto C, & Chave J.2009. Diversity of the volatile organic compoundsemitted by 55 species of tropical trees: a surveyin French Guiana. J. Chem. Ecol. 35(11): 1349–1362.
Colazza S & Wajnberg E. 2013. Chemical ecology ofinsect parasitoids: towards a new era. In: WajnbergE & Colazza S (Eds.). Chemical Ecology ofInsect Parasitoids. 1st Edition pp. 1–8. John Wiley& Sons Ltd., Chichester, UK.
Cordeiro EMG, Correa AS, & Guedes RNC. 2014.Insecticide mediated shift in ecological dominancebetween two competing species of grain beetles.PLoS One 9(6): e1009900.
Depieri RA & Panizzi AR. 2011. Duration of feedingand superficial and in-depth damage to soybean
seed by selected species of stink bugs(Heteroptera: Pentatomidae). Neotrop. Entomol.40(2): 197–203.
Furstenberg-Hagg J, Zagrobelny M, & Bak S. 2013.Plant defense against insect herbivores. Int. J.Mol. Sci. 14(5): 10242-10297.
Guedes RNC, Smagghe G, Stark JD, & Desneux N.2016. Pesticide-induced stress in arthropod pestsfor optimized integrated pest managementprograms. Annu. Rev. Entomol. 61: 43-62.
Haddi K, Mendonça LP, Dos Santos MF, Guedes RNC,& Oliveira EE. 2015. Metabolic and behavioralmechanisms of indoxacarb resistance in Sitophiluszeamais (Coleoptera: Curculionidae). J. Econ.Entomol. 108(1): 362–369.
Hendrival, Latifah, & Nisa A. 2013. Efikasi beberapainsektisida nabati untuk mengendalikan hamapengisap polong di pertanaman kedelai. J. Agrista17(18): 18–27.
Krisnawati A, Bayu MSYI, & Adie MM. 2016.Identification of soybean resistance to pod suckingbug (Riptortus linearis) by no-choice test.Biosaintifika 8(3): 407–413.
Leonard BR, Boquet DJ, Padgett B, Davis JA, SchneiderR, Griffin JL, Valverde RA, & Levy Jr RJ. 2011.Soybean green plant malady contributing factorsand mitigation. Louisiana Agric. 54: 32–34.
Lourenção AL, Reco PC, Braga NR, do Valle GE, &Pinheiro JB. 2010. Produtividade de genótipos desoja sob infestação da lagarta-da-soja e depercevejos. Neotropical Entomology. 39(2): 275-281.
Mainali BP & Lim UT. 2012. Annual pattern ofoccurrence of Riptortus pedestris (Hemiptera:Alydidae) and its egg parasitoids Ooencyrtusnezarae Ishii and Gryon japonicum (Ashmead)in Andong, Korea. Crop. Prot. 36: 37–42.
Mainali BP, Kim HJ, Yoon YN, Oh IS, & Bae SD. 2014.Evaluation of different leguminous seeds as foodsources for the bean bug Riptortus pedestris. J.Asia-Pac. Entomol. 17(2): 115–117.
McPherson RM, Buss GR, & Roberts PM. 2007.Assessing stink bug resistance in soybean breedinglines containing genes from germplasm IAC-100.J. Econ. Entomol. 100(4): 1456–1463.
Michereff MFF, Laumann RA, Borges M, Michereff-Filho M, Diniz IR, Neto ALF, & Moreas MCB.

136 J. HPT Tropika Vol. 17 No. 2, 2017: 128 -136
2011. Volatiles mediating a plant-herbivore-naturalenemy interaction in resistant and susceptiblesoybean cultivars. J. Chem. Ecol. 37(3): 273–285.
Morando R, Baldin ELL, Cruz PL, Lourencao AL, &Chiorato AF. 2015. Antixenosis of bean genotypesto Chrysodeixis includens (Lepidoptera:Noctuidae). Pesq. Agropec. Bras. 50(5): 450–458.
Panizzi AR. 2013. History and contemporaryperspectives of the integrated pest managementof soybean in Brazil. Neotrop. Entomol. 42(2):119–127.
Razaq M, Suhail A, Aslam M, Arif MJ, Saleem MA, &Khan HA. 2013. Patterns of insecticides used oncotton before introduction of genetically modifiedcotton in Southern Punjab, Pakistan. Pakistan J.Zool. 45(2): 574–577.
Seo MJ, Kwon HR, Yoon KS, Kang MA, Park MW, JoSH, Shin HS, Kim SH, Kang EJ, Yu YM, & YounYN. 2011. Seasonal occurrence, development, andpreference of Riptortus pedestris on hairy vetch.Korean J. Appl. Entomol. 50(1): 47–53.
Smith CM & Clement SL. 2012. Molecular bases ofplant resistance to arthropods. Annu. Rev.Entomol. 57: 309–328.
Souza BHS, Junior ALB, Janini JC, Silva AGD, &Rodrigues NEL. 2012. Feeding of Spodopteraeridania (Lepidoptera: Noctuidae) on soybeangenotypes. Revista Colombiana de Entomologia38(2): 215–223.
Suharsono & Sulistyowati L. 2012. Expression ofresistance of soybean to the pod sucking bugRiptortus linearis F. (Hemiptera: Coreidae).Agrivita 34(1): 55–59.
Timbo RV, Hermes-Lima M, Silva LP, Mehta A, MoraesMCB, & Paula DP. 2014. Biochemical aspectsof the soybean response to herbivory injury bythe brown stink bug Euschistus heros (Hemiptera:Pentatomidae). PLoS One 9(10): e109735.
Veiga RFA, Rossetto CJ, Razera LF, Galo PB, &Bortoleto M. 1999. Characterizing the morfolologicand agronomic soybean cultivar IAC-100.Agronomic Institute of Campinas. BulletinTechnic 175: 1–25.
War AR, Paulraj MG, Ahmad T, Buhroo AA, HussainB, Ignacimuthu S, & Sharma HC. 2012.Mechanisms of plant defense against insectherbivores. Plant Signal. Behav. 7(10): 1306–1320.

Sudiono et al. Vegetation Diversity 137 J. HPT Tropika. ISSN 1411-7525
Vol. 17, No. 2: 137 – 146, Maret 2017
VEGETATION DIVERSITY AND INTENSITY OF PLANTPESTS AND DISEASES IN TWO POLYCULTURE SYSTEMS
IN TANGGAMUS DISTRICT
Sudiono1,3, Surjono Hadi Sutjahyo2, Nurheni Wijayanto2, Purnama Hidayat2, & Rachman Kurniawan2
1Doctoral Program of Natural Resource and Environment Management, Institut Pertanian BogorJl. R. Pajajaran Baranangsiang, Bogor 16151
2Postgraduate Program, Institut Pertanian BogorJl. Kamper Kampus IPB Dramaga, Bogor 16680
3Plant Protection Department, Faculty of Agriculture, Lampung UniversityJl. Prof. Dr. Sumantri Brodjonegoro No 1, Bandar Lampung 35145
E-mail: [email protected]
ABSTRACT
Vegetation diversity and intensity of plant pests and diseases in two polyculture systems in Tanggamus District. Thevegetable crop management cannot be separated from infestation of plant pest and disease which influences the quality andquantity of crop yield. The pest organism development is influenced by agroecosystem. The objective of this research was toanalyze vegetation diversity and intensity of pest and disease in Tanggamus District. Methods used in this research wereanalysis of vegetation diversity based on Shannon index, percentage of pests damage and diseases incidence. The resultsshowed that the vegetation diversity in polyculture of agriculture typology were 11 plant species with diversity index of 0.64;while in polyculture of agroforestry typology there were 11 plant species with diversity index of 0.74 and both of these indiceswere less than 1 (H’ < 1). The percentage of pests damage in the polyculture of agriculture larger than polyculture ofagroforestry typology ranging from 7.20% to 81.67% and 3.04% to 26.67% respectively. While the incidence of disease inpolyculture of agriculture ranging from 0.65% up to 100% and polyculture of agroforestry typology 0.65% up to 68.00%.
Key words: polyculture of agriculture, polyculture of agroforestry, Shannon Index
ABSTRAK
Keanekaragaman vegetasi dan intensitas hama dan penyakit tanaman sayuran pada dua sistem polikultur diKabupaten Tanggamus. Pengelolaan tanaman sayuran tidak terlepas dari adanya gangguan oleh organisme pengganggutanaman yang berdampak terhadap kualitas dan kuantitas produksi sayuran yang dalam perkembangan organisme tersebutdipengaruhi oleh agroekosistem. Tujuan penelitian ini adalah menganalisis keanekaragaman vegetasi tumbuhan dan besarnyaserangan hama dan penyakit tanaman pada tanaman sayuran di Kabupaten Tanggamus. Metode penelitian yang digunakanyaitu analisis keanekaragaman vegetasi berdasarkan indek Shannon, persentase serangan hama dan kejadian penyakit. Hasilpenelitian menunjukkan keanekaragam vegetasi di tipologi polikultur pertanian tercatat 11 species tumbuhan dengan indekkeanekaragaman vegetasi sebesar 0,64; sedangkan pada tipologi polikultur agroforestri tercatat 11 species tumbuhan denganindek keanekaragaman sebesar sebesar 0,74 yang keduanya lebih kecil dari 1 (H’ < 1). Intensitas serangan hama dan kejadianpenyakit tanaman pada tipologi polikultur pertanian lebih besar dibandingkan polikultur agroforestri dengan kisaran hamasebesar 7,20% sampai 81,67% berbanding 3,04% sampai 26,67% dan penyakit antara 0,65% sampai 100% berbanding 0,65%sampai 68,00%.
Kata kunci: indeks Shanon, polikultur pertanian, polikultur agroforestri
INTRODUCTION
An agricultural typology or landscape hasdifferent aesthetical qualities from one to another. Thisdepends on diversity of plant types to cultivate.Polyculture of agriculture and agroforestry are examples
of differences of agricultural diversity types. These twotypes of culture have roles not only at production aspects,but also in other environment aspects including diversityof pests and plant pathogens. Diversity, which is allcompositions of types of plants, animals, andmicroorganisms interacting in a particular ecosystem is

138 J. HPT Tropika Vol. 17, No. 2, 2017: 137 - 146
very influential in determining agriculture productivitylevel. Diversity in an agricultural landscape attractsattentions from many experts, because it contributessignificantly to the agriculture productivity, foodsustainability, financial profit, and conservation of nature(Liu et al., 2013).
Plant diversity in an agroecosystem can reducethe effects of pest and diseases through some waysboth individually and in combinations, including someeffects of pest oppression in visual and pest smellingways, disorders in pest life cycles, reducing inoculumsbecause there is no host plant, antagonist mechanism,the plant physiology resistance because sufficiency ofplant nutrients, natural predator conservation, andagricultural landscape effects such as physical barriersand micro climate changes (Ratnadass et al., 2012).Global practices of diversity are familiar to agriculturepeople, because the agriculture activity covering 25-30%areas in the world is an important activity influencingdiversity.
There is a correlation between agroecosystemdiversity and pest disease incidence with indicators suchas level of pests/disease incidence, low productivity, andpesticide residue. Some research results showed thatagrochemical treatments (especially pesticide anfertilizer) had caused problem social and environmentalchanges (Altieri et al., 1984; Altieri, 1999; Scherret al.,2008). Agricultural and forestry environmentmanagements are keys for conservation of diversitywhich significantly improve richness and abundancesof species, while tree able to reduce able to improvepest, pathogen incidence, and also to provide ecologyservices in strengthening beneficial insects (Batary etal., 2011; Tomback et al., 2016; Aluja et al., 2014).
Agriculture systems in developing countries withrelatively small sizes of land provide opportunities toreduce effects of pest and disease through practices ofincrease diversity. This method is very important inconducting sustainable agriculture system (Simon et al.,2010, Muniappan & Heinrichs, 2014). The objective ofpest management is to contribute agriculturesustainability with some different aspects such as foodsustainability, balanced relationship between human andecosystem, and ecosystem conservation. Theagroecosystem diversification efforts often reducenegative effects of modern agriculture or agriculturalindustry (Timprasert et al., 2014; Gurr et al., 2013;Savary et al. , 2012). Agriculture landscape isrepresentation of habitat including plants, forests, wetlands, and grass land. The diversity of vegetation canserve the dynamics of pest and natural enemy (Philpott,2013).
The objective of this research was to analyze thevegetation diversity and the damage of pests and theincidence of disease vegetable crop in TanggamusDistrict Lampung Province.
MATERIALS AND METHODS
Study Site. This research was conducted in Gisting andSumberejo Subdistrict, Tanggamus District LampungProvince and Laboratory of Plant Pests and Disease ofFaculty of Agriculture, Lampung University, from Marchto November 2015.
Research Procedure. Data in this research werevegetation diversity, damage of pests and the disease ofincidence in some vegetable crops in polyculture ofagriculture and polyculture of agroforestry typologies.
This research used quadrant sampling technique.The research locations were taken purposively torepresent typology of the regions i.e. the polyculture withcrop (polyculture of agriculture) and polyculture withforest tree (polyculture of agroforestry). From eachtypology, five were selected where plot 400 m2. Fromeach plot the following variable were observed 1) plantspecies and numbers, 2) vegetation diversity, 3) intensityof pests damage, and 4) incidence of disease. Thevegetation diversity was determined using Shannon’sindex (Shannon and Wienner in Ludwig & Reynolds,1988) as follows:
With:H’ = Shannon’s diversity index of vegetatiton in sampled plotni = Number of species in plotn = Total number of plant in plots = Total number of plant species observed in the plot
The value of diversity index is defined as folow(1) H’ > 3, indicates high diversity high; (2) H’ 1 < H’ <3 indicates moderate diversity, and (3) H’ < 1 indicateslow diversity (Mason & McDonald, 1986).
The intensity of plant pest and incidence ofdiseasein each sampaled plot was determined with thefollowing equation:
)}nn(ln )
nn{( H' i
s
1 i
i
100%Nn
P

Sudiono et al. Vegetation Diversity 139
P = Intensity of pest damage or disease incidencen = Numbers of plants damaged by pests or disease in
plotN = Total numbers of plants observed in plot
RESULTS AND DISCUSSION
The research was conducted in Gisting AtasVillage, Gisting Bawah Village of Gisting Subdistrict, andWonoharjo Village, Sumber Muyo Village, SimpangKanan Village of Sumberejo Subdistrict in TanggamusDistrict, Lampung Province. The site can be reached in± 2-3 hours by car from the Province capital BandarLampung. This site at 600-1000 m above the sea level(asl) with air temperature ranges of 18-28 oC.Geographically Tanggamus District is located at 104°18’-105°12’ east longitude and 5°05’-5°56’ south latitude,with flat to wavy topographies. The most common
vegetation types in these regions area annual plants suchas rice and horticultures and perennial plants such ascacao and coffee. There is a conservational forest inthe Tanggamus mountain slopes and this is integrated toConservational Forest Management Unit (or KPHL) ofNorth Kota Agung area, Lampung Province. The forestsite is located in the vicinity of local state crop area andthe site is managed jointly by the local community andlocal goverment ini form community forest or publicforest (or Hkm). This public forest has poor road acces.It only footpaths where farmers can walk along or drivetheir motorcyle. Meanwhile the agricultural fields in thesitewith relatively flat topography were owned andcultivated by local farmers. Infrastructures like roads,drainages, irrigation, and electricity are available in thisagricultural part of the site. The summary of researchsite presented in Table 1.
Table 1. Summary of information abaout research site
Plot Villages Location Coordinate (GPS) Elevation
(asl,M) Kinds of plants South latitude East longitude
1 Gisting Bawah, Gisting
05°.25’.30,4" 104°.43’.56,5" 512 chili, cabbage, mung bean, Green mustard, eggplant, tomato
2 Simpang Kanan, Sumberejo
05°.23'.23,3" 104°.43'.09,5" 536 chili, mung bean, cabbage, rice
3 Sumber Mulyo, Sumberejo
05°.22'.21,1" 104°.43'.10,5" 490 chili, mung bean, eggplant, rice, jabon, papaya, banana, mindi
4 Wonoharjo, Sumberejo
05°.22'.21,1" 105°.23'.16,4" 496 tomato, cabbage, mung bean, yardlong bean, chili, eggplant, rice, coconut, mindi
5 Gisting Atas, Gisting
05°.26'.39,7" 104°.43'.42,4" 568 Tomato, clove, banana, coconut, mindi, nutmeg, papaya
6 Gisting Permai, Gisting
05°.27'.12,9" 104°.42'.56,7" 600 eggplant, jabon, white teak, nutmeg, papaya, cacao, rubber, mindi, coffee
7 Gisting Atas, Gisting
05°.26'.05,9" 104°.42'.22,3" 767 chili, cabbage, tomato, green mustard, nutmeg, banana, mahogany, coffee, bitter bean, teak, mindi, avocado
8 Gisting Atas, Gisting
05°.26'.06,7" 104°.42'.10,5" 806 Green mustard, onion leafs, cabbage, chili, rubber, banana, bamboo, cacao, coffee, mindi, papaya
9 Gisting Atas, Gisting
05°.26'.00,1" 104°.42'.09,1" 842 chili, cabbage, green mustard, banana, mahogany, mindi, lamtoro, medang, pule
10 Gisting Atas, Gisting
05°.25'.54,8" 104.42'.03,2" 900 cabbage, eggplant, onion leaf, banana, pea, mahogany, pule, coffee, sugar palm, avocado, papaya
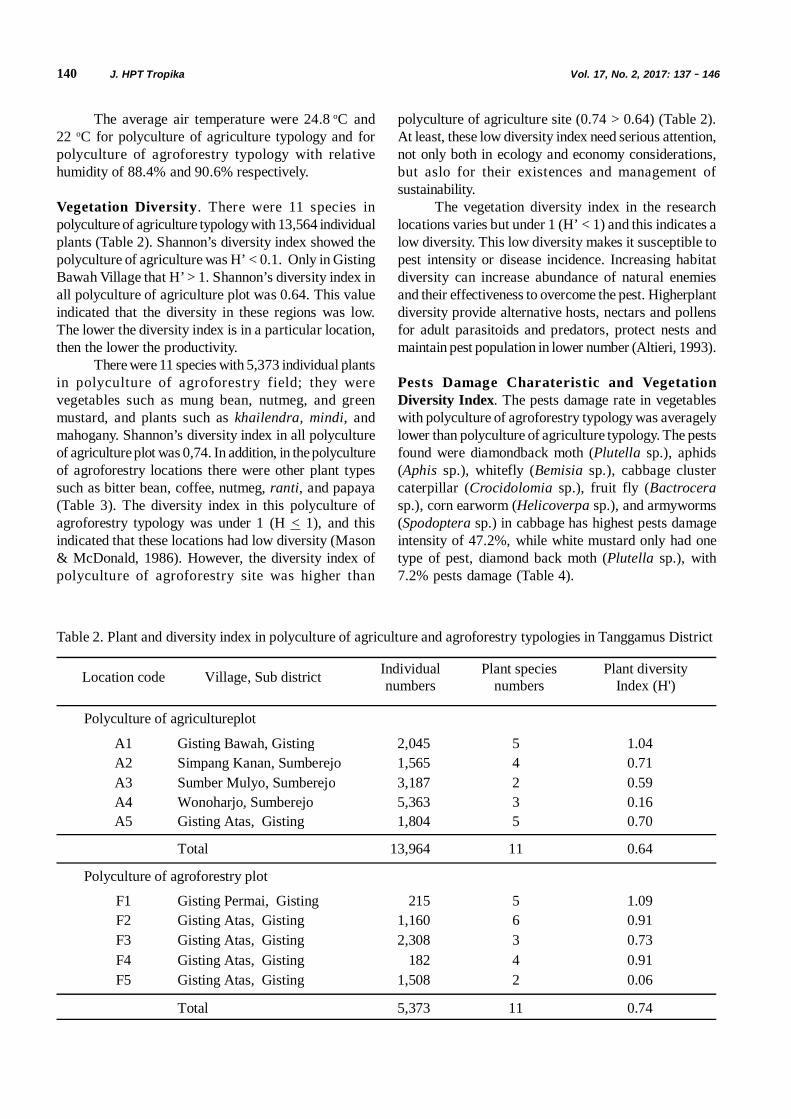
140 J. HPT Tropika Vol. 17, No. 2, 2017: 137 - 146
The average air temperature were 24.8 oC and22 oC for polyculture of agriculture typology and forpolyculture of agroforestry typology with relativehumidity of 88.4% and 90.6% respectively.
Vegetation Diversity. There were 11 species inpolyculture of agriculture typology with 13,564 individualplants (Table 2). Shannon’s diversity index showed thepolyculture of agriculture was H’ < 0.1. Only in GistingBawah Village that H’ > 1. Shannon’s diversity index inall polyculture of agriculture plot was 0.64. This valueindicated that the diversity in these regions was low.The lower the diversity index is in a particular location,then the lower the productivity.
There were 11 species with 5,373 individual plantsin polyculture of agroforestry field; they werevegetables such as mung bean, nutmeg, and greenmustard, and plants such as khailendra, mindi, andmahogany. Shannon’s diversity index in all polycultureof agriculture plot was 0,74. In addition, in the polycultureof agroforestry locations there were other plant typessuch as bitter bean, coffee, nutmeg, ranti, and papaya(Table 3). The diversity index in this polyculture ofagroforestry typology was under 1 (H < 1), and thisindicated that these locations had low diversity (Mason& McDonald, 1986). However, the diversity index ofpolyculture of agroforestry site was higher than
polyculture of agriculture site (0.74 > 0.64) (Table 2).At least, these low diversity index need serious attention,not only both in ecology and economy considerations,but aslo for their existences and management ofsustainability.
The vegetation diversity index in the researchlocations varies but under 1 (H’ < 1) and this indicates alow diversity. This low diversity makes it susceptible topest intensity or disease incidence. Increasing habitatdiversity can increase abundance of natural enemiesand their effectiveness to overcome the pest. Higherplantdiversity provide alternative hosts, nectars and pollensfor adult parasitoids and predators, protect nests andmaintain pest population in lower number (Altieri, 1993).
Pests Damage Charateristic and VegetationDiversity Index. The pests damage rate in vegetableswith polyculture of agroforestry typology was averagelylower than polyculture of agriculture typology. The pestsfound were diamondback moth (Plutella sp.), aphids(Aphis sp.), whitefly (Bemisia sp.), cabbage clustercaterpillar (Crocidolomia sp.), fruit fly (Bactrocerasp.), corn earworm (Helicoverpa sp.), and armyworms(Spodoptera sp.) in cabbage has highest pests damageintensity of 47.2%, while white mustard only had onetype of pest, diamond back moth (Plutella sp.), with7.2% pests damage (Table 4).
Location code Village, Sub district Individual numbers
Plant species numbers
Plant diversity Index (H')
Polyculture of agricultureplot
A1 Gisting Bawah, Gisting 2,045 5 1.04 A2 Simpang Kanan, Sumberejo 1,565 4 0.71 A3 Sumber Mulyo, Sumberejo 3,187 2 0.59 A4 Wonoharjo, Sumberejo 5,363 3 0.16 A5 Gisting Atas, Gisting 1,804 5 0.70
Total 13,964 11 0.64
Polyculture of agroforestry plot F1 Gisting Permai, Gisting 215 5 1.09
F2 Gisting Atas, Gisting 1,160 6 0.91 F3 Gisting Atas, Gisting 2,308 3 0.73 F4 Gisting Atas, Gisting 182 4 0.91 F5 Gisting Atas, Gisting 1,508 2 0.06
Total 5,373 11 0.74
Table 2. Plant and diversity index in polyculture of agriculture and agroforestry typologies in Tanggamus District

Sudiono et al. Vegetation Diversity 141Ta
ble
3.Ve
geta
tion
in v
ario
us p
olyc
ultu
re o
f agr
icul
ture
and
agro
fore
stry
typo
logi
es in
Tan
ggam
us D
istric
t

142 J. HPT Tropika Vol. 17, No. 2, 2017: 137 - 146
Pests damage intensity on polyculture ofagroforestry typology was lower than that polycultureof agriculture typology. Chili and tomato had highestpests damage intensity (13.6% and 11.8%), while greenmustard and currant tomato (rampai) had no pestsdamage. A possible reason for the lower pests damageon polyculture of agroforestry was that agroecosystem
might be stable so that it was able to facilitate betteractivities of natural enemies which in enable them controland maintain pest population at low number.
The differences of seasons and populationdynamics influence insects in different habitats whichchange over time and according to development stages(Teodore et al., 2008). Natural vegetation is able to
Table 4.Crop pest damage in polyculture of agriculture and agroforestry typologies in Tanggamus District
Location code Type of plant Kinds of pests Species Pests damage (%) Polyculture of agriculture
A1 White mustard Diamond back moth Plutella sp. 7.2
Mung bean - -
Chili - -
Currant tomato - -
A2 Yardlong bean Aphids Aphis sp. 81.7
Pod borer Lepidoptera sp. 10.6
Chili Whitefly Bemisia sp. 32.9
A3 Eggplant - - -
Mung bean Diamond back moth Plutella sp. 67.0
Leaf caterpillar Crocidolomia sp. 18.7 A4 Cabbage Diamond back moth Plutella sp 63.0
Leaf caterpillar Crocidolomia sp. 63.0
Green mustard - -
Chili Aphids Aphis sp. 31.4 Whitefly Bemisia sp. 31.4 A5 Tomato Fruit flies Bactrocera sp. 12.6
Corn earwarm Helicoverpa sp. 10.3
Whitefly Bemisia sp. 12.6
Leek - - -
Polyculture of agaroforestry F1 Mung bean Diamond back moth Plutella sp. 8.8 F2 Currant tomato - - - Green mustard - - - F3 Tomato Whitefly Bemisia sp. 9.0
Corn earwarm Helicoverpa sp. 5.0
Fruit flies Bactrocera sp. 20.0 F4 Currant tomato Armyworm Spodoptera sp. 6.0 Corn earwarm Helicoverpa sp. 3.0 F5 Chili Trips Trips sp. 3.0
Armyworm Spodoptera sp. 11.0
Whitefly Bemisia sp. 27.0

Sudiono et al. Vegetation Diversity 143
Table 5. Vegetation diversity index (H’) and pests damage percentage for vegetable crop in Tanggamus District
Location code Village, Sub district Vegetation diversity index (H')
Ranges of pest damage (%)
Polyculture of agriculture A1 Gisting Bawah, Gisting 1.26 0.00-7.20 A2 Simpang Kanan, Sumberejo 0.71 10.64-81.67 A3 Sumber Mulyo, Sumberejo 0.59 18.74-66.99 A4 Wonoharjo, Sumberejo 0.83 31.36-62.99 A5 Gisting Atas, Gisting 0.70 10.32-12.59
Polyculture of agroforestry F1 Gisting Permai, Gisting 1.09 0.00-8.83 F2 Gisting Atas, Gisting 0.91 0.00-0.00 F3 Gisting Atas, Gisting 0.73 5.08-19.92 F4 Gisting Atas, Gisting 1.01 2.50-6.41 F5 Gisting Atas, Gisting 0.75 3.04-26.67
promote natural enemy diversity with varying effects;the same agroecosystem can explain some pests whichare more influential than others in a more complexlandscape (Henri et al., 2015).
The agroforestry landscape perspective enablesknowledge transfer between farmers on agronomy andecology in participative approach to help driving amanagement which balances economy and ecologyneeds (Tscharntke et al. , 2011). Polyculture ofagriculture reduces ecological sustainability of land usesystem, while environment change and extreme climaterequire higher quality responses than ever. Adaptationstrategies to environment changes such as cultivatingshady trees and field conversion type are commonpractices to improve sustainability against environmentchanges. Polyculture of agriculture pattern in a wideoverlay is a good for plant growth and development aswell as for pest migration from one place to another. Auniformed cultivar in a wide field creates the samesituations with effects of interactions of pest, pathogen,and plant growth (Altieri & Nichollas, 2004).
The vegetation diversity index in polyculture ofagriculture system ranged 0.59-1.26; while in polycultureof agroforestry system the vegetatiaon diversity indexranged 0.73-1.09 (Table 5). The diversity index value islower than criteria to use; under 1 (H’< 1). Lowerdiversity index value means lower productivity asindications of severe ecology pressure and unstableecosystem. Mason & McDonald (1986) suggests thatin diversity index value is lower than 1, it means lowdiversity. If it is around 1-3, it means moderate diversity,and more than 3 means high diversity. The low diversityindex in the locations are caused by the facts that thefield is continually planted, with low supply of soil
nutrients, excessive sun rays, and few water supply,which make trees are difficult to grow in these areas.
Charateristic of Disease Incidence and VegetationDiversity Index. The disease incidence rate inpolyculture of agriculture typology varies between 1%to 100%, pathogen infection of Colletotrichum sp. andAlternaria sp. caused 100% disease incidence especiallyin mung bean. The infections are was low in whitemustard, cabbage, eggplant, chili, and their pathogensare Erwinia sp., Plasmidiophora sp., and Cersosporasp. The only vegetable crop free from disease is leek,and the highest rate of disease occurred in tomato.
The disease incidence in vegetable crop inpolyculture of agroforestry typology is lower than inpolyculture of agriculture typology range 0.65% to 100%versus 0.67% to 68.00%. The highest disease incidenceis in tomato and chili, while the lowest is in currant tomatovegetable (Table 6).
Each typology has different diversity index anddisease incidence rate and polyculture of agroforestryhave higher diversity index than polyculture ofagriculture (Table 7). The polyculture of agriculturetypology is potentially more susceptible to disease thanthe polyculture of agroforestry. The agroforestry patterninfluences on pest and disease are not only dependingon the types of plants, but also depending on otherfactors including kinds of pests, pest preferences, andmicro climate (Tomlinson et al., 2015; Pumarino et al.,2015). Structure and landscape heterogeneity tends toinfluence disease dynamics and distributions and theyfunction as inhibitors to limit pathogen distribution(Plantegenest & Fabre, 2007).

144 J. HPT Tropika Vol. 17, No. 2, 2017: 137 - 146
Table 6. Types diseases and incidence in some vegetable crops in polyculture of agriculture and agroforestrytypologies in Tanggamus District
Location code Kinds of plant Disease Pathogen
Disease incidence
(%) Polyculture of agriculture
A1 White mustard Leaf blight Erwinia sp. 6.24 Clubroot Plasmodiophora sp. 0.65 Leaf spots Xanthomonas sp. 1.91 Mung bean Anthracnose Colletotrichum sp. 100.00 Black spot leaf Alternaria sp. 100.00 Chili Leaf curl Curly virus 5.33 Yellow leaf Gemini virus 1.34
Currant tomato - - - A2 Yardlong bean Black spot leaf Uromyces sp. 10.90
Cowpea witches-broom Cowpea witches-broom virus 2.05 Chili Anthracnose Colletotrichum sp. 100.00
Leaf curl Curly virus 100.00 A3 Eggplant Leaf spot Cercospora sp. 3.70
Water molds Phytophthora sp. 1.00 Mung bean Anthracnose Colletotrichum sp. 20.00
Black spot leaf Uromyces sp. 80.00 A4 Cabbage Leaf spot Cercospora sp. 1.26
Clubroot Plasmodiophora sp. 1.51 Green mustard Clubroot Plasmodiophora sp. 0.50 Leaf blight Erwinia sp. 2.83 Chili Leaf curl Curly virus 33.90
Yellow leaf Gemini virus 16.95 A5 Tomato Anthracnose Colletotrichum sp. 78.00
Leaf spots Cercospora sp. 68.00 Soft rot Phytophthora sp. 100.00
Leek - Erwinia sp. - Polyculture of agroforestry
F1 Mung bean Black spot leaf Uromyces sp. 5.00 F2 Currant tomato - - - Green mustard Leaf blight Erwinia sp. 2.34
F3 Tomato Leaf spot Cercospora sp. 16.02 Black spot leaf Uromyces sp. 10.81
Anthracnose Colletotrichum sp. 40.00 F4 Currant tomato Leaf spot Cercospora sp. 14.06 Anthracnose Colletotrichum sp. 4.69
F5 Chili Anthracnose Colletotrichum sp. 68.00 Yellow leaf Gemini virus 0.67
Leaf curl Curly virus 2.17

Sudiono et al. Vegetation Diversity 145
Table 7. Vegetation diversity index (H’) and disease incidence in vegetable crop in Tanggamus District
Location code Village, sub district Vegetation diversity index (H')
Ranges of disease incidence(%)
Polyculture of agriculture A1 Gisting Bawah, Gisting 1.26 0.65-100 A2 Simpang Kanan, Sumberejo 0.71 2.05-100 A3 Sumber Mulyo, Sumberejo 0.59 1.00-80.00 A4 Wonoharjo, Sumberejo 0.83 0.50-33.90 A5 Gisting Atas, Gisting 0.70 68.00-100
Polyculture of agroforestry F1 Gisting Permai, Gisting 1.09 0.65-8.83 F2 Gisting Atas, Gisting 0.91 0.00-0.00 F3 Gisting Atas, Gisting 0.73 5.08-19.92 F4 Gisting Atas, Gisting 1.01 2.50-6.41 F5 Gisting Atas, Gisting 0.75 3.04-26.67
CONCLUSION
The conclusions of this research are that thevegetation diversity in polyculture of agriculture typologywere 11 plant species with diversity index of 0.64, whilein polyculture of agroforestry typology there were 11plant species with diversity index of 0.74, and both ofthese indices were less than 1 (H’ < 1). The percentageof pests damage in the polyculture of agriculture largerthan polyculture of agroforestry typology the range of7.20 to 81.67% and 3.04 to 26.67% and the incidencedisease in polyculture of agriculture ranged from 0.65up to 100% and polyculture of agroforestry typology0.65 up to 68.00%.
ACKNOWLEDGMENT
We thank the majors of Plant Protection, Facultyof Agriculture, Lampung University for supporting thisresearch and for providing facilities to do laboratoriumanalysis.
REFERENCES
Altieri MA, Letourneau DK, & Risch SJ. 1984.Vegetation diversity and insect pest outbreaks.Critical Reviews in Plant Sciences. 2(2): 131–169.
Altieri MA & Nicholls. 2004. Agroecology and theSearch for a Truly Sustainable Agriculture.United Nations Environment Programme.Mexico.
Altieri MA. 1993. Ethnoscience and biodiversity: keyelements in the design of sustainable pestmanagement systems for small farmers indeveloping countries. Agr. Ecosyst. Environ. 46:257–272.
Altieri MA. 1999. The ecological role of biodiversity inagroecosistems. Agr. Ecosyst. Environ. 74(1):19–31.
Aluja M, Sivinski J, Driesche RV, Dadda AA, & GuillenL. 2014. Pest management through tropical treeconservation. Biodivers. Conserv. 23(4): 831–853.
Batary P, Baldi A, Kleijn D, & Tscharntke T. 2011.Landscape-moderated biodiversity effects ofagrienvironmental management: a meta-analysis.Proc. Biol. Sci. 278(1713): 1894–1902.
Henri DC, Jones O, Tsiattalos A, Thebault E, SeymourCL, & Van Voen FJF. 2015. Natural vegetationbenefits synergistic control of the three main insectand pathogen pests of a fruit crop in SouthernAfrica. J. Appl. Ecol. 52(4): 1092–1101.
Gurr GM, Wratten SD, & Luna JM. 2003. Multi-functionagricultural biodiversity: pest management andother benefits. Basic Appl. Ecol. 4: 107–116.
Liu Y, Duan M, & Yu Z. 2013. Agricultural landscapesand biodiversity in China. Agri. Ecosyst. Environ.166: 46–54.

146 J. HPT Tropika Vol. 17, No. 2, 2017: 137 - 146
Ludwig JA & JF Reynolds JF. 1988. StatisticalEcology: an Primer Methods and Computing.John Wiley and Sons Inc. New York.
Mason CF & McDonald SM. 1986. Otters: Ecologyand Conservation. Cambridge University Press.Cambridge.
Muniappan R, Director P, & Heinrichs EA. 2014.Biodiversity and integrated pest management:working together for a sustainable future. CropProt. 61: 102–110.
Philpott SM. 2013. Biodiversity and pest control services.Reference Module in Life SciencesEncyclopedia of Biodiversity 1: 373–385.
Plantegenest M, May CL, & Fabre F. 2007. Landscapeepidemiology of plant diseases. J. R. Soc.Interface. 4(16): 963–972.
Pumarinoa L, Sileshib GW, Gripenbergc S, KaartinenaR, Barriosb E, Muchaned MN, Midegae C, &Jonssona M. 2015. Effects of agroforestry on pest,disease and weed control: a meta-analysis. BasicAppl. Ecol. 16(7): 573–582.
Ratnadass A, Fernandes P, Avelino J, & Habib R. 2012.Plant species diversity for sustainablemanagement of crop pests and diseases inagroecosystems: a review. Agron. Sustain. Dev.32(1): 273–303.
Savary S, Horgan F, Willocquet L, & Heong K.L. 2012.A review of principles for sustainable pestmanagement in rice. Crop Prot. 32: 54–63.
Scherr SJ & McNeely JA. 2008. Biodiversityconservation and agricultural sustainability:towards a new paradigm of ‘ecoagriculture’landscapes. Philos Trans. R. Soc. Lond. B. Biol.Sci. 363: 477–494.
Simon S, Bouvier JC, Debras JF, &Sauphanor B. 2010.Diversity and pest management in orchardsystems. A review. Agron. Sustain. Dev. 30: 139–152.
Teodoro A, Klein AM, & Tscharntke T. 2008.Environmentally mediated coffee pest densitiesin relation to agroforestry management, usinghierarchical partitioning analyses. Agr. Ecosyst.Environ. 125(1–4): 120–126.
Timprasert S, Datta A, & Ranamukhaarachchi SL. 2014.Factors determining adoption of integrated pestmanagement by vegetable growers in NakhonRatchasima Province, Thailand. Crop Prot. 62:32–39.
Tomback DF, Resler LM, Keane RE, Pansing ER,Andrade AJ, & Wagner AC. 2016. Communitystructure, biodiversity, and ecosystem services intreeline whitebark pine communities: potentialimpacts from a non-native pathogen. Forests 7:1–22.
Tomlinson I, Potter C, & Bayliss H. 2015. Managingtree pests and diseases in urban settings: the caseof oak processionary moth in London, 2006-2012.Urban For. Urban Gree.14(2): 286–292.
Tscharntke T, Clough Y, Bhagwat SA, Buchori D, FaustH, Hertel D, Holscker D, Juhrbandt J, KesslerM, Perfecto I, Scherber C, Schoth G, VeldkampE, & Wanger TC. 2011. Multifunctional shade-tree management in tropical agroforestrylandscapes-a review. Appl. Ecol. 48(3): 619–629.

156 J. HPT Tropika Vol. 17 No. 2, 2017: 156 - 161 J. HPT Tropika. ISSN 1411-7525
Vol. 17, No. 1: 156 – 161, September 2017
INTRODUKSI FORMULA PUPUK HAYATI BERBAHAN AKTIFPSEUDOMONAD FLUORESCENT ISOLAT PF-122 UNTUK
MENINGKATKAN PERTUMBUHAN DAN PRODUKSITANAMAN CABAI DI LAPANG
Yenny Wuryandari, Sri Wiyatiningsih, & Maroeto
Jurusan Agroteknologi, Fakultas Pertanian, UPN “Veteran” Jawa TimurJl. Raya Rungkut Madya, Gunung Anyar, Surabaya, Jawa Timur 60294
E-mail: [email protected]
ABSTRACT
Introduction of biofertilizer containing pseudomonad fluorescent isolate Pf-122 as active ingredient for improving growthand production of chili plant in the field.Some biological control agents required an efficient formulation required to beapply on large scale in the field. The purpose of this research is to review some biofertilizer formulations formof active ingredients pseudomonad fluorescent Pf-122 which can increase growth and production chilli in the field. Theresearch using Randomized Complete Block Design (RCBD) which consists three kinds of formulation. The formulations arepowder, granules and pellets. Chilli without formulations as a control. The best results was growth and production of chilli canbe showed from plant height, branch number, productive branch number, fruit number, and fruit weight which was givenpowder formulation. The results of fruit number, the powder formulation which is given can produce double chilli yield thanyield from control plant. Moreover, weight fruit from chilli which is given powder formulation can produce fourfold yields.Growth and production chilli which is given others formulations such as granules and pellets were not significantly differentresults. The overall result of this research, the best formulation to increase growth and production of chilli in the field wasformulations contain active biopesticide pseudomonad fluorescent Pf-122 in powder.
Key words: granules, pellets, powder, pseudomonads fluorescent
ABSTRAK
Introduksi formula pupuk hayati berbahan aktif pseudomonad fluorescent isolat Pf-122 untuk meningkatkan pertumbuhandan produksi tanaman cabai di lapang. Untuk beberapa agens pengendali biologi tanaman, suatu formulasi yang efisiendiperlukan untuk aplikasi dalam skala besar di lapang. Tujuan penelitian untuk mengkaji beberapa formulasi pupuk hayatiberbahan aktif pseudomonad fluoresent isolat Pf-122 dalam memacu pertumbuhan dan produksi cabai pada skala lapang.Penelitian menggunakan Rancangan Acak Kelompok (RAK) yang terdiri dari 3 macam formulasi yaitu serbuk, granul dan pelet.Sebagai kontrol tanaman cabai tanpa pemberian formula. Hasil penelitian menunjukkan bahwa pertumbuhan dan produksicabai baik tinggi tanaman, jumlah cabang, jumlah cabang produktif, jumlah buah dan berat buah pada tanaman yangdiperlakukan dengan formulasi serbuk paling baik dibandingkan pada kontrol dan formulasi yang lain. Untuk hasil jumlahbuah, pemberian formulasi serbuk dapat mencapai dua kali hasil pada kontrol bahkan untuk berat buah dapat mencapai lebihdari empat kali lebih tinggi dari pada kontrol. Pada peubah pertumbuhan dan produksi tanaman cabai pemberian formulasigranul dan pelet tidak berbeda nyata hasilnya. Dengan demikian introduksi formulasi serbuk berbahan aktif pseudomonadfluorescent isolat Pf-122 adalah yang paling baik dalam memacu pertumbuhan dan produksi cabai di lapang.
Kata kunci: granul, pelet, pseudomonad fluorescent, serbuk
PENDAHULUAN
Salah satu jenis sayuran buah yang memilikipotensi untuk dikembangkan karena cukup pentingperanannya baik untuk memenuhi kebutuhan konsumsinasional maupun komoditas ekspor adalah cabai merah(Capsicum annuum L .). Produktivitas cabai diIndonesia saat ini masih tergolong rendah. Faktor-faktor
yang menyebabkan rendahnya produksi antara lainpenggunaan benih yang kurang bermutu, teknikbudidaya yang belum efisien dan penanaman kultivarcabai yang tidak tahan terhadap hama serta penyakit(Soelaiman & Ernawati, 2013). Untuk mengatasimasalah penyakit cabai ini, pada umumnya petanimelakukan pengendalian secara konvensional, yaitudengan menggunakan pestisida secara intensif.

Wuryandari et al. Introduksi Formula Pupuk Hayati 157
Penggunaan pestisida ini bahkan mencapai 51% daribiaya produksi. Dari jumlah tersebut, 17,6% digunakanuntuk mengatasi penyakit tanaman, dan sisanya untukpenggunaan insektisida (Anonim, 2009). Upaya lainuntuk mengatasi penyakit tanaman yang tidak kalahpenting adalah meningkatkan ketahanan danpertumbuhan cabai.
Rhizobacteria atau bakteri rizosfer banyak yangmempunyai prospek dan potensi pengimbas pertumbuhantanaman. Bakteri rizosfer ada yang bersifat saprofitikdan bersifat parasit. Beberapa bakteri saprofitiksebagai agens hayati pengendali penyakit layu antaralain adalah bakteri Pseudomonas fluorescens padatomat, kentang, dan terung, serta Pseudomonas sp. padacabai (Saddler, 2005). Pengendalian hayati patogentanaman dengan rhizobakteria dapat didasarkan padaproduksi metabolit bakteri seperti siderofor, antibiotik,dan hydrogen cyanida. Rhizobakteria juga mampumemacu pertumbuhan tanaman dengan memproduksizat pengatur tumbuh seperti turunan auxin (Pinton etal., 2001). Sebagian besar rhizobakteria sepertiPseudomonas, Bacillus, Azotobacter memproduksisenyawa pemacu pertumbuhan seperti Indolacetic acid(IAA), gibberellin, substansi seperti cytokinin(Brimecombe et al., 2001). Bakteri mampu memacuketahanan sistemik dengan memproduksi fitohormon,melarutkan fosfat anorganik, meningkatkan pengikatanFe dengan siderofor (Podile & Kishore, 2007; Haas &Defago, 2005).
Mekanisme lain dari rhizobacteria atau bakteririzosfer dalam meningkatkan pertumbuhan tanamanadalah sebagai penyedia fosfat bagi tanaman. Olehkarena itu salah satu cara untuk memperbaiki defisiensifosfat pada tanaman adalah dengan inokulasi tanahdengan mikroorganisme pelarut fosfat. Salah satubakteri yang merupakan pelarut potensial dari fosfatadalah Pseudomonas dan Bacillus (Beattie, 2007).
Hasil penelitian Wuryandari et al. (2013)menunjukkan bahwa isolat pseudomonad fluoresent Pf-122 mampu meningkatkan pertumbuhan dan produksitanaman cabai pada skala lapang. Peningkatan jumlahbuah maupun berat buah cabai sangat tinggi sampai lebihdari 4 kali dibandingkan kontrol atau tanpa pemberianpseudomonad fluoresent Pf-122. Aplikasi pseudomonadfluoresent Pf-122 pada tanaman cabai adalah denganpencelupan bibit cabai.
Untuk beberapa agens pengendali biologitanaman, suatu formulasi yang efisien diperlukan untukaplikasi dalam skala besar di lapangan. Pabrik mini yangdimiliki UPN Veteran Jawa Timur telah memproduksisecara rutin beberapa pupuk organik. Produksi pupukbelum optimal sehingga perlu dilakukan upaya untuk
mengembangkan dan meningkatkan produksinyasehingga kualitas dan kuantitasnya semakin meningkat.Untuk itu perlu memberi nilai tambah pupuk organik yaitudengan menambah atau menyisipkan bahan aktifrhizobakteria pseudomonad fluoresent isolat Pf-122.Pupuk organik yang telah ditambah rhizobakteriapseudomonad fluoresent isolat Pf-122 diharapkan selainmampu bertahan dari penyakit juga lebih menginduksidan meningkatkan produksi tanaman. Oleh karena itudilakukan pengembangan produk pupuk menjadiformula dengan bahan aktif pseudomonad fluoresentisolat Pf-122 yang mampu memacu pertumbuhan danproduksi tanaman cabai. Tujuan penelitian ini adalahuntuk mendapatkan formulasi pupuk hayati denganbahan aktif pseudomonad fluoresent isolat Pf-122 yangpaling baik dalam meningkatkan pertumbuhan danproduksi cabai.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilakukan di lahanpertanaman cabai di Desa Lebo, Kecamatan Sukodono,kabupaten Sidoarjo, Jawa Timur. Penelitian dilakukanpada bulan Mei sampai Juli 2015.
Rancangan Penelitian. Penelitian menggunakanRancangan Acak Kelompok (RAK) yang terdiri dari 3perlakuan yaitu serbuk, pelet, dan granul. Bentukformula tersebut dibuat dengan bahan aktif bakteripseudomonad fluorescent isolat Pf-122. Setiapperlakuan terdapat 3 ulangan.
Formula dibuat dengan mengkombinasikan bahanpembawa berupa pupuk organik produk Pabrik MiniUPN Veteran Jawa Timur, dan inokulum rhizobakteriapseudomonad fluorescent isolat Pf-122. Komposisipupuk organik yang digunakan mengacu padaWuryandari et al. (2015) yaitu dengan bahan pembawaformulasi antara lain blotong, kotoran sapi, kandarwati,dan guano dengan masing-masing perbandingan 3 kg :1 kg : 3/4 kg : 1/2 kg. Semua bahan selanjutnya dicampurdengan 120 ml pseudomonad fluorescent isolat Pf-122konsentrasi 1010 CFU/ml. Semua bahan dimasukkanpada alat pembuat pupuk sehingga campuran yangterbentuk berupa beberapa formula yaitu serbuk, granul,dan pelet.
Tahapan uji yang dilakukan untuk mengetahuipengaruh formula pseudomonad fluorescent terhadappertumbuhan dan produksi tanaman cabai adalah sebagaiberikut; a). Bibit cabai yang berumur 30 hari dipindahkanke lapang yang telah diberi formula sebanyak 10 g perlubang tanam, b). Pemberian formula yang kedua,diberikan pada umur 8 minggu yaitu pada saat tanaman

158 J. HPT Tropika Vol. 17 No. 2, 2017: 156 - 161
belum membentuk fase generatif atau pembungaan, c).Inokulasi bakteri patogen Ralstonia solanacearum danFusarium oxysporum dilakukan secara alami atau tidakdilakukan inokulasi R. solanacearum dan F. oxysporumsecara buatan.
Keefektifan formula pupuk hayati dalam memacupertumbuhan dan produksi tanaman cabai akanditentukan dengan mengamati: 1). Indeks penyakit layupada tanaman cabai, 2). Pertumbuhan tanaman cabaimeliputi tinggi tanaman, jumlah cabang, dan jumlahcabang produktif, 3). Produksi cabai meliputi jumlahbuah dan berat buah.
Pengamatan perkembangan penyakit atau indekspenyakit dilakukan setiap 5 hari sampai hari ke-30.Besarnya indeks penyakit dihitung dengan rumus(Arwiyanto, 1995):
dengan:I = indeks penyakitnk = jumlah tanaman yang bergejala sakit dengan skala k (0, 1, 2, 3, 4, 5)N = jumlah total tanaman yang diinokulasiZ = kategori serangan tertinggi
Berat serangan dihitung menurut skala sebagai berikut:skor 0 = tidak terdapat gejalaskor 1 = terdapat daun layu 1-10%skor 2 = terdapat daun layu 11-30%skor 3 = terdapat daun layu 31-60%skor 4 = terdapat daun layu 61-99%skor 5 = terdapat daun layu 100%
Semua data yang diperoleh dianalisis dengananalisis varian (Anova). Uji lanjut dengan BNT padataraf kepercayaan 5%. Untuk besarnya penekanan
perkembangan penyakit dihitung dari Indeks penyakitkontrol dikurangi Indeks penyakit pada perlakuan.
HASIL DAN PEMBAHASAN
Indeks Penyakit. Selain pertumbuhan dan produksitanaman, untuk mengetahui pengaruh perlakuanpemberian formula terhadap tanaman cabai makadilakukan pengamatan terhadap indeks penyakit layupada cabai. Pada pengamatan hari ke-30 setelahperlakuan, indeks penyakit pada kontrol paling tingi yaitu55,96%. Adapun indeks penyakit pada tanaman cabaiyang diberi formula serbuk menunjukkan paling rendah,yaitu 13,23% kemudian diikuti formula granul danformula pelet yaitu 21,43 % dan 28,10 % (Tabel 1). NilaiIndeks penyakit layu di lahan penelitian di bawah 75%,Hal ini menunjukkan bahwa penyakit layu di lahanKelurahan Lebo masih kategori sedang, namun demikiantetap patut diwaspadai serangannya karena sudah diatas50%.
Apabila dibandingkan dengan kontrol, makaformula serbuk mampu menekan perkembanganpenyakit sebesar 42,73% sedangkan dengan granul danpelet hanya mampu menekan perkembangan penyakitsebesar 34,53% dan 27,86%. Hal tersebut membuktikanbahwa pemberian formula serbuk sangat berperan dalammenekan penyakit layu yang disebabkan oleh F.oxysporum dan R. solanacearum. Adanya agens hayatipseudomonad fluorescent isolat Pf-122 dalam formulaberperan menekan perkembangan patogen penyebablayu. Pada formula serbuk, populasi pseudomonadfluorescent isolat Pf-122 mungkin cukup berkembangdengan lebih baik dibandingkan pada formula yang lain.Bakteri pseudomonad fluorescent isolat Pf-122 dalamformula serbuk langsung dapat menyatu dengan tanahatau lahan sehingga populasinya dapat berkembang lebihcepat. Selain itu diduga dalam formula serbukpseudomonad fluorescent isolat Pf-122 lebih dapat
Formula Indeks penyakit hari ke 30
Serbuk 13,23 a Granul 21,43 b Pelet 28,10 c Kontrol 55,96 d
Tabel 1. Indeks penyakit layu tanaman cabai dengan perlakuan formula berbahan aktif pseudomonad fluorescentisolat Pf-122
100%NZ
k.nkI
k
0i
Angka-angka yang diikuti huruf yang berbeda dalam satu kolom menunjukkan tidak berbeda nyata berdasarkan ujiBNT taraf 5%.

Wuryandari et al. Introduksi Formula Pupuk Hayati 159
berkembang karena porositasnya lebih tinggi, tidakseperti formula pelet dan granul, sehingga kebutuhanoksigen tercukupi. Hal tersebut seperti pendapatPalleroni (1984), bahwa bakteri Pseudomonasfluorescens termasuk bakteri obligat aerob.
Adanya populasi pseudomonad fluoresent isolatPf-122 yang tinggi pada daerah perakaran akan lebihdapat menekan populasi patogen penyebab penyakit layu.Hal tersebut mungkin karena pseudomonad fluorescentisolat Pf-122 dapat menghasilkan senyawa yang dapatmenghambat patogen. Bakteri genus Pseudomonas jugamemproduksi senyawa antimikroba atau senyawapenghambat seperti phenazine, pyrrolnitrin, pyluteorin,phloroglucinols, cyclic lipopeptides, dan hydrogencyanida (Haas & Defago, 2005). Selain itu mungkindengan adanya pseudomonad fluorescent isolat Pf-122akan dapat memacu pertumbuhan tanaman sehinggameningkatkan ketahanan tanaman cabai terhadapserangan patogen layu. Keberadaan pseudomonadfluorescent yang cukup pada daerah perakaran didugadapat melarutkan fosfat memegang peranan dalammemperbaiki tanaman. Salah satu bakteri yangmerupakan pelarut potensial dari fosfat adalahPseudomonas dan Bacillus. Beberapa bakteri saprofitikyang berperan sebagai agens hayati pengendali penyakitlayu adalah bakteri Pseudomonas fluorescens padatomat, kentang, dan terung, dan Pseudomonas sp. padacabai (Saddler, 2005).
Pertumbuhan Tanaman Cabai. Untuk pengamatanpertumbuhan tanaman cabai dibedakan menjadi 2 faseyaitu pada fase vegetatif dan fase generatif. Parameterpada fase vegetatif meliputi tinggi tanaman dan jumlahcabang. Adapun pengamatan pertumbuhan pada fasegeneratif meliputi jumlah cabang produktif. Apabiladitinjau dari pertumbuhan tanaman cabai pada fasevegetatif, baik pertumbuhan tinggi tanaman maupunjumlah cabang pemberian formula serbuk menunjukkan
hasil yang paling tinggi dibandingkan formula lain dankontrol (Tabel 2). Adapun perlakuan dengan pemberianformula pelet dan granul, keduanya tidak berbeda nyatapada pertumbuhan tinggi tanaman maupun jumlahcabang. Namun demikian pemberian formula granul danpelet masih menunjukkan jumlah cabang yang lebih tinggidan berbeda nyata dengan kontrol.
Pertumbuhan tanaman cabai pada fasegeneratif yaitu jumlah cabang produktif, denganpemberian formula serbuk menunjukkan hasil yang palingbaik dibandingkan formula lain dan kontrol. Aplikasiformula granul dan pelet tidak berbeda dalampertumbuhan jumlah cabang produktif, tetapi masihberbeda nyata dibandingkan kontrol (Tabel 2).
Dengan pemberian formula serbuk berbahan aktifpseudomonad fluorescent isolat Pf-122, pertumbuhantanaman cabai menunjukkan hasil tertinggi, mungkinkarena populasi bakteri pseudomonad fluorescent isolatPf-122 dalam tanah lebih tinggi dibandingkan denganpemberian formula yang lain. Bakteri pseudomonadfluorescent isolat Pf-122 dalam formula serbuk lebihcepat tercampur dengan tanah sehingga bakteri cepatberkembang dan mengkolonisasi sistem perakaran.Pada akar, pseudomonad fluorescent isolat Pf-122diduga akan membantu pengikatan fosfor bagi tanaman,sehingga pertumbuhan akan semakin terpacu. Selain itukemungkinan rhizobakteria pseudomonad fluorescentisolat Pf-122 menghasilkan hormon pertumbuhan yangmemacu pertumbuhan tanaman cabai. Hal tersebutseperti pendapat Podile & Kishore (2007), bakterimampu memacu ketahanan sistemik denganmemproduksi fitohormon, melarutkan fosfat anorganik,dan meningkatkan pengikatan Fe dengan siderofor.Pengendalian hayati dengan mikroorganisme yangbermanfaat akan memacu pertumbuhan akar sehinggaakar menjadi lebih banyak. Mikroorganisme tanah yangdapat melarutkan fosfat memegang peranan dalammemperbaiki tanaman.
Tabel 2. Rata-rata tinggi tanaman, jumlah cabang, dan jumlah cabang produktif tanaman cabai yang diperlakukandengan 3 formulasi berbeda berbahan aktif pseudomonad fluorescent isolat Pf-122
Angka-angka yang diikuti huruf yang berbeda dalam kolom yang sama menunjukkan tidak berbeda nyata berdasarkanuji BNT taraf 5%.
Formula Tinggi tanaman (cm) Jumlah cabang Jumlah cabang produktif
Serbuk 59,47 b 21,80 c 9,67 c Granul 51,20 a 18,73 b 6,53 b Pelet 47,93 a 19,07 b 6,47 b Kontrol 47,93 a 17,67 a 5,00 a

160 J. HPT Tropika Vol. 17 No. 2, 2017: 156 - 161
Mikroorganisme tanah juga mungkinmembebaskan fosfat anorganik yang dapat larut kedalam tanah. Salah satu cara untuk memperbaikidefisiensi fosfor pada tanaman adalah dengan inokulasitanah dengan mikroorganisme pelarut fosfat. Salah satubakteri yang merupakan pelarut potensial dari fosfatadalah Pseudomonas (Rao, 1994). Banyak genusbakteri, salah satu diantaranya adalah Pseudomonas,menghasilkan suatu senyawa giberelin dan yang serupagiberelin yang dapat merangsang pertumbuhan tanaman.Bakteri ini juga mempengaruhi perkembangan rambutakar, sekresi getah, perkembangan akar lateral tanaman.
Produksi Tanaman Cabai. Untuk pengamatanterhadap produksi tanaman cabai, parameter yangdiamati meliputi jumlah buah dan berat buah. Apabiladitinjau dari produksi tanaman cabai baik jumlah buahmaupun berat buah, pemberian formula serbuk denganbahan aktif pseudomonad fluorescens Pf-122menunjukkan hasil yang paling tinggi dibandingkanformula lain dan kontrol (Tabel 3).
Apabila ditinjau dari jumlah buah, pemberianformula serbuk dapat meningkatkan hasil sampai duakali hasil dari tanaman kontrol. Sedangkan untuk hasilberat buah, pemberian formula serbuk dapatmeningkatkan hasil sampai empat kali dari tanamankontrol. Untuk pemberian formula pelet dan granul,nampak bahwa hasil jumlah buah dan berat buah jugaberbeda nyata dibandingkan kontrol. Adanyapeningkatan hasil produksi cabai pada tanaman cabaiyang diberi formula menunjukkan adanya pengaruhbaik dari bakteri pseudomonad fluorescent isolat Pf-122.Adanya populasi di dalam sistem perakaran akibataplikasi formula, kemungkinan mengkolonisasi akartanaman cabai dan dapat melarutkan fosfat yangdibutuhkan tanaman atau menghasilkan hormonpertumbuhan sehingga dapat memacu pertumbuhantanaman cabai.
Pemberian formula serbuk dibandingkan denganformula granul, dan pelet baik dari rata-rata jumlah buahdan berat buah menunjukkan nilai yang paling baik,
kemudian diikuti formula granul dan pelet. Hal tersebutmungkin disebabkan karena populasi pseudomonadfluorescent isolat Pf-122 lebih cepat berkembang dalamformula serbuk dibandingkan dengan dalam formulagranul dan pelet. Kondisi formula serbuk yang lebihporos memungkinkan lebih banyak oksigen yang dapatdiperoleh bakteri pseudomonad fluorescent isolat Pf-122, sehingga populasi lebih banyak. Sedangkan untukformula pelet, kondisi formula paling padat, sehingga haltersebut akan dapat mempengaruhi viabilitas bakterikarena sangat sedikit mendapatkan oksigen. Selain haltersebut di atas, formula pelet paling lama terdegradasiatau tercampur dengan tanah di daerah perakarandibandingkan granul dan serbuk, sehingga akanmempengaruhi bakteri dalam mengkolonisasi sistemperakaran dan membantu melarutkan fosfat untukpertumbuhan tanaman cabai.
Adanya pseudomonad fluorescens mungkin jugadapat memacu pertumbuhan dan produksi tanamancabai karena produksi hormon pertumbuhan yangdihasilkan. Hal itu sesuai dengan pendapat Pinton et al.(2001), rhizobakteria juga mampu memacu pertumbuhantanaman dengan memproduksi zat pengatur tumbuhseperti turunan auxin. Banyak genus bakteri, salah satudiantaranya adalah Pseudomonas, menghasilkan suatusenyawa giberelin dan yang serupa giberelin yang dapatmerangsang pertumbuhan tanaman. Bakteri ini jugamempengaruhi perkembangan rambut akar, sekresigetah, perkembangan akar lateral tanaman. Sebagianbesar rhizobakteria seperti Pseudomonas, Bacillus,Azotobacter memproduksi senyawa pemacupertumbuhan seperti Indolacetic acid (IAA),gibberellin, substansi seperti cytokinin (Brimecombeet al., 2001). Bakteri mampu memacu ketahanansistemik tanaman dengan memproduksi fitohormon,melarutkan fosfat anorganik, dan meningkatkanpengikatan Fe dengan siderofor (Haas & Defago,2005).
Dibandingkan hasil perlakuan dengan formulapelet dan granul, formula serbuk memberikan hasil yangterbaik. Hal tersebut diduga pada saat diaplikasikan ke
Tabel 3. Rata-rata jumlah buah dan berat buah cabai yang diperlakukan dengan formula pseudomonad fluorescentisolat Pf-122
Angka-angka yang diikuti huruf yang berbeda pada kolom yang sama menunjukkan tidak berbeda nyata berdasarkanuji BNT taraf 5%.
Formula Jumlah buah Berat buah (g) Serbuk 69,47 d 675,56 d Granul 43,93 b 300,31 b Pelet 51,00 c 379,36 c Kontrol 30,27 a 150,77 a

Wuryandari et al. Introduksi Formula Pupuk Hayati 161
dalam lubang tugal, formula serbuk lebih cepat tercampurke dalam tanah dekat sistem perakaran, sehinggapseudomonad fluorescent isolat Pf-122 lebih cepatberkembang biak dan mengkolonisasi pada systemperakaran. Selain itu dalam formula serbuk bakteripseudomonad fluorescent isolat Pf-122 lebih dapatberkembang karena kondisi tidak padat seperti formulapelet dan granul, sehingga kebutuhan oksigen tercukupidan populasi bakteri lebih tinggi. Menurut Palleroni(1984), bakteri Pseudomonas fluorescens termasukbakteri obligat aerob. Populasi pseudomonad fluorescentdalam tanah yang t inggi selanjutnya dapatmembebaskan fosfat anorganik yang dapat larut kedalam tanah, sehingga dapat memacu pertumbuhandan produksi tanaman cabai.
SIMPULAN
Introduksi formula berbahan aktif pseudomonadfluorescent isolat Pf-122 dalam bentuk serbuk memberihasil yang paling baik dalam memacu pertumbuhan danproduksi cabai di lapang.
SANWACANA
Penelitian ini dapat terlaksana atas biaya HibahPenelitian Unggulan Perguruan Tinggi Nasional No.079/SP2H/PL/Dit.Litabmas/II/2015, untuk itu kamiucapkan terima kasih kepada Direktur Penelitian danPengabdian kepada Masyarakat, Direktorat JenderalPendidikan Tinggi, Kementerian Pendidikan danKebudayaan Republik Indonesia.
DAFTAR PUSTAKA
Anonim. 2009. Budidaya Cabai Merah Keriting diLahan Kering. http://www.mojokertokota.go.id.Diakses 2014.
Arwiyanto T. 1995. Strategi pengendalian penyakit layubakteri tembakau cerutu di Sumatera Utarasecara terpadu. Ekspose Hasil PenelitianTembakau Deli IV. Medan.
Beattie GA. 2007. Survey, molecular phylogeny-genomics and recent advances. In:Gnanamanickam SS (Ed.). Plant-AssociatedBacteria. pp. 1–56. Springer, The Netherlands.
Brimecombe MJ, De Leij FA, & Lynch JM. 2001. Theeffect of root exudates on rhizosphere microbiaepopulations. In: Pinton R, Varanini Z, & NanipieriP (Eds.). The Rhizophere. Biochemistry and
Organic Substances at the Soil-PlantInterface. pp. 95–140. Marcel Dekker, Inc. NewYork–Basel.
Haas D & Defago G. 2005. Biological control of soilborne pathogens by fluorescent pseudomonads.Nat. Rev. Microbial. 3: 307–319.
Palleroni NJ. 1984. Pseudomonadaceae. Bergey’sManual of Systematic Bacteriology. In: Krieg NR& Holt JG (Eds.). pp. 141–199. The Williamsand Wilkins Co., Baltimore.
Pinton R, Varanini Z, & Nanipieri P. 2001. The rhizophereas a site of biochemical interactions among soilcomponents, plants, and microorganisms. In:Pinton R, Varanini Z, & Nanipieri P (Eds.). TheRhizophere. Biochemistry and OrganicSubstances at the Soil-Plant Interface. pp.1–18. Marcel Dekker. New York-Basel.
Podile AR & Kishore GK. 2007. Plant Growth-Promoting Rhizobacteria. In: GnanamanickamSS. (Ed.). Plant-Associated Bacteria. pp. 195–230. Springer. The Netherlands.
Rao SNS. 1994. Mikroorganisme Tanah danPertumbuhan Tanaman (Edisi kedua).Universitas Indonesia Press. Jakarta.
Saddler GS. 2005. Management of Bacterial WiltDisease. In: Allen C, Prior P, & Hayward AC(Eds.). Bacterial Wilt Disease and theRalstonia solanacearum Species Complex. pp.121–132. The American PhytopathologicalSociety Press. America.
Soelaiman V & Ernawati A. 2013. Pertumbuhan danperkembangan cabai keriting (Capsicumannuum L.) secara in vitro pada beberapakonsentrasi BAP dan IAA. Bul. Agrohorti 1: 62–66.
Wuryandari Y, Wiyatiningsih S, & Sulistyono A. 2013.Kajian Introduksi Rhizobacteria pseudomonadfluorescens terhadap Pertumbuhan dan ProduksiCabai di Lapang. Prosiding Seminar NasionalPengembangan Luaran Penelit ianMendukung Dunia Industri. pp. 37–43. 10-11Desember 2013. LPPM UPN “Veteran” JawaTimur.
Wuryandari Y, Wiyatiningsih S, & Maroeto. 2015.Formula berbahan aktif pseudomonad fluoresendan pengaruhnya terhadap perkembanganpenyakit layu pada cabai. J. HPT Tropika 15(1):89–94.

170 J. HPT Tropika Vol. 17, No. 2, 2017: 170 - 178 J. HPT Tropika. ISSN 1411-7525
Vol. 17, No. 2: 170 – 178, September 2017
AKTIVITAS SIDEROFOR BACILLUS SUBTILIS SEBAGAI PEMACUPERTUMBUHAN DAN PENGENDALI PATOGEN TANAMAN TERUNG
Nur Prihatiningsih1, Heru Adi Djatmiko1, & Puji Lestari2
1Jurusan Agroteknologi, Fakultas Pertanian, Universitas Jenderal Soedirman2Jurusan Kimia, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Jenderal Soedirman
Jl. dr. Suparno Purwokerto 53123E-mail: [email protected]
ABSTRACT
Siderophore activity of Bacillus subtilis as plant growth promoters and biological control agent of eggplants pathogens.The aims of this research were to identify the siderophores of B. subtilis, to assess its activities as plant growth promotersand biological control agent of eggplants pathogens. Five isolates of B. subtilis i.e.B46, B209, B211, B298 and B 315grown on SD-CASA medium. The isolate which showed the best siderophores production was then further studied on itsability as a growth promoter on eggplants in two soil types with different Fe content. The inhibitory test was conductedagainst two kinds of pathogens, namely Colletotrichum sp. and Ralstonia solanacearum. The greenhouse experiment wasarranged using a factorial completely randomized block design. The first factor was the B. subtilis (B. subtilis B298 andwithout B. subtilis B298), second factor was the type of soil (Ultisol and Andisol). The variables measured were Fe uptakeby plants, plant growth parameters on eggplant i.e. height, leaf number, root length, root volume, weight of fresh and driedshoot as well as fresh and dry root, percentage of inhibition to fungal and bacterial eggplant pathogens. The results showedthat the five isolates of B. subtilis were able to produce siderophores as catecholate and hydroxamate types. The bestsiderophore production was showed by B. subtilis B298. The ability of B. subtilis B298 in accelerating the growth ofplants was indicated by the increased of uptake Fe, plant height, leaf number, root volume, weight of dried plants by 45.62%,25.48%, 19.45%, 41.10% and 34.89% respectively. The inhibition to the fungal and bacterial eggplant pathogens bestshown by the isolates of B. subtilis B298 with 55.4% and 22 mm respectively.
Key words: Bacillus subtilis, siderophore, cathecolate, hydroxamate, growth of eggplants, inhibition to pathogens
ABSTRAK
Aktivitas siderofor Bacillus subtilis sebagai pemacu pertumbuhan dan pengendali patogen tanaman terung. Penelitianini bertujuan untuk mengidentifikasi siderofor dari B. subtilis, menilai aktivitasnya sebagai pemacu pertumbuhan, danpengendali patogen tanaman terung. Lima isolat B. subtilis yaitu B46, B209, B211, B298 dan B 315 ditumbuhkan padamedium SD-CASA. Isolat yang dapat memproduksi siderofor terbaik ini selanjutnya diuji kemampuannya sebagai pemacupertumbuhan tanaman terung pada dua jenis tanah dengan kandungan Fe berbeda. Uji penghambatan dilakukan terhadap duamacam patogen, yaitu Colletotrichum sp. dan Ralstonia solanacearum. Percobaan di rumah kaca dirancang menggunakanRancangan Acak Kelompok Faktorial dengan 2 faktor. Faktor pertama adalah B. subtilis (dengan B. subtilis B298 dan tanpaB. subtilis B298), faktor ke dua adalah jenis tanah (Ultisol dan Andisol). Variabel yang diamati antara lain serapan Fe olehtanaman dan parameter pertumbuhan tanaman terung meliputi tinggi tanaman, jumlah daun, panjang akar, volume akar, bobotsegar dan kering tanaman serta bobot segar dan kering akar, persentase penghambatan terhadap jamur dan bakteri patogen.Hasil penelitian menunjukkan bahwa kelima isolat B. subtilis mampu menghasilkan siderofor tipe catecholat dan hydroxamatdan produksi siderofor terbaik ditunjukkan oleh B. subtilis B298. Kemampuan B. subtilis B298 dalam memacu pertumbuhantanaman ditunjukkan dengan adanya peningkatan serapan Fe, tinggi tanaman, jumlah daun, volume akar, bobot kering tanamansecara berturut-turut sebesar 45,62%, 25,48%, 19,45%, 41,10% dan 34,89%. Penghambatan terhadap jamur dan bakteripatogen yang terbaik ditunjukkan oleh isolat B. subtilis B298 yaitu 55,4% dan 22 mm.
Kata kunci: Bacillus subtilis, siderofor, cathecolat, hydroxamat, pertumbuhan tanaman terung, penghambatan patogen

Prihatiningsih et al. Aktivitas Siderofor Bacillus subtilis 171
PENDAHULUAN
Bakteri antagonis merupakan bakteri yangmempunyai sifat mampu mengendalikan patogen danmeningkatkan pertumbuhan tanaman yang juga disebutsebagai PGPR (plant growth promoting rhizobacteria).Bakteri antagonis umumnya hidup mengkolonisasi akardan bersifat menguntungkan, karena mampumeningkatkan pertumbuhan tanaman dan menginduksiketahanan sistemik (Choudhary & Johri, 2009). Selainmengkolonisasi akar, bakteri antagonis juga berada padapermukaan akar (rizoplan) atau di dalam jaringanradikular (Kloepper et al., 1989; Dawwam et al., 2013).Salah satu bakteri yang secara luas telah dilaporkansebagai antagonis adalah Bacillus subtilis. Bakteri inimampu berperan sebagai antagonis melalui mekanismeantibiosis dan kompestisi baik ruang maupun nutrisi(Beneduzi et al., 2012).
Prihatiningsih & Djatmiko (2016) dan Lestari etal. (2017) melaporkan bahwa Bacillus subtilis asalrizosfer kentang isolat B315 (mampu menghasilkan enzimamilase) dan isolat B298 (menghasilkan kitinase) terbuktimampu menekan pertumbuhan jamur patogen tanaman.Pengendalian hayati menggunakan B. subtilis B315terhadap bakteri layu pada kentang juga telah dilakukandan menunjukkan mekanisme antibiosis (Prihatiningsihet al., 2015). Morikawa (2006) menyebutkan bahwa B.subtilis mampu menghasilkan enzim amilase, protease,pullunase, chitinase, xylanase dan lipase yang merupakanmetabolit sekunder untuk mengendalikan patogen danmemacu pertumbuhan tanaman.
Selain berperan sebagai PGPR, bakteri antagonisjuga dapat berperan sebagai biofertilizer danbioenhancer bagi tanaman. Bakteri antagonis sebagaibiofertilizer karena bakteri tersebut mampu memperbaikipertumbuhan akar dan nutrisi (Egamberdiyeva & Hoflich,2004). Peranan bakteri antagonis sebagai PGPR dapatdilihat dari kemampuannya untuk menghasilkansiderofor, IAA, sebagai pelarut fosfat, dan pemfikasasinitrogen. Siderofor adalah senyawa berbobot molekulrendah yang mampu mengkhelat besi (Fe3+), danresponsif untuk pelarutan serta pengangkutan elemen inike dalam sel bakteri (Sharma & Johri, 2003). Padakondisi keterbatasan besi, mikroorganisme penghasilsiderofor dapat mengikat dan mengangkut komplekssiderofor-besi dengan mengekspresikan protein spesifik(Nudel et al., 2001). Kelompok bakteri yang mampumenghasilkan siderofor menguntungkan untuk tanamankarena dapat menekan patogen. Besi merupakan elemenvital dalam aktivasi enzim yang mendukung ketahanantanaman terhadap patogen. Hingga saat ini terdapat dua
tipe siderofor yaitu catecholat dan hydroxamat (Neilands& Nakamura, 1991).
Dalam penelitian ini peranan siderofor yangdihasilkan oleh bakteri antagonis sebagai faktor pemacupertumbuhan tanaman diuji pada 2 jenis tanah (Ultisoldan Andisol) dengan kandungan besi yang berbeda.Tanah Ultisol dikenal juga dengan tanah podsolik merahkuning merupakan tanah marginal dengan warna oranye-merah, bersifat asam, dengan kandungan fosfor tinggi.Warna merah pada tanah ini karena kandungan Al, Fedan Mn yang tinggi. Tanah Andisol merupakan bentukandari abu vulkanik, berwarna hitam kelabu sampai cokelatdan merupakan tanah yang produktif untuk tanamanhortikultura.
Tujuan penelitian ini adalah untuk mengetahuiproduksi siderofor sebagai senyawa yang dihasilkan olehlima isolat B. subtilis asal rizosfer kentang danmengevaluasi aktivitasnya sebagai pemacu pertumbuhanserta mengendalikan jamur dan bakteri patogen tanamanterung.
METODE PENELITIAN
Tempat dan Waktu Penelitian. Penelitian dilaksanakandi Laboratorium Perlindungan Tanaman dan Rumah KacaFakultas Pertanian UNSOED dimulai Agustus 2015sampai Maret 2016.
Perbanyakan Isolat B. subtilis. Lima isolat B. subtilisyang digunakan berasal dari rizosfer kentang dengannomor isolat B46, B209, B211, B298 dan B315. Isolatini diperbanyak pada medium Yeast Peptone GlukosaAgar (YPGA) (5 g ekstrak yeast, 10 g bacteriologicalpeptone, 10 g glukosa, 20 g agar) (Lelliot & Stead, 1987).
Deteksi Siderofor yang Dihasilkan oleh B. subtilis.Pengujian sebagai penghasil siderofor dilakukanberdasarkan metode yang dilakukan oleh Shin et al.(2001) dengan menggunakan medium simple double-layered Chrome Azurol Sulphonate Agar (SD-CASA).Medium ini dibuat dengan cara: 60,5 mg Chrome azurolS (CAS) dilarutkan dalam 50 mL air suling dan dicampurdengan 10 mL larutan besi (III) (1 mmol/L FeCl3.6H2O10 mmol/L HCl), sambil dishaker, larutan ini ditambahsecara perlahan 72,9 mg HDTMA yang dilarutkan kedalam 40 mL air. Hasil larutan yang berwarna biru gelapdilarutkan dengan 2000 ml air, agar 2% w/v ditambahkanuntuk pemadat, kemudian diautoklaf 121 oC selama 15menit. Cawan Petri berdiameter 9 cm disiapkan untukplate agar (CASA) sebanyak 10 mL sebagai dasar plate.Setelah memadat dilapis lagi dengan medium Nutrient

172 J. HPT Tropika Vol. 17, No. 2, 2017: 170 - 178
Agar (NA) sebanyak 6 mL, dan diinkubasi semalam padasuhu 32 oC.
Untuk mengamati pengaruh pH terhadappembentukan siderofor digunakan paper-disc. Paper-disc steril berdiameter 5 mm diletakkan pada plate agaryang telah diinkubasi 1 malam secara aseptik. Sebanyak10 µL supernatan B. subtilis dari sentrifugasi mediumcair diteteskan pada paper-disc. Sebagai kontroldilakukan tanpa B. subtilis. Pengaruh pH terhadapkemampuan menghasilkan siderofor dilakukan pada pH5,7, dan 9. Variabel yang diamati adalah terbentuknyazona oranye dan pink/ungu pada medium yang semulaberwarna biru.
Aktivitas B. subtilis sebagai Pemacu PertumbuhanTanaman Terung. Pengujian aktivitas B. subtilis sebagaiPGPR tanaman terung dilakukan di rumah kaca padatanah Ultisol (kandungan Fe total 2391 ppm) dan Andisol(742 ppm). B. subtilis digunakan untuk perendamanbenih selama 30 menit. Perlakuan berikutnya disiramkandi sekitar tanaman pada 10, 20 dan 30 hst. Penelitiandisusun menggunakan Rancangan Acak KelompokFaktorial 2 faktor dengan 6 ulangan. Faktor ke 1 adalahB. subtilis (tanpa B. subtilis B298 dan dengan B. subtilis298). Faktor ke 2 adalah jenis tanah yaitu Ultisol danAndisol. Variabel yang diamati meliputi serapan Fe olehtanaman, tinggi tanaman, diameter batang, jumlah daun,panjang akar, volume akar, bobot tanaman segar dankering, bobot akar segar dan kering. Data yang diperolehdianalisis menggunakan sidik ragam, apabila terdapatperbedaan yang nyata dilanjutkan dengan uji Beda NyataTerkecil (BNT) dengan tingkat kepercayaan 95%.
Aktivitas Penghambatan terhadap PatogenTanaman. Pengujian penghambatan B. subtilis terhadapjamur patogen dilakukan secara dual culture berdasarkanmetode yang dikemukakan oleh Leelasuphakul et al.(2008) dan Muthukumar & Venkatesh (2013), denganmenumbuhkan secara bersama berhadapan di dalamcawan Petri yang berisi medium PDA. Potongan kolonijamur patogen Colletotrichum gloeosporioidesberdiameter 5 mm diletakkan pada medium PDAberjarak 3 cm dari tepi cawan Petri, kemudian digoreskanB. subtilis di sebelahnya dengan jarak 2,5-3 cm.Pengamatan penghambatan jamur patogen menggunakanrumus Leelasuphakul et al. (2008) dan Muthukumar &Venkatesh (2013)
dengan:I = persentase penghambatanC = pertumbuhan jari-jari koloni yang berlawanan dengan arah bakteri antagonisT = pertumbuhan jari-jari koloni yang menuju arah bakteri antagonis
Pengujian penghambatan B. subtilis terhadapbakteri patogen Ralstonia solanacearum dilakukandengan metode dua lapis medium seperti dikemukakanoleh Ghosh et al.(2007) dan Prihatiningsih & Djatmiko(2016) secara inokulasi titik, setelah 2 hari inkubasi padatutup cawan Petri diberi chloroform 0,5 mL dibiarkan2-3 jam sampai chloroform menguap. Biakan murnibakteri patogen R. solanacearum berumur 2 hari padamedium YPGA miring dipanen dengan 10 ml air steril.Sebanyak 200 ìL dimasukkan ke dalam 4 mL mediumagar air 0,6%, selanjutnya digojog dengan vortekskemudian dituang ke dalam cawan Petri tersebut sebagailapis ke dua. Setelah inkubasi 24-48 jam pada suhu28+1oC terbentuk zona terang yang diukur dari tepikoloni bakteri antagonis dalam satuan mm.
HASIL DAN PEMBAHASAN
Deteksi Siderofor yang Dihasilkan oleh B. subtilis.Hasil penelitian menunjukkan bahwa B. subtilis yangdiuji mampu berperan sebagai penghasil siderofor secarakualitatif. Pada isolat B. subtilis B46 terbentuk zonaberwarna pink/ungu dengan ukuran zona lebih kecildibandingkan dengan ukuran zona dari keempat isolatlainnya (Tabel 1). B. subtilis B209, B211, B298 danB315 menghasilkan zona oranye dengan zona terkuatadalah isolat B. subtilis B 298 (Gambar 1). Isolat B.subtilis B46 menghasilkan zona pink yang menurutPerez-Miranda et al. (2007) dan Radhakrishnan et al.(2014) adalah siderofor tipe catecholat, sedangkankeempat isolat B. subtilis B209, B211, B298 dan B315menghasilkan zona oranye adalah siderofor tipehydroxamat. Siderofor yang dihasilkan olehmikroorganisme umumnya adalah tipe hydroxamat,catecholat dan karboksilat (Ahmed & Holmstrom, 2014).
Kemampuan bakteri menghasilkan sideroformerupakan komponen penting dalam PGPR, karenasiderofor mampu mengikat besi ( Fe3+) menjadi ikatansiderofor-besi yang menjadi tersedia bagi tanaman.Ukuran zona bening atau zona ungu dan oranye yangdihasilkan menunjukkan kuat lemahnya B. subtilis dalammenghasilkan siderofor (Gambar 2). Isolat B. subtilisB298 menunjukkan zona oranye terbesar yaitu 24 mm.Siderofor yang dihasilkan oleh mikroorganisme,
I =C− T
C× 100%

Prihatiningsih et al. Aktivitas Siderofor Bacillus subtilis 173
menguntungkan tanaman karena dapat menghambatpertumbuhan patogen. Terjadinya kekurangan Fe3+ yangdibutuhkan oleh patogen karena Fe3+ sudah terikat olehsiderofor (Sharma & Johri, 2003). Selain itu besimerupakan elemen penting dalam perkembanganpenyakit, sehingga dengan terikatnya besi oleh sideroformaka patogen kurang mampu menginfeksi, sehinggamenghambat perkembangan penyakit. Jenis siderofor
yang dihasilkan B. subtilis yang terbaik adalah pada pH7. Hasil penelitian Hu & Xu (2011) mengemukakanbahwa pH 7 adalah yang terbaik untuk Bacillus strainQM3 dalam menghasilkan siderofor.
Aktivitas Siderofor B. subtilis sebagai PemacuPertumbuhan Tanaman Terung. Setelah aplikasi B.subtilis, kandungan Fe total pada dua jenis tanah yang
Gambar 2. Ukuran zona bening sebagai deteksi B. subtilis menghasilkan siderofor
Gambar 1. Siderofor yang dihasilkan oleh lima isolat B. subtilis dengan zona ungu dan oranye; A. isolat B. subtilisB46, B. isolat B. subtilis B209, C. isolat B. subtilis B211, D. isolat B. subtilis B298, E. isolatB. subtilis B315.
A B C D E
Tabel 1. Bacillus subtilis menghasilkan siderofor tipe catecholat dan hydroxamat
+ menghasilkan siderofor sedang, ++: menghasilkan siderofor kuat
Isolat B. subtilis Penghasil siderofor Warna zona Jenis siderofor B46 + ungu Catecholat B209 ++ oranye Hydroxamat B211 ++ oranye Hydroxamat B298 ++ oranye Hydroxamat B315 + oranye Hydroxamat

174 J. HPT Tropika Vol. 17, No. 2, 2017: 170 - 178
diuji (Ultisol dan Andisol) menunjukkan peningkatan dari2391 ppm menjadi 2982 ppm (Ultisol) dan 742 ppmmenjadi 854 ppm (Andisol). Aplikasi B. subtilis tersebutmampu meningkatkan Fe total berturut-turut sebesar19,81% dan 13,11%. Potensi B. subtilis B298 sebagaipemacu pertumbuhan tanaman terung didasarkan padaaktivitas siderofor yaitu serapan Fe oleh tanaman (Tabel2). Efektivitas serapan Fe lebih tinggi pada Ultisol sebesar45,62% dibanding Andisol. Jenis tanah Ultisol danAndisol memberikan pengaruh yang berbeda padavariabel serapan Fe dan luas daun. Hal ini menunjukkanbahwa Ultisol dengan kandungan Fe tinggi namun tidaktersedia bagi tanaman. Setelah aplikasi B. subtilis B298tanaman dapat menyerap Fe lebih baik pada Ultisolsebesar 45,62% dibanding pada Andisol. Serapan Fe olehtanaman pada Ultisol setelah aplikasi B. subtilismeningkat sebesar 20,34% dan Andisol 19,13%dibanding tanpa aplikasi B. subtil is. Hasil inimenunjukkan bahwa B. subtilis menghasilkan siderofor,merubah Fe total menjadi Fe tersedia bagi tanaman.
Berdasarkan Tabel 2 dapat dilihat bahwa adapengaruh interaksi antara B. subtilis dengan jenis tanahdilihat dari serapan Fe dan volume akar. Perlakuan tanahUltisol dan Andisol memberikan pengaruh yang berbedanyata pada variabel serapan Fe dan luas daun, sedangkanpada variabel lainnya tidak berpengaruh. Serapan Fe lebihtinggi peningkatannya pada Ultisol dibandingkan Andisolyaitu sebesar 20,34%. Luas daun terung meningkat18,29% dan 12,90% pada tanah Andisol dan Ultisol.
Perlakuan B. subtilis B298 menunjukkanpengaruh nyata pada serapan Fe, tinggi tanaman, jumlahdaun, bobot kering tanaman dan volume akar. AplikasiB. subtilis B298 menunjukkan adanya peningkatansebesar 19,92%, 25,48%, 19,45%, 41,10%, 34,89%secara berturut-turut pada variabel serapan Fe, tinggitanaman, jumlah daun, bobot kering tanaman dan volumeakar. Pada tanah Ultisol terjadi peningkatan volume akar57,70% setelah aplikasi B. subtilis B298. Hal inimenunjukkan bahwa B. subtilis B298 mampumenghasilkan senyawa yang dapat memacu pertumbuhantanaman seperti siderofor, IAA, dan pelarut fosfat.Siderofor yang dihasilkan B. subtilis B298 membantuketersediaan Fe bagi tanaman dan mengakibatkanpertumbuhan tanaman meningkat karena Fe dalamtanaman merupakan unsur hara esensial, berfungsi dalamfotosintesis, respirasi dan menekan serangan patogen,karena Fe juga menjadi tidak tersedia bagi patogen.
Variabel tinggi tanaman dan bobot kering tanamanterlihat meningkat disebabkan oleh tercukupinya hormontumbuh dan senyawa yang dihasilkan oleh B. subtilisyaitu IAA. Medium tanam yang baik memberikankeuntungan pada tanaman untuk menyebar dan
memperdalam perakaran (Compant et al., 2005). AplikasiB. subtilis mampu meningkatkan jumlah daun sebesar19,45% dan bobot kering tanaman sebesar 41,10%,menunjukkan bahwa tanaman mampu menyerap nutrisidengan baik. karena jumlah daun yang meningkat akanmeningkatkan fotosintat, sehingga bobot kering tanamanmeningkat. Siderofor hydroxamat yang terdapat dirizosfer secara efektif meningkatkan ketersediaan unsurFe dan P pada tanah masam. Kemampuan pengkhelatanFe tanah masam dengan kandungan Fe dan fosfat yangtinggi berimplikasi pada penyediaan P bagi tanamansekaligus dapat meningkatkan ketahanan terhadappatogen. Bobot kering tanaman menunjukkan efektivitaspenggunaan air, unsur hara dan metabolisme tanaman.Sivasakthi et al. (2014) mengatakan B. subtilis mampumeningkatkan bobot kering tanaman, yangmencerminkan banyaknya unsur hara yang terserap persatuan bobot biomassa yang dihasilkan.
B. subtilis pada Ultisol mampu meningkatkanvolume akar sebesar 57,70%. Menurut Afreen & Chavan(2014), senyawa siderofor akan mengikat Fe kemudianmentransportkan ke dalam sel, senyawa transporter yangspesifik sebagai komponen penyusun enzim nitrogenase.Besi yang terserap digunakan untuk pertumbuhan akarsehingga mampu meningkatkan biomassa akar. Sivasakthiet al. (2014) menyatakan bahwa pemberian B. subtilisdalam bentuk pupuk hayati (biofertilizer) meningkatkanjumlah daun karena membantu memfiksasi nitrogen danmengkolonisasi akar.
Penghambatan B. subtilis B298 terhadap patogentanaman in vitro ditunjukkan dengan adanya zona terang.Fenomena ini menunjukkan mekanisme penghambatandari B. subtilis B298 adalah antibiosis dengan metabolitsekunder yang dihasilkan seperti siderofor, enzim danantibiotik. Tabel 3 dan Gambar 3 menunjukkan aktivitaspenghambatan B. subtilis B298 terhadap jamur patogenC. gloeosporioides dan bakteri patogen R. solanacearum.Isolat B298 adalah yang terbaik, dengan aktivitaspenghambatan 55,4% dan 22 mm berturut-turut terhadapC. gloeosporioides dan R. solanacearum. Hal inimenunjukkan karakter B. subtilis B298 dapat sebagaiantagonis patogen tanaman. Saha et al. (2012)menyatakan bahwa B. subtilis strain AI01 dan AI03 isolatterung mampu menghambat pertumbuhan C.gloeosporioides secara berturut-turut sebesar 74,4 dan58,1%. B. subtilis BCC 6327 yang diperoleh dari“National Center for Genetic Engineering andBiotechnology (BIOTEC) Thailand mampumenghasilkan kitinase, protease dan b-1,3-glucanase yangmerupakan metabolit sekunder dan mempunyai spektrumluas sebagai aktivitas antibiotik untuk menekanpertumbuhan beberapa jamur patogen (Thakaew &
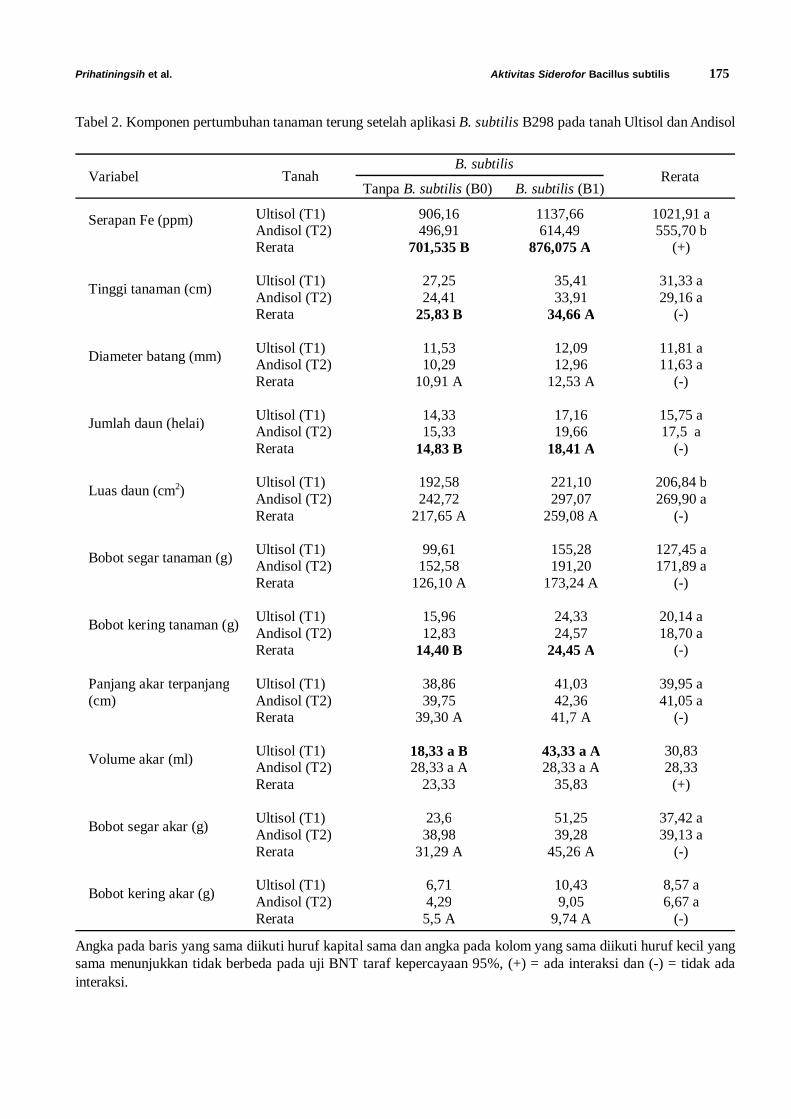
Prihatiningsih et al. Aktivitas Siderofor Bacillus subtilis 175
Tabel 2. Komponen pertumbuhan tanaman terung setelah aplikasi B. subtilis B298 pada tanah Ultisol dan Andisol
Angka pada baris yang sama diikuti huruf kapital sama dan angka pada kolom yang sama diikuti huruf kecil yangsama menunjukkan tidak berbeda pada uji BNT taraf kepercayaan 95%, (+) = ada interaksi dan (-) = tidak adainteraksi.
Variabel Tanah B. subtilis
Rerata Tanpa B. subtilis (B0) B. subtilis (B1)
Serapan Fe (ppm) Ultisol (T1) 906,16 1137,66 1021,91 a Andisol (T2) 496,91 614,49 555,70 b
Rerata 701,535 B 876,075 A (+)
Tinggi tanaman (cm) Ultisol (T1) 27,25 35,41 31,33 a Andisol (T2) 24,41 33,91 29,16 a
Rerata 25,83 B 34,66 A (-)
Diameter batang (mm) Ultisol (T1) 11,53 12,09 11,81 a Andisol (T2) 10,29 12,96 11,63 a
Rerata 10,91 A 12,53 A (-)
Jumlah daun (helai) Ultisol (T1) 14,33 17,16 15,75 a Andisol (T2) 15,33 19,66 17,5 a
Rerata 14,83 B 18,41 A (-)
Luas daun (cm2) Ultisol (T1) 192,58 221,10 206,84 b Andisol (T2) 242,72 297,07 269,90 a
Rerata 217,65 A 259,08 A (-)
Bobot segar tanaman (g) Ultisol (T1) 99,61 155,28 127,45 a Andisol (T2) 152,58 191,20 171,89 a
Rerata 126,10 A 173,24 A (-)
Bobot kering tanaman (g) Ultisol (T1) 15,96 24,33 20,14 a Andisol (T2) 12,83 24,57 18,70 a
Rerata 14,40 B 24,45 A (-) Panjang akar terpanjang (cm)
Ultisol (T1) 38,86 41,03 39,95 a Andisol (T2) 39,75 42,36 41,05 a
Rerata 39,30 A 41,7 A (-)
Volume akar (ml) Ultisol (T1) 18,33 a B 43,33 a A 30,83 Andisol (T2) 28,33 a A 28,33 a A 28,33
Rerata 23,33 35,83 (+)
Bobot segar akar (g) Ultisol (T1) 23,6 51,25 37,42 a Andisol (T2) 38,98 39,28 39,13 a
Rerata 31,29 A 45,26 A (-)
Bobot kering akar (g) Ultisol (T1) 6,71 10,43 8,57 a Andisol (T2) 4,29 9,05 6,67 a
Rerata 5,5 A 9,74 A (-)

176 J. HPT Tropika Vol. 17, No. 2, 2017: 170 - 178
Niamsup, 2013). Metabolit antifungal ditunjukkandengan adanya siderofor dan beberapa enzim hidrolitikseperti kitinase, protease, lipase dan amilase, dan strainB. subtilis AI01 dan AI03 juga mampu menghasilkanIAA sebagai pemacu pertumbuhan tanaman. Hasilpenelitian Prihatiningsih & Djatmiko (2016)memperlihatkan bahwa lima isolat B. subtilis dari rizosferkentang mampu menghasilkan IAA. Produksi kitinasedan amilase ditunjukkan oleh B. subtilis isolat B298dan B315 (Prihatiningsih & Djatmiko, 2016) dan Lestariet al. (2017).
SIMPULAN
Siderofor yang dihasilkan B. subtilis adalah tipecatecholat (isolat B46) dan hydroxamat (isolat B209,B211, B298 dan B315). Aplikasi B. subtilis B298 padatanah Ultisol meningkatkan serapan Fe sebesar 45,62%.Aktivitas siderofor dari B. subtilis B298 mampumeningkatkan tinggi tanaman, jumlah daun, bobot keringtanaman dan volume akar berturut-turut sebesar 25,48%,19,45%, 41,10% dan 34,89%. B. subtilis B298 asalrizosfer kentang dapat diaplikasikan pada tanah ultisolatau podsolik merah kuning yang dapat meningkatkanpertumbuhan tanaman, sehingga membantu mengatasipelandaian produksi tanaman pada lahan marginal.
SANWACANA
Terima kasih diucapkan kepada KementerianRiset, Teknologi, dan Pendidikan Tinggi atas dukungandananya untuk penelitian Fundamental tahun 2015-2016.
DAFTAR PUSTAKA
Ahmed E & Holmstrom SJM. 2014. Siderophores inenvironmental research: roles and applications.Microb. Biotechnol. 7: 196–208.
Afreen JM & Chavan MD. 2014. Siderophore Bacillusspp. GN-01 isolated from rhizosphere of groundnut field. Int. J. Pharm. Phytopharmacol. Res.3(4): 311–313.
Beneduzi A, Ambrosini A, & Passaglia LMP. 2012. Plantgrowth-promoting rhizobacteria (PGPR): Theirpotential as antagonists and biocontrol agents.Genet. Mol. Biol. 35(4): 1044–1051.
Choudhary DK & Johri BN. 2009. Interaction of Bacillusspp. and plants-with special reference to inducedsystemic resistance (ISR). Microb. Res. 164(5):493–513.
Isolat B. subtilis Penghambatan terhadap C. gloeosporioides (%)
Zona hambatan terhadap R. solanacearum (mm)
B46 33,3 + 0,3 14,6 + 0,4 B209 44,4 + 0,3 18,0 + 0,4 B211 45,7 + 0,3 18,2 + 0,4 B298 55,4 + 0,3 22,0 + 0,4 B315 51,5 + 0,3 20,0 + 0,4
Tabel 3. Penghambatan B. subtilis terhadap jamur dan bakteri patogen
Gambar 3. Penghambatan B. subtilis terhadap C. gloeosporioides dan R. solanacearum
B298 B315
B315
B298

Prihatiningsih et al. Aktivitas Siderofor Bacillus subtilis 177
Compant S, Duffy B, Nowak J, Clement C, & BarkaEA. 2005. Use of plant growth-promoting bacteriafor biocontrol of plant diseases: principles,mechanism of action, and future prospects. Appl.Environ. Microb. 71(9): 4951–4959.
Dawwam GE, Elbeltagy A, Emara HM, Abbas IH, &Hassan MM. 2013. Beneficial effect of plantgrowth promoting bacteria isolated from the rootsof potato plant. Ann. Agric. Sci. 58(2): 195–201.
Egamberdiyeva D & Hoflich G. 2004. Effect of plantgrowth-promoting bacteria on growth and nutrientuptake of cotton and pea on a semi-arid region ofUzbekistan. J. Arid. Environ.56(2): 293–301.
Ghosh S, Sinha A, & Sahu C. 2007. Isolation of putativeprobionts from the intestines of Indian mayorcarps. Isr. J. Aquacult-Bamid. 59(3): 127–132.
Hu Q-P & Xu J-G. 2011. A simple double-layered chromeazurol S agar (SD-CASA) plate assay to optimizethe production of siderophores by a potentialbiocontrol agent Bacillus. Afr. J. Microbiol. Res.5(25): 4321–4327.
Kloepper JW, Lifshitz R, & Zablotowicz RM. 1989. Free-living bacterial inocula for enhancing cropproductivity. Trends Biotechnol. 7(2): 39–43.
Leelasuphakul W, Hemmanee P, & Chuenchitt S. 2008.Growth inhibitory properties of Bacillus subtilisstrains and their metabolites against the greenmold pathogen (Penicillium digitatum Sacc.) ofcitrus fruit. Postharvest Biol. Technol. 48(1): 113–121.
Lelliot RA & Stead DE. 1987. Methods for TheDiagnosis of Bacterial Disease of Plant. BritishSociety for Plant Pathology by Blackwel ScientificPublication, Melbourne.
Lestari P, Prihatningsih N, & Djatmiko HA. 2017. Partialbiochemical characterization of crude extractextracellular chitinase enzyme from Bacillussubtilis B298. IOP Conference Series: MaterialsScience and Engineering. 172 012041.
Morikawa M. 2006. Beneficial biofilm formation byindustrial bacteria Bacillus subtilis and relatedspecies. J. Biosci. Bioeng. 101(1): 1–8.
Muthukumar A & Venkatesh A. 2013. Exploitaton offungal and endophytic bacteria for the managementof leaf blight of ribbon plant. J. Plant Pathol.Microb. 4(10): 209.
Neilands JB & Nakamura K. 1991. Detection,determination, isolation characterization andregulation of microbial iron chelates. In.Winkelmann G. Handbook of Microbial IronChelates. CRC Press. London.
Nudel C, Gonzalez R, Castaneda N, Mahler G, ActisLA. 2001. Influence of Iron on growth, productionof siderophore compound, membrane protein, andlipase activity in acinetobacter calcoaceticus BD413. Microbiol. Res. 155(4): 263–269.
Perez-Miranda S, Cabirol N, George-Tellez R,Zamudio-Rivera LS, & Fernandez FJ. 2007. O-CAS, a fast and universal method for siderophoredetection. J. Microbiol. Methods 70 (1): 127–131.
Prihatiningsih N, Arwiyanto T, Hadisutrisno B, & WidadaJ. 2015. Mekanisme antibiosis Bacillus subtilisB315 untuk pengendalian penyakit layu bakterikentang. J. HPT Tropika 15(1): 64–71.
Prihatiningsih N & Djatmiko HA. 2016. Enzim amilasesebagai komponen antagonis Bacillus subtilisB315 terhadap Ralstonia solanacearum kentang.J. HPT Tropika 16(1): 10–16.
Radhakrishnan M, Samshath KJ, & Balagurunathan R.2014. Hydroxamate siderophore from Bacillus sp.SD12 isolated from iron factory soil. Curr. WorldEnviron. 9(3): 990–993.
Saha D, Purkayastha GD, Ghosh A, Isha M, & Saha A.2012. Isolation and characterization of two newBacillus subtilis strains from the rhizosphere ofeggplant as potential biocontrol agents. J. PlantPathol. 94 (1): 109–118.
Sharma A, & Johri BN. 2003. Growth promotinginfluence of siderophore-producing Pseudomonasstrains GRP3A and PRS9, in maize (Zea maysL.) under iron limiting conditions. Microbiol. Res.158(3): 243–248.

178 J. HPT Tropika Vol. 17, No. 2, 2017: 170 - 178
Shin SH, Lim Y, Lee SE, Yang NW, & Rhee JH. 2001.CAS agar diffusion assay for the measurement ofsiderophore in biological fluids. J. Microbiol.Methods 44(1): 89–95.
Sivasakthi S, Usharani G, & Saranraj P. 2014. Biocontrolpotentiality of plant growth promoting bacteria(PGPR)-Pseudomonas fluorescens and Bacillussubtilis: A review. Afr. J. Agric. Res. 9(16): 1265–1277.
Thakaew R & Niamsup H. 2013. Inhibitory activity ofBacillus subtilis BCC 6327 metabolites againstgrowth of aflatoxigenic fungi isolated from birdchili powder. Int. J. Biosci. Biochem. Bioinforma.3(1): 27–31.

Efri et al. Efektivitas Fraksi Ekstrak Daun Pacar Cina 179 J. HPT Tropika. ISSN 1411-7525Vol. 17, No. 2: 179 – 184, September 2017
PENGARUH FRAKSI EKSTRAK DAUN PACAR CINA(AGLAIA ODORATA L.) TERHADAP PERTUMBUHAN COLLETOTRICHUM
CAPSICI PENYEBAB PENYAKIT ANTRAKNOSAPADA CABAI (CAPSICUM ANNUUM L.) SECARA IN VITRO
Efri, Titik Nur Aeny, Tri Maryono, & Eko Ronalddi
Jurusan Proteksi Tanaman, Fakultas Pertanian, Universitas LampungJl. Prof. Dr. Sumantri Brodjonegoro No 1, Bandar Lampung 35145
E-mail: [email protected]
ABSTRACT
Influence of Aglaia odorata L. leaf extract on in vitro growth of chilli antrachnose fungi (Colletotrichum capsici). Pacarcina (Aglaia odorata L.) is known as one of the potential herbs as fungicides. The objective of this study was to determine theeffectiveness of various leaf extract fractions of pacar cina as a fungicide to suppress the in vitro growth of C. capsici, thecausal agent of anthracnose disease on chili. This study consisted of two sub-experiments. The first experiment consisted of12 treatments: leaf extract fractions with distilled water solvent, leaf extract fractions with ethanol solvent with a concentrationof 10, 20, 30, 40, 50, 60, 70, 80 and 90%, propineb 0.2 % (synthetic fungicides) and control (without fungicide). The secondexperiment consisted of 12 treatments: leaf extract fractions with distilled water solvent, leaf extract fractions with the solventof n-hexane at a concentration of 10, 20, 30, 40, 50, 60, 70, 80 and 90%, propineb 2% (synthetic fungicides), and control (withoutfungicides). The treatmentswerearranged by completely randomized design (CRD) with three replications. Variables measuredin this experiment were colony diameter and density of spores of C. capsici. Data were analyzed by analysis of variance(ANOVA) and differences between the mean values was tested by Duncan Multiple Range Test (DMRT) with the level of 5%significant different. Additionally, leaf extract that effectively inhibited the growth of C. capsici was the fraction of the leafextract with 80% ethanol solvent and 10%, 60%, and 90% n-hexanesolvent. The result showed that the leaf extract fraction ofpacar cina was potentially used as fungicide to inhibit the growth of C. capsici.
Key words: Aglaia odorata L., anthracnose, Capsicum annuum L., Colletotrichum capsici, growth inhibition
ABSTRAK
Pengaruh fraksi ekstrak daun pacar cina (Aglaia odorata L.) terhadap penghambatan pertumbuhan Colletotrichumcapsici penyebab penyakit antraknosa pada cabai (Capsicum annuum L.) secara in vitro. Pacar cina merupakan salah satutumbuhan yang berpotensi sebagai fungisida nabati. Penelitian ini bertujuan untuk mengetahui efektivitas berbagai fraksiekstrak daun pacar cina sebagai fungisida nabati dalam menekan pertumbuhan C. capsici penyebab penyakit antraknosapada cabai secara in vitro. Penelitian terdiri dari dua sub percobaan. Percobaan pertama terdiri dari 12 perlakuan yaitu, ekstrakdaun pacar cina dari fraksi pelarut aquades, pelarut alkohol dengan konsentrasi 10, 20, 30, 40, 50, 60, 70, 80, dan 90%, propineb(fungisida sintetik) dan kontrol (tanpa fungisida). Percobaan kedua terdiri dari 12 perlakuan yaitu ekstrak daun pacar cina darifraksi pelarut aquades, pelarut n-hexana dengan konsentrasi 10%, 20%, 30, 40%, 50%, 60%, 70%, 80%, dan 90%, fungisidapropineb, dan kontrol. Perlakuan dalam percobaan disusun dalam Rancangan Acak Lengkap (RAL) dengan tiga ulangan.Peubah yang diamati adalah pertumbuhan diameter koloni dan kerapatan spora C. capsici. Data yang diperoleh dianalisisdengan sidik ragam (Anova) dan perbedaan nilai tengah diuji dengan uji Duncan Multiple Range Test (DMRT) pada tarafnyata 5%. Fraksi ekstrak daun pacar cina yang efektif menghambat pertumbuhan in vitro C. capsici adalah fraksi ekstrakdengan pelarut alkohol 80% dan pelarut n-hexana 10%, 60%, dan 90%. Hasil penelitian menunjukkan bahwa ekstrak daunpacar cina berpotensi sebagai bahan fungisida nabati yang menghambat pertumbuhan C. capsici.
Kata kunci: Aglaia odorata L., antraknosa, Capsicum annuum L., Colletotrichum capsici, penghambatan pertumbuhan

180 J. HPT Tropika Vol. 17 No. 2, 2017: 105 - 110
PENDAHULUAN
Cabai adalah tanaman asli wilayah tropika dansubtropika Amerika. Cabai merah (Capsicum annuumL.) adalah spesies yang paling luas dibudidayakan karenamerupakan salah satu komoditas sayuran yang sangaterat hubungannya dalam kehidupan masyarakat sehari-hari (Rubatzky, 2000). Salah satu kendala dalambudidaya cabai adalah serangan penyakit antraknosabuah cabai yang disebabkan oleh jamur Colletotrichumcapsici L. (Semangun, 2000). Penyakit antraknosamerupakan salah satu penyakit penting pada cabaikarena dapat menimbulkan kerugian yang besar, tidakhanya di lapangan tetapi juga pada saat pasca panen.Pakdeevaraporn et al. (2005) melaporkan bahwakehilangan hasil akibat penyakit antraknosa dapatmencapai 50%. Serangan patogen antraknosa adalahsalah satu penyebab menurunnya produksi cabai diProvinsi Lampung. Pada tahun 2001 produksi cabaimerah di Provinsi Lampung sebesar 44.374 ton, tetapipada tahun 2012 menurun menjadi 42.437 ton (BadanPusat Statistik, 2012). Salah satu cara pengendalian yangbanyak dilakukan petani adalah dengan penyemprotanfungisida sintetik secara intensif.
Pengendalian dengan fungisida sintetikmemerlukan biaya yang besar dan efek residunya dapatmenimbulkan dampak negatif baik bagi lingkunganmaupun manusia (Than et al., 2008). Oleh karenanyapenggunaan fungisida nabati yang ramah lingkungan danaman bagi manusia perlu diterapkan. Dari penelitiansebelumnya juga diketahui bahwa daun tanamanmengkudu juga dapat menghambat pertumbuhan C.capsici penyebab antraknosa cabai (Efri, 2010). Harnaset al. (2009) melaporkan bahwa ekstrak rumput tekiyang memiliki kandungan alkaloid, saponin, flavonoid,tanin dan minyak atsiri efektif menekan pertumbuhanjamur Candida albicans. Menurut Sudarmo (2005),daun pacar cina memiliki kandungan alkaloid, saponin,flavonoid, tanin, dan minyak atsiri. Wang & Yang(2013) melaporkan bahwa daun pacar cina mengandungmetabolit sekunder yang mampu berperan sebagaiinsektisida maupun fungisida. Oleh karena itu perludilakukan kajian ilmiah untuk mengetahui potensi daunpacar cina sebagai fungisida nabati dalam menekanpertumbuhan jamur C. capsici. Sebagai tahap awal,penelitian ini dilakukan dengan tujuan untuk mendapatkanfraksi (senyawa) ekstrak daun pacar cina yang efektifmenekan pertumbuhan jamur C. capsici penyebabpenyakit antraknosa pada cabai secara in vitro.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilaksanakan diLaboratorium Penyakit Tumbuhan Fakultas PertanianUniversitas Lampung. Pelaksanaan penelitian dimulaibulan Juni sampai September 2013.
Rancangan Percobaan. Penelitian terdiri dari dua subpercobaan. Percobaan pertama terdiri dari 12 perlakuanyaitu ekstrak daun pacar cina dari fraksi pelarut aquades,pelarut alkohol dengan konsentrasi 10, 20, 30, 40, 50,60, 70, 80, dan 90%, propineb konsentrasi 0,2%(fungisida sintetik) dan kontrol (tanpa fungisida).Percobaan kedua dengan perlakuan ekstrak daun pacarcina dari fraksi pelarut aquades, pelarut n-hexana dengankonsentrasi 10, 20, 30, 40, 50, 60, 70, 80, dan 90%,fungisida propineb konsentrasi 0,2%, dan kontrol.Perlakuan dalam percobaan disusun dalam RancanganAcak Lengkap (RAL) dengan tiga ulangan. Data hasilpengamatan dianalisis dengan sidik ragam (Anova), danperbedaan nilai tengah antar perlakuan diuji dengan ujiDuncan Multiple Range Test (DMRT) pada tarafnyata 5% menggunakan program Statistical AnalysisSystem (SAS).
Ekstraksi dan Fraksinasi Daun. Daun pacar cinayang digunakan diperoleh dari daerah Talang Padang,Kabupaten Tanggamus. Ekstraksi daun pacar cinadilakukan dengan cara sebagai berikut: daun segarseberat 100 g dicuci dengan air bersih lalu dikering-anginkan dan kemudian diblender sampai halus.Selanjutnya daun pacar cina yang sudah diblenderdifraksinasi secara bertingkat menggunakan alatpenyaring yang terbuat dari paralon berbagai ukuransehingga membentuk corong yang berisi arang aktifsebagai filter dan penjerab (adsorbsi) senyawa bersifatnonpolar. Proses ekstraksi dimulai dari aquades sebagaikonsentrasi 0% yang diteruskan dengan larutan alkoholatau n-hexana masing-masing dengan konsentrasi 10,20, 30, 40, 50, 60, 70, 80, dan 90%.
Pengujian Penghambatan. Pengukuran diameterkoloni jamur C. capsici dilakukan untuk mengetahuipenghambatan laju pertumbuhan vegetatif jamur C.capsici pada media PSA (Potato Sucrose Agar) yangtelah dicampur dengan masing-masing fraksi ekstrakkering daun pacar cina. Pengujian penghambatandilakukan dengan cara menumbuhkan koloni jamur C.capsici berumur 10 hari dan berukuran 5 mm pada

Efri et al. Efektivitas Fraksi Ekstrak Daun Pacar Cina 181
cawan Petri media PSA sesuai perlakuan. Selanjutnyacawan Petri diinkubasi pada suhu ruang selama 7 hari.Diameter koloni jamur C. capsici diamati setiap harimulai hari ke-2 sampai hari ke-7 setelah inokulasi,sedangkan kerapatan spora jamur C. capsici diamatipada hari ke-7. Diameter jamur dihitung dari rata-ratapengukuran diameter dari empat arah yang berbeda.Kerapatan spora jamur dihitung dengan metode hitunglangsung menggunakan haemocytometer dengan rumusSyahnen et al. (2014):
dengan:S = Jumlah sporaR = Jumlah rata-rata spora pada 5 bidang
pandang haemocytometerK = Konstanta koefisien alat (2,5 x 105)F = Faktor pengenceran yang dilakukan
HASIL DAN PEMBAHASAN
Pertumbuhan Jamur C. capsici. Hasil uji statistikmenunjukkan bahwa ekstrak daun pacar cina dalampelarut alkohol dengan berbagai tingkat konsentrasi danfungisida sintetik dapat menekan pertumbuhan jamur C.capsici (Tabel 1). Dari data yang diperoleh, pengaruhperlakuan dapat digolongkan menjadi 3 kategori, yaitu :
(1) menghambat pertumbuhan jamur, (2) memacupertumbuhan jamur, dan (3) tidak berpengaruh terhadappertumbuhan jamur.
Perlakuan yang dapat menghambat pertumbuhanjamur yaitu ekstrak daun pacar cina dengan pelarutaquades, alkohol 40%, alkohol 50%, dan alkohol 80%.Perlakuan yang sangat berpotensi adalah ekstrak daunpacar cina dengan pelarut alkohol 80% karenakeefektifannya sama dengan perlakuan menggunakanfungisida kimia. Reaksi penghambatan yang ditunjukkanoleh 4 perlakuan tersebut diduga karena adanyasenyawa aktif anti jamur dalam daun pacar cina sepertisaponin, alkaloid, flavonoid, tanin, dan minyak atsiriyang terlarut dalam ekstrak kasar daun pacar cina.Menurut Soetan et al. (2006), saponin dan flavonoiddapat membentuk kompleks dengan protein ekstraselulerdan juga dinding sel jamur sehingga menyebabkanmembran sel jamur terganggu. Minyak atsiri dapatmempengaruhi permeabilitas dan reaksi enzim yangterdapat pada sel jamur sehingga menyebabkanmembran sel lisis dan mati (Ridawati et al., 2011).Senyawa alkaloid mampu merusak membran seldengan cara mendenaturasi protein sehingga membransel lisis dan mati, sedangkan tanin akan bereaksi denganlipid dan asam amino yang terdapat pada dinding seljamur sehingga dinding sel akan rusak dan tanin akanmasuk ke dalam inti sel jamur. Senyawa tanin yang
Tabel 1. Rata-rata diameter pertumbuhan C. capsici dalam media yang dicampur ekstrak pacar cina yang diekstraksidengan pelarut alkohol
Perlakuan
Rata-rata diameter pertumbuhan C. capsici pada hari ke
2 hsi 3 hsi 4 hsi 5 hsi 6 hsi 7 hsi
----------------------------------------------cm-----------------------------------------
Kontrol 1,66 a 2,33 b 3,07 bc 3,41 bc 3,78 cd 4,31 bc Fungisida 0,50 b 0,50 c 0,50 g 0,50 f 0,50 g 0,50 f Aquades 0,81 b 0,98 c 1,18 f 1,51 e 1,97 f 2,43 e Daun pacar cina + alkohol 10% 1,68 a 2,58 abc 3,63 ab 3,90 ab 4,84 ab 5,85 a Daun pacar cina + alkohol 20% 1,90 a 2,31 b 2,85 cd 3,32 bc 3,45 de 3,53 cd Daun pacar cina + alkohol 30% 1,87 a 2,57 abc 3,13 bc 3,67 b 4,12 bcd 4,41 bc Daun pacar cina + alkohol 40% 1,73 a 2,33 b 2,42 de 2,57 cd 2,78 ef 2,82 de Daun pacar cina + alkohol 50% 1,78 a 2,06 b 2,16 e 2,36 d 2,39 f 2,51 de Daun pacar cina + alkohol 60% 1,98 a 2,91 a 3,02 bcd 3,66 b 4,05 bcd 4,50 bc Daun pacar cina + alkohol 70% 1,81 a 2,41 abc 3,46 abc 4,01 ab 4,51 abc 5,17 ab Daun pacar cina + alkohol 80% 0,67 b 0,70 c 1,00 f 1,00 ef 1,00 g 1,00 f Daun pacar cina + alkohol 90% 2,08 a 2,92 a 3,80 a 4,60 a 5,19 a 5,77 a
F hitung 15,9* 25,9* 28,8* 21,6* 28,2* 26,9*
*berbeda pada taraf nyata 5% ; hsi = hari setelah inokulasi ; angka yang diikuti huruf yang sama tidak berbedanyata setelah diuji dengan Uji Duncan pada taraf nyata 5%.
S = R × K × F
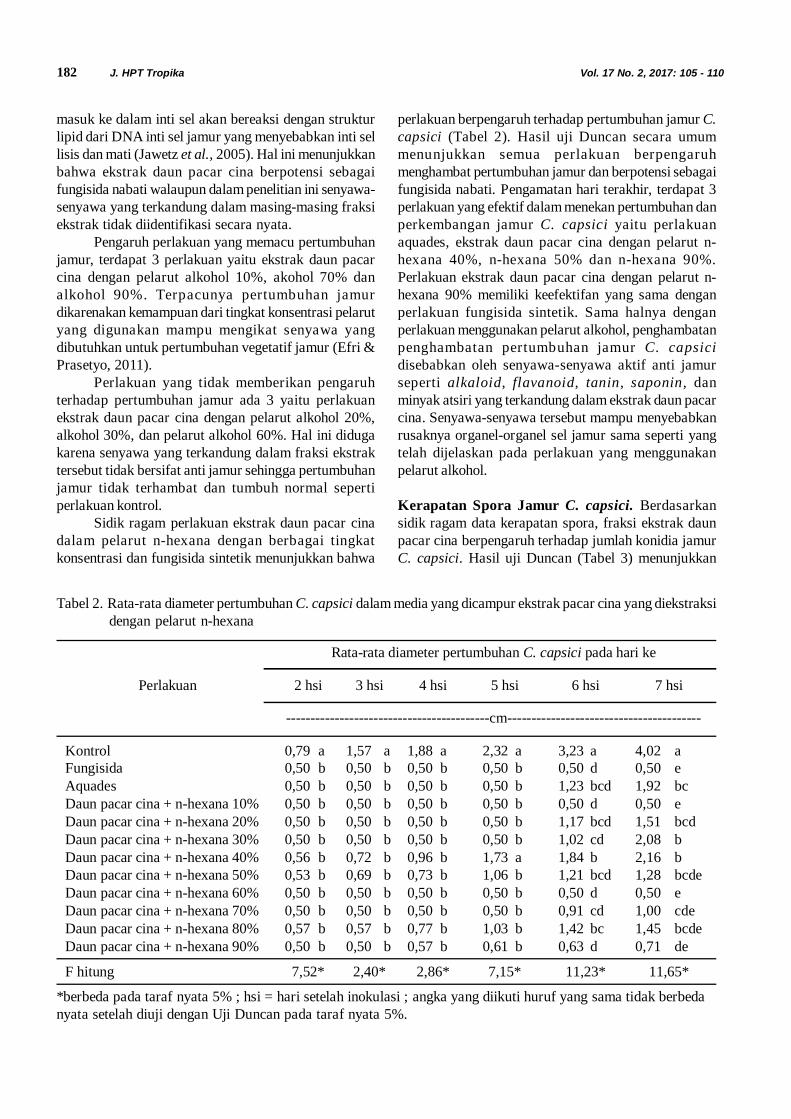
182 J. HPT Tropika Vol. 17 No. 2, 2017: 105 - 110
masuk ke dalam inti sel akan bereaksi dengan strukturlipid dari DNA inti sel jamur yang menyebabkan inti sellisis dan mati (Jawetz et al., 2005). Hal ini menunjukkanbahwa ekstrak daun pacar cina berpotensi sebagaifungisida nabati walaupun dalam penelitian ini senyawa-senyawa yang terkandung dalam masing-masing fraksiekstrak tidak diidentifikasi secara nyata.
Pengaruh perlakuan yang memacu pertumbuhanjamur, terdapat 3 perlakuan yaitu ekstrak daun pacarcina dengan pelarut alkohol 10%, akohol 70% danalkohol 90%. Terpacunya pertumbuhan jamurdikarenakan kemampuan dari tingkat konsentrasi pelarutyang digunakan mampu mengikat senyawa yangdibutuhkan untuk pertumbuhan vegetatif jamur (Efri &Prasetyo, 2011).
Perlakuan yang tidak memberikan pengaruhterhadap pertumbuhan jamur ada 3 yaitu perlakuanekstrak daun pacar cina dengan pelarut alkohol 20%,alkohol 30%, dan pelarut alkohol 60%. Hal ini didugakarena senyawa yang terkandung dalam fraksi ekstraktersebut tidak bersifat anti jamur sehingga pertumbuhanjamur tidak terhambat dan tumbuh normal sepertiperlakuan kontrol.
Sidik ragam perlakuan ekstrak daun pacar cinadalam pelarut n-hexana dengan berbagai tingkatkonsentrasi dan fungisida sintetik menunjukkan bahwa
perlakuan berpengaruh terhadap pertumbuhan jamur C.capsici (Tabel 2). Hasil uji Duncan secara umummenunjukkan semua perlakuan berpengaruhmenghambat pertumbuhan jamur dan berpotensi sebagaifungisida nabati. Pengamatan hari terakhir, terdapat 3perlakuan yang efektif dalam menekan pertumbuhan danperkembangan jamur C. capsici yaitu perlakuanaquades, ekstrak daun pacar cina dengan pelarut n-hexana 40%, n-hexana 50% dan n-hexana 90%.Perlakuan ekstrak daun pacar cina dengan pelarut n-hexana 90% memiliki keefektifan yang sama denganperlakuan fungisida sintetik. Sama halnya denganperlakuan menggunakan pelarut alkohol, penghambatanpenghambatan pertumbuhan jamur C. capsicidisebabkan oleh senyawa-senyawa aktif anti jamurseperti alkaloid, flavanoid, tanin, saponin, danminyak atsiri yang terkandung dalam ekstrak daun pacarcina. Senyawa-senyawa tersebut mampu menyebabkanrusaknya organel-organel sel jamur sama seperti yangtelah dijelaskan pada perlakuan yang menggunakanpelarut alkohol.
Kerapatan Spora Jamur C. capsici. Berdasarkansidik ragam data kerapatan spora, fraksi ekstrak daunpacar cina berpengaruh terhadap jumlah konidia jamurC. capsici. Hasil uji Duncan (Tabel 3) menunjukkan
Tabel 2. Rata-rata diameter pertumbuhan C. capsici dalam media yang dicampur ekstrak pacar cina yang diekstraksidengan pelarut n-hexana
*berbeda pada taraf nyata 5% ; hsi = hari setelah inokulasi ; angka yang diikuti huruf yang sama tidak berbedanyata setelah diuji dengan Uji Duncan pada taraf nyata 5%.
Perlakuan
Rata-rata diameter pertumbuhan C. capsici pada hari ke
2 hsi 3 hsi 4 hsi 5 hsi 6 hsi 7 hsi
------------------------------------------cm----------------------------------------
Kontrol 0,79 a 1,57 a 1,88 a 2,32 a 3,23 a 4,02 a Fungisida 0,50 b 0,50 b 0,50 b 0,50 b 0,50 d 0,50 e Aquades 0,50 b 0,50 b 0,50 b 0,50 b 1,23 bcd 1,92 bc Daun pacar cina + n-hexana 10% 0,50 b 0,50 b 0,50 b 0,50 b 0,50 d 0,50 e Daun pacar cina + n-hexana 20% 0,50 b 0,50 b 0,50 b 0,50 b 1,17 bcd 1,51 bcd Daun pacar cina + n-hexana 30% 0,50 b 0,50 b 0,50 b 0,50 b 1,02 cd 2,08 b Daun pacar cina + n-hexana 40% 0,56 b 0,72 b 0,96 b 1,73 a 1,84 b 2,16 b Daun pacar cina + n-hexana 50% 0,53 b 0,69 b 0,73 b 1,06 b 1,21 bcd 1,28 bcde Daun pacar cina + n-hexana 60% 0,50 b 0,50 b 0,50 b 0,50 b 0,50 d 0,50 e Daun pacar cina + n-hexana 70% 0,50 b 0,50 b 0,50 b 0,50 b 0,91 cd 1,00 cde Daun pacar cina + n-hexana 80% 0,57 b 0,57 b 0,77 b 1,03 b 1,42 bc 1,45 bcde Daun pacar cina + n-hexana 90% 0,50 b 0,50 b 0,57 b 0,61 b 0,63 d 0,71 de
F hitung 7,52* 2,40* 2,86* 7,15* 11,23* 11,65*

Efri et al. Efektivitas Fraksi Ekstrak Daun Pacar Cina 183
bahwa perlakuan ekstrak daun pacar cina memberikanpengaruh yang berbeda antar perlakuan. Kerapatanspora terkecil ditunjukkan oleh perlakuan yangmenggunakan fungisida sintetik dan perlakuan ekstrakdengan pelarut alkohol 80%. Uji Duncan menunjukkanperlakuan ekstrak dengan pelarut alkohol 80% memilikikeefektifan yang hampir menyamai keefektifan fungisidasintetik. Hasil uji Duncan (Tabel 1 dan Tabel 3) secaraumum menunjukkan bahwa panjang diameterpertumbuhan jamur sejalan dengan kerapatan spora. Uji
Duncan pada Tabel 1 menunjukkan bahwa perlakuanyang menggunakan pelarut aquades, alkohol 40%,alkohol 50% dan alkohol 80% mampu menekanpertumbuhan diameter jamur C. capsici. Hal ini sejalandengan hasil uji Duncan pada Tabel 2 yang menunjukkanperlakuan yang menggunakan pelarut alkohol 40%,alkohol 50% dan alkohol 80% juga mampu menekankerapatan spora jamur C. capsici kecuali perlakuanaquades.
Tabel 3. Kerapatan spora C. capsici dalam media yang dicampur ekstrak pacar cina yang diekstraksi denganpelarut alkohol pada 7 hsi
Perlakuan Kerapatan spora C. capsici (x 104 spora/ml)
Kontrol 10,88 e Fungisida 0 i Aquades 14,19 c Daun pacar cina + alkohol 10% 14,90 bc Daun pacar cina + alkohol 20% 9,34 f Daun pacar cina + alkohol 30% 11,21 de Daun pacar cina + alkohol 40% 2,12 g Daun pacar cina + alkohol 50% 2,11 g Daun pacar cina + alkohol 60% 11,82 d Daun pacar cina + alkohol 70% 15,13 ab Daun pacar cina + alkohol 80% 0,88 h Daun pacar cina + alkohol 90% 15,77 a
F hitung 611,33*
*berbeda pada taraf nyata 5% ; hsi = hari setelah inokulasi ; angka yang diikuti huruf yang sama tidak berbedanyata setelah diuji dengan Uji Duncan pada taraf nyata 5%.
*berbeda pada taraf nyata 5% ; hsi = hari setelah inokulasi ; angka yang diikuti huruf yang sama tidak berbedanyata setelah diuji dengan Uji Duncan pada taraf nyata 5%.
Tabel 4. Kerapatan spora C. capsici dalam media yang dicampur ekstrak pacar cina yang diekstraksi denganpelarut n-hexana pada 7 hsi
Perlakuan Kerapatan spora C. capsici (x 104 spora/ml)
Kontrol 7,70 a Fungisida 0 f Aquades 2,64 c Daun pacar cina + n-hexana 10% 0 f Daun pacar cina + n-hexana 20% 0,58 e Daun pacar cina + n-hexana 30% 4,80 b Daun pacar cina + n-hexana 40% 1,47 d Daun pacar cina + n-hexana 50% 1,22 d Daun pacar cina + n-hexana 60% 0 f Daun pacar cina + n-hexana 70% 0,67 e Daun pacar cina + n-hexana 80% 0,70 e Daun pacar cina + n-hexana 90% 0,17 f
F hitung 363,74*

184 J. HPT Tropika Vol. 17 No. 2, 2017: 105 - 110
Sidik ragam menunjukkan bahwa perlakuanekstrak dengan pelarut n-hexana dengan berbagaitingkat konsentrasi dan perlakuan yang menggunakanfungisida sintetik memberikan pengaruh yang nyataterhadap kerapatan spora jamur C. capsci. Uji Duncan(Tabel 4) menunjukkan perlakuan ekstrak dengan pelarutn-hexana 10% dan n-hexana 60% tidak menghasilkanspora, sama dengan perlakuan fungisida sintetik.Perlakuan ekstrak dengan pelarut n-hexana 90%memiliki efektifitas yang sama dengan perlakuan ekstrakyang menggunakan pelarut n-hexana 10% dan n-hexana60% walau masih memiliki spora. Sedangkan nilaikerapatan spora terkecil pada setiap perlakuan, jikadibandingkan dengan kontrol ditunjukkan oleh perlakuanekstrak dengan pelarut n-hexana 20%, n-hexana 70%dan n-hexana 80%.
SIMPULAN
Ekstrak daun pacar cina berpotensi sebagai bahanfungisida nabati yang menghambat pertumbuhan C.capsici. Ekstrak daun pacar cina yang efektifmenghambat petumbuhan in vitro C. capsici adalahfraksi pelarut aquades, alkohol konsentrasi 40, 50, &80% dan pelarut n-hexana konsentrasi 40%, 50%, dan90%.
SANWACANA
Penelitian ini terlaksana atas bantuan dana dariFakultas Pertanian Universitas Lampung melalui danaDIPA 2013-2014.
DAFTAR PUSTAKA
Badan Pusat Statistik. 2012. Produksi Cabai MerahMenurut Provinsi. Badan Pusat StatistikIndonesia. Jakarta.
Efri & Prasetyo J. 2011. In vitro effectivity test of activeingredients of biofungicide compound from someextract fractions of mengkudu leaf on growth ofColletotrichum capsici pathogen of antracnosedisease on chilli piper. Prosiding SeminarNasional Sains dan Teknologi IV. pp. 267–278.Bandar Lampung. 29-30 November 2011.
Efri. 2010. Pengaruh ekstrak berbagai bagian tanamanmengkudu (Morinda citrifolia) terhadapperkembangan penyakit antraknosa padatanaman cabe (Capsicum annuum L.). J. HPTTropika 10(1): 52–58.
Harnas EDMD, Winarsih S, & Nurdiana. 2009. EfekAntifungi Ekstrak Etanol Rumput Teki (Cyperusrotundus L.) terhadap Candida albicans isolatvaginitis secara in vitro. Laporan Penelitian.Fakultas Kedokteran Universitas Brawijaya.Malang.
Jawetz E, Melnick GE, & Adelberg CA. 2005.Mikrobiologi Kedokteran. Edisi I.Diterjemahkan oleh Penerjemah BagianMikrobiologi Fakultas Kedokteran UniversitasAirlangga. Surabaya.
Pakdeevaraporn P, Wasee S, Taylor PWJ, &Mongkolporn O. 2005. Inheritance of resistanceto anthracnose caused by Colletotrichum capsiciin Capsicum. Plant Breeding 124(2): 206–208.
Ridawati, Jenie BSL, Djuwita I, & Sjamsuridzal W. 2011.Aktivitas antifungal minyak atsiri jinten putihterhadap Candida parapsilosis SS25, C.orthopsilosis NN14, C. metapsilosis MP27, danC. etchellsii MP18. Makara Sains 15(1): 58–62.
Rubatzky VE. 2000. Sayuran Dunia 3. InstitutTeknologi Bandung. Bandung.
Semangun H. 2000. Penyakit-Penyakit TanamanHortikultura di Indonesia. Gadjah MadaUniversity Press. Yogyakarta.
Syahnen, Sirait DDN, & Pinem SEB. 2014. Teknik UjiMutu Agens Pengendali Hayati (APH) diLaboratorium. Laboratorium Lapangan BalaiBesar Perbenihan dan Proteksi TanamanPerkebunan (BBPPTP). Medan.
Soetan KO, Oyekunle MA, Aiyelaagbe O, & FafunsoMA. 2006. Evaluation of the antimicrobial activityof saponins extract of Shorgum bicolor L.Moench. Afr. J. Biotechnol 5(23): 2405–2407.
Sudarmo S. 2005. Teknologi Tepat Guna PestisidaNabati Pembuatan dan Pemanfaatannya .Kanisius. Yogyakarta.
Than PP, Jeewon R, Hyde KD, Pongsupasamit S,Mongkolporn O, & Taylor PWJ. 2008.Characterization and pathogenicity ofColletotrichum species associated withanthracnose on chilli (Capsicum spp.) in Thailand.Plant Pathol. 57(3): 562–572.
Wang DX & Yang SM. 2013. Chemical constituentsfrom the leaves of Aglaia odorata. Z.Naturforsch. 68(3–4): 82–86.

162 J. HPT Tropika Vol. 17, No. 2, 2017: 162 - 169 J. HPT Tropika. ISSN 1411-7525
Vol. 17, No. 2: 162–169, September 2017
KESINTASAN BEBERAPA JAMUR ANTAGONIS PADA BUAHCABAI DAN POTENSINYA DALAM MENEKAN PENYAKIT
ANTRAKNOSA YANG DISEBABKAN OLEHCOLLETOTRICHUM GLOEOSPORIOIDES
Nurbailis, Martinius, & Rizka Naipinta
Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas AndalasKampus Limau Manis, Padang
E-mail: [email protected]
ABSTRACT
Persistence of several antagonistic fungus on chilli and its potential to suppress anthracnose disease caused byColletotrichum gloeosporioides. Anthracnose disease caused by C. gloeosporioides is one of the important diseases onchilli because its directly gives negative impact on chilli production. The aim of this research was to obtain the superiorantagonistic fungi that have ability to persist on chili fruit and potential to control anthracnose disease caused by C.gloeosporioides The experiment consist of two units: 1. Testing of persistence ability of antagonistic fungi on chilli fruit, 2.Testing the potential of antagonistic fungi to suppress anthracnose disease on chilli fruit. Both of the test used RandomizedBlock Design (RBD) with 10 treatments and 4 replication, each of replication consist of 4 chilies fruit. Those treatments wereTrichoderma-PP1, Trichoderma-PP3, Trichoderma-AG2, Trichoderma-PYK3, Paecilomyces-PP6, Paecilomyces-PP7,Paecilomyces-AG4, Paecilomyces-PYK4, Aspergillus PP2 and without antagonistic fungi (control). The result indicated thatall antagonistic fungi isolate could persist on chili fruit. The highest persistence were Trichoderma-PP3 and Trichoderma-AG2 (95.83%) and the lowest belonged to Paecilomyces-PP7 (50%). Trichoderma-PP3 and Trichoderma-AG2 were the bestisolates for suppressing anthracnose disease caused by C. gloeosporioides.
Key words : anthracnose, Aspergillus, colonization, Paecilomyces, Trichoderma
ABSTRAK
Kesintasan beberapa jamur antagonis pada buah cabai dan potensinya dalam menekan penyakit antraknosa yangdisebabkan oleh Colletotrichum gloeosporioides. Penyakit antraknosa yang disebabkan oleh Colletotrichum gloeosporioidesmerupakan salah satu penyakit penting pada cabai karena menyerang buah yang berakibat langsung pada produksi. Tujuanpenelitian ini adalah untuk mendapatkan jamur antagonis unggul yang mampu tumbuh dan bertahan pada buah cabai sertaefektif menekan penyakit antraknosa yang disebabkan oleh C. gloeosporioides. Penelitian terdiri dari 2 unit percobaan: 1. Ujikesintasan jamur antagonis pada buah cabai. 2. Uji potensi jamur antagonis terhadap penekanan penyakit antraknosa padabuah cabai. Rancangan yang digunakan adalah Acak Kelompok (RAK) yang terdiri dari 10 perlakuan dan 4 ulangan.Perlakuannya adalah: Trichoderma-PP1, Trichoderma-PP3, Trichoderma-AG2, Trichoderma-PYK3, Paecilomyces-PP6,Paecilomyces-PP7, Paecilomyces-AG4, Paecilomyces-PYK4, Aspergillus-PP2 dan tanpa jamur antagonis (kontrol). Hasilpenelitian menunjukkan bahwa 9 isolat jamur antagonis mampu tumbuh dan bertahan pada buah cabai. Kemampuan kesintasanyang tertinggi terdapat pada Trichoderma-PP3 dan Trichoderma-AG2 (95,83%) dan yang terendah terdapat pada Paecilomyces-PP7 (50%). Trichoderma-PP3 dan Trichoderma-AG2 merupakan isolat yang terbaik dalam menekan penyakit antraknosa yangdisebabkan oleh C. gloeosporioides.
Kata kunci: antraknosa, Aspergillus, kolonisasi, Paecilomyces, Trichoderma
PENDAHULUAN
Penyakit antraknosa merupakan salah satupenyakit penting pada tanaman cabai karena menyerangbuah yang berakibat langsung pada produksi. Penyakitini disebabkan oleh Colletotrichum capsici danColletotrichum gloeosporioides (Montri et al., 2009;
Sharma & Kulshrestha, 2015). Penyakit antraknosa sulitdikendalikan karena patogen memiliki keragamangenetik yang tinggi (Than et al., 2008).
Pengendalian penyakit pada tanaman cabaisecara umum dilakukan dengan fungisida yang biasanyadilakukan sangat insentif sekali. Hal ini dapatmenimbulkan efek negatif terhadap lingkungan. Untuk

Nurbailis et al. Kesintasan Jamur Antagonis 163
menghindari dampak negatif tersebut, diperlukanalternatif pengendalian lain yang ramah lingkungan salahsatunya pengendalian hayati menggunakan jamur yangbersifat antagonis terhadap patogen.
Beberapa jamur antagonis telah teruji efektifdalam menghambat pertumbuhan Colletotrichumpenyebab penyakit antraknosa baik secara in vitromaupun in vivo. Vasanthakumari & Shivanna (2013)melaporkan bahwa dari 138 isolat jamur yang berasaldari rizosfer dan rizoplan tanaman rumput ada 15 isolatyang mampu menghambat pertumbuhan C. capsici padacabai. Isolat tersebut juga mampu mengkolonisasi akardan rizosfer tanaman cabai. Shovan et al. (2008)melaporkan bahwa 20 isolat T. harzianum yang berasaldari rizosfer dan rizoplan kedelai mampu menghambatpertumbuhan miselium C. dematium dengan dayahambat 50,93-89,44% dengan daya hambat yang tertinggiterdapat pada T. harzianum (T3). Nurbailis et al. (2014)melaporkan bahwa dari 59 isolat jamur yang berasaldari rizosfer cabai, ada sembilan isolat yang berpotensimenghambat pertumbuhan C. gloeosporioidespenyebab penyakit antraknosa pada cabai denganmekanisme antagonisme: kompetisi, antibiosis, danhiperparasitis. Isolat tersebut terdiri atas genusTrichoderma, Paecilomyces dan Aspergillus.
Sembilan jamur antagonis tersebut mampumenghasilkan toksin yang terindikasi terdapatnya zonabening pada pengujian metode biakan ganda (Nurbailiset al., 2014). Aspergillus menghasilkan aflatoksin yangmerupakan penyebab penyakit pada manusia dan hewan(Amaike & Keller, 2011). Trichoderma menghasilkantoksin yang erat kaitannya dengan perannya sebagaiagens pengendali hayati (Woo et al., 2006; Howell, 2006;Leelavathi et al., 2014). Jamur ini terdiri dari berbagaistrain dan spesies, dapat ditemukan pada berbagaihabitat tanah dan bahan organik, mudah diisolasi dandibiakkan serta tumbuh cepat pada berbagai substrat(Well, 1986; Blaszczyk et al., 2011). Paecilomycesadalah jamur yang bersifat kosmopolit yang hidup ditanah, sisa tanaman yang telah membusuk dan produkmakanan. Menurut Kalay et al. (2008) jamurPaecilomyces lilacianus dapat mengendalikannematoda Globodera rostochiensis pada tanamankentang secara in vitro.
Salah satu faktor yang penting dicermati dalampemanfaatan jamur antagonis sebagai agenspengendalian hayati C. gloeosporioides penyebabpenyakit antraknosa pada cabai adalah kemampuankesintasan jamur antagonis pada buah cabai.Caisemsaeng et al. (2013) melaporkan bahwa ragiisolat HS6, SS11, SLD5, SS10 dan PLN13 berpotensimenghambat pertumbuhan C. capsici pada cabai. Isolat
PLN13 merupakan isolat yang tinggi keberadaanyapada buah cabai. Menurut Parey et al. (2013), beberapajamur yang berasosiasi dengan buah cabai yangmenunjukkan gejala busuk dari berbagai lokasi di Indiaantara lain: C. capsici, Fusarium pallidoroseum, F.oxysporum, F. moniliforme, Alternaria alternata danAspergillus flavus. Hasil penelitian Basha et al. (2010)menunjukkan dari 9 sampel buah cabai yang berasaldari lokasi yang berbeda di India, berhasil diisolasi 26isolat jamur eksofit dan 18 isolat jamur endofit. Isolatyang dominan didapatkan adalah A. flavus. Isolat inimerupakan yang terbaik dalam menghambatpertumbuhan C. gloeosporioides (70%) in vitro.
Penelitian ini bertujuan mendapatkan jamurantagonis unggul yang mampu tumbuh dan bertahanpada buah cabai serta efektif dalam menekan tingkatserangan penyakit antraknosa yang disebabkan oleh C.gloeosporioides pada cabai.
METODE PENELITIAN
Tempat dan Waktu. Penelitian ini berlangsung diLaboratorium Fitopatologi Jurusan Hama dan PenyakitTanaman dan Rumah kaca Fakultas PertanianUniversitas Andalas Padang dari bulan Mei sampaiDesember 2015.
Rancangan Penelitian. Penelitian terdiri atas dua unitpercobaan yaitu uji kesintasan jamur antagonis pada buahcabai dan uji potensi jamur antagonis terhadappenekanan penyakit antraknosa pada buah cabai.Rancangan yang digunakan adalah Rancangan AcakKelompok (RAK) yang terdiri dari 10 perlakuan dan 4batang tanaman cabai sebagai ulangan, setiap tanamanditetapkan 4 buah cabai untuk diberi perlakuan.Perlakuannya adalah: Trichoderma-PP1, Trichoderma-PP3, Trichoderma-AG2, Trichoderma-PYK3,Paecilomyces-PP6, Paecilomyces-PP7,Paecilomyces-AG4, Paecilomyces-PYK4,Aspergillus-PP2 dan tanpa jamur antagonis (kontrol).Data diolah menggunakan analisis ragam dan dilanjutkandengan uji Tukey pada taraf nyata 5%.
Penyediaan Bibit dan Penanaman Cabai. Bibit cabaiyang digunakan berasal dari petani cabai di daerahKuranji, Kota Padang. Bibit yang telah berumur 30 haridipindahkan ke dalam polibag yang telah diisi denganmedium tanam berupa campuran tanah dengan pupukkandang (2:1, v/v) sebanyak 10 kg setiap polibag. Polibagditempatkan di rumah kaca dan dilakukan pemupukanmenggunakan pupuk mutiara yang mengandung unsurN, P, K dengan dosis 5 g untuk setiap tanaman. Pupuk

164 J. HPT Tropika Vol. 17, No. 2, 2017: 162 - 169
buatan diberikan 2 kali, yaitu 15 dan 45 hari setelahtanam. Penyiraman dilakukan pada pagi dan sore hari,perawatan tanaman dilakukan sampai tanaman berbuah.
Perbanyakan Isolat Jamur Antagonis. Jamurantagonis yang berasal dari rizosfer cabai hasil penelitiansebelumnya, dibiakkan kembali pada Agar kentangDekstrosa (AKD). Jamur yang digunakan dalampenelitian adalah seperti pada Tabel 1.
Persiapan Jamur Patogen C. gloeosporioides. JamurC. gloeosporioides yang digunakan berasal dari koleksiLaboratorium Fitopatologi Fakultas PertanianUniversitas Andalas Padang. Kultur stok dibiakkankembali ke dalam cawan petri yang telah berisi mediumAKD. Setelah jamur tumbuh, diuji kembali virulensinyapada buah cabai dengan cara: suspensi C.gloeosporioides dengan kerapatan konidia 106 ml-1
akuades, diambil sebanyak 10 ml dan dimasukkan kedalam botol semprot. Suspensi ini kemudiandisemprotkan secara merata pada permukaan buahcabai,yang telah ditusuk sebanyak tiga titik menggunakanjarum pentul. Pengamatan dilakukan setiap hari sampai15 hari setelah inokulasi. Isolat dinyatakan virulen jikabuah cabai yang diinokulasi menunjukkan gejala dalamwaktu 15 hari.
Penyediaan Suspensi Konidia Jamur Antagonis danJamur C. gloesporioides. Suspensi konidia jamurantagonis dan patogen disiapkan dengan cara: jamurantagonis dibiakkan dalam medium AKD sampaimemenuhi cawan petri, kemudian ditambahkan 10 mlakuades steril. Konidia dilepas menggunakan kuas halus.Penghitungan konidia menggunakan Haemocytometer.Kerapatan konidia yang digunakan untuk perlakuanadalah 106 konidia ml-1. Suspensi konidia diicampurdengan Tween 80 sebanyak 0,05% sebagai perata.Perlakuan Buah Cabai. Buah cabai yang diberiperlakuan adalah buah cabai yang masih berada padabatang tanaman cabai. Buah yang telah berumur 1 bulansetelah pembungaan diaplikasikan dengan suspensikonidia jamur antagonis (106 konidia ml-1 ) sebanyak 1ml suspensi untuk setiap buah cabai. Teknik aplikasiuntuk pengamatan kesintasan adalah denganmenyemprot suspensi konidia ke seluruh permukaanbuah cabai.
Teknik aplikasi untuk pengamatan potensi jamurantagonis dalam menekan penyakit antraknosa adalahbuah cabai yang telah diperlakukan dengan jamurantagonis, 7 hari kemudian diinokulasi dengan suspensiC. gloeosporioides (106 konidia ml-1 ) sebanyak 1 ml
suspensi untuk setiap buah cabai dengan teknik yangsama dengan aplikasi jamur antagonis.
Pengamatan Kesintasan Jamur Antagonis padaBuah Cabai. Kemampuan kesintasan jamur antagonispada buah cabai ditentukan mulai 7 hari sampai 28 harisetelah aplikasi jamur antagonis dengan interval waktu7 hari. Tingkat kesintasan ditentukan dengan metodeyang dikemukakan oleh Ozbay & Steven (2004) yaitudengan membuat fragmentasi buah. Buah dipotongsepanjang 1 cm, kemudian potongan buah ditempatkandidalam cawan petri yang telah berisi media AKD, setiapcawan petri berisi 6 potong buah cabai. Buah cabaidinyatakan dikolonisasi oleh jamur antagonis apabila daripotongan buah tumbuh minimal satu koloni jamurantagonis. Kesintasan dihitung dengan rumus :
dengan:K = kesintasanA = Jumlah buah cabai yang membentuk koloni jamur antagonis pada medium AKDB = Jumlah potongan buah cabai yang diinkubasikan pada medium AKD
Penyakit Antraknosa pada Buah Cabai.Pengamatan penyakit antraknosa pada buah cabaidiamati pada percobaan unit 2 yaitu: percobaan untukmengamati potensi jamur antagonis terhadap penekananpenyakit antraknosa pada buah cabai. Pengamatandilakukan 3 hari setelah inokulasi jamur patogen, variabelyang diamati adalah:
Masa Inkubasi. Masa inkubasi diamati mulai dari harike tiga sampai 28 hari setelah inokulasi jamur patogen.Pengamatan dilakukan setiap hari dengan mengamatiadanya gejala antraknosa pada buah cabai.
Persentase Buah Terserang. Pengamatan dilakukanmulai hari ke 7 sampai 28 hari setelah inokulasi patogendengan interval waktu 7 hari. Persentase buah terserangdihitung menggunakan rumus:
dengan:P = Persentase buah terserangJBS = Jumlah buah yang bergejala antraknosaJBK = Jumlah buah yang diamati
100%JBKJBS
P
100%BA
K

Nurbailis et al. Kesintasan Jamur Antagonis 165
Keparahan Penyakit. Keparahan penyakitantraknosa ditentukan berdasarkan rumus dari Zadoks& Schein (1979):
dengan:X = Keparahan penyakitni = Jumlah buah yang terinfeksivi = Nilai numerik untuk setiap kategori seranganN = Jumlah seluruh buah yang diamatiV = Nilai skor tertinggi
Skor berdasarkan interval serangan penyakitantraknosa pada buah cabai adalah (Herwidyarti et al.,2013, modifikasi)0 = tidak ada gejala penyakit antraknosa yang diinokulasi dengan C. gloeoporioides1 = terdapat gejala penyakit antraknosa 1-<20% pada bagian buah cabai2 = terdapat gejala penyakit antraknosa 20-<40% pada bagian buah cabai3 = terdapat gejala penyakit antraknosa 41-<60% pada bagian buah cabai4 = terdapat gejala penyakit antraknosa 61-< 80% pada bagian buah cabai5 = terdapat gejala penyakit antraknosa 81-<100% pada bagian buah cabai
Data yang diperoleh kemudian dianalisis sidikragam dan dilanjutkan dengan uji Tukey pada taraf nyata5%.
100%VN
ivinX
HASIL DAN PEMBAHASAN
Kesintasan Jamur Antagonis pada Buah Cabai.Kemampuan kesintasan berbagai isolat jamur antagonispada buah cabai berbeda nyata antar perlakuan (Tabel2). Kemampuan kesintasan jamur antagonis pada buahcabai bervariasi, secara umum semua isolat mamputumbuh dan bertahan pada buah cabai.
Beberapa isolat menunjukkan kemampuan yangtinggi untuk tumbuh dan bertahan pada buah cabaiseperti Trichoderma-PP3 dan Trichoderma AG2 sertaAspergillus-PP2 yaitu di atas 90% sedangkan isolatyang mempunyai kemampuan yang rendah adalahPaecilomyces-PP7 dan Paecilomyces-AG4.Kolonisasi beberapa jamur antagonis pada buah cabaiyang telah ditumbuhkan pada medium AKD (Gambar1).
Kesintasan Jamur Antagonis pada Buah CabaiSelama 28 Hari Setelah Aplikasi. Kesintasanberbagai jamur antagonis pada buah cabai berfluktuasi,secara umum peningkatan kesintasan jamur antagonispada buah cabai terjadi pada minggu ke-3 sampai mingguke-4 setelah aplikasi. Isolat yang relatif stabilkeberadaannya pada buah cabai adalah Trichoderma-PP3 dan Trichoderma-AG2 (Gambar 2).
Trichoderma spp. termasuk jamur saprofit yangbersifat antagonis terhadap berbagai patogen tanaman,jamur ini dapat ditemui pada berbagai habitat sepertitanah dan bahan organik. Isolat Trichoderma-PP3 danTrichoderma-AG2 mempunyai kemampuan yang tinggiuntuk tumbuh dan bertahan pada permukaan buah cabaidan kesintasannya pada buah cabai lebih stabil dari isolat
Tabel 1. Jamur antagonis yang digunakan dalam penelitian
Genus jamur Kode isolat
No. Koleksi Sumber Karakter mekanisme antagonis
Trichoderma PP1 1 Rizosfer cabai Padang Panjang Kompetisi, antibiosis, parasitisme Trichoderma PP3 2 Rizosfer cabai Padang Panjang Kompetisi, antibiosis, parasitisme Trichoderma AG2 3 Rizosfer cabai Agam Kompetisi, antibiosis, parasitisme Trichoderma PYK3 4 Rizosfer cabai Payakumbuh Kompetisi, antibiosis, parasitisme Paecilomyces PP6 5 Rizosfer cabai Padang Panjang Antibiosis, parasitisme Paecilomyces PP7 6 Rizosfer cabai Padang Panjang Antibiosis, parasitisme Paecilomyces AG4 7 Rizosfer cabai Agam Antibiosis, parasitisme Paecilomyces PYK4 8 Rizosfer cabai Payakumbuh Antibiosis Aspergillus PP2 9 Rizosfer cabai Padang Panjang Antibiosis

166 J. HPT Tropika Vol. 17, No. 2, 2017: 162 - 169
Gambar 1. Kesintasan jamur antagonis pada buah cabai yang ditumbuhkan pada medium AKD (A) Trichoderma-AG2; (B) Paecilomices-PP7; (C) Aspergillus-PP2
A B C
Tabel 2. Kesintasan beberapa jamur antagonis pada buah cabai 28 hari setelah aplikasi
Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji Tukey pada taraf 5%.
Perlakuan Kemampuan kesintasan
Trichoderma-PP3 95,83 a Trichoderma-AG2 95,83 a Aspergilus-PP2 91,66 a Paecilomyces-PP6 87,50 ab Trichoderma-PP1 83,33 ab Trichoderma-PYK3 83,33 ab Paecilomyces-PYK4 83,32 ab Paecilomyces-AG4 66,67 ab Paecilomyces-PP7 50,00 b Kontrol 0,000 c
Gambar 2. Kesintasan jamur antagonis pada buah cabai selama 28 hari setelah aplikasi. MSA= minggu setelahaplikasi
Trichoderma-PP1
Trichoderma-PP3Trichoderma-AG2
Trichoderma-PYK3
Paecilomyces-PP6Paecilomyces-PP7Paecilomyces-AG4
Paecilomyces-PYK4Aspergillus-PP2
Kontrol
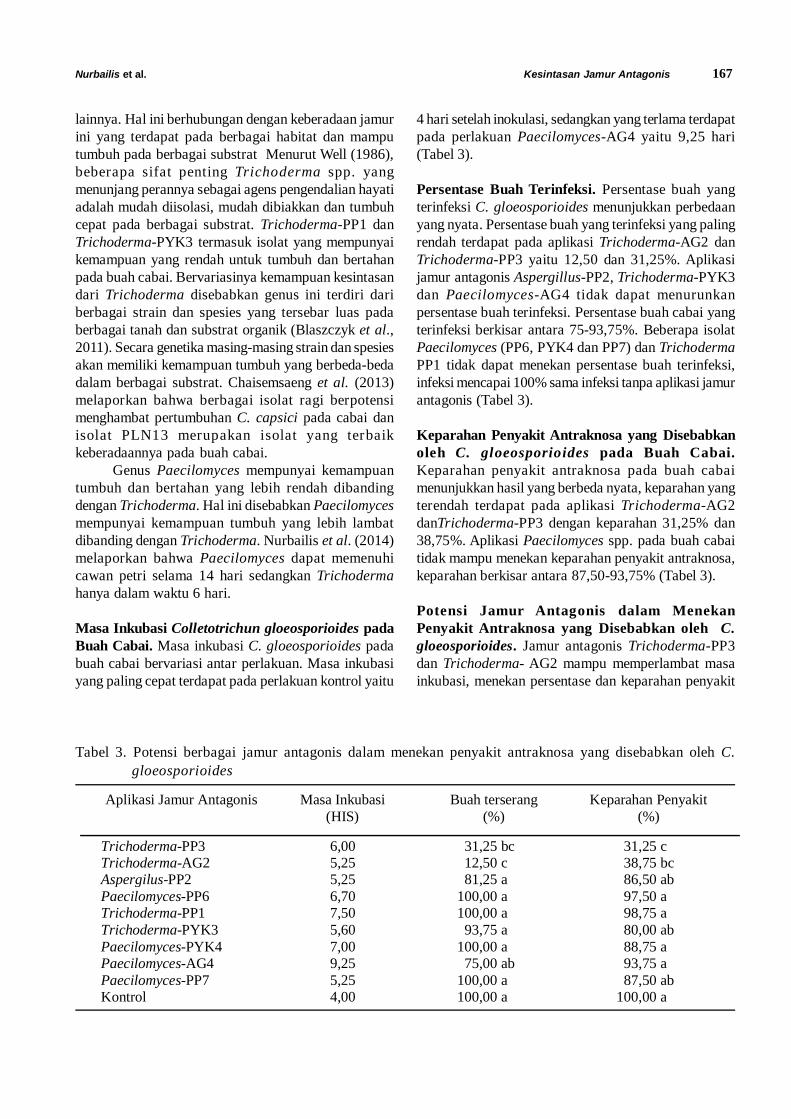
Nurbailis et al. Kesintasan Jamur Antagonis 167
lainnya. Hal ini berhubungan dengan keberadaan jamurini yang terdapat pada berbagai habitat dan mamputumbuh pada berbagai substrat Menurut Well (1986),beberapa sifat penting Trichoderma spp. yangmenunjang perannya sebagai agens pengendalian hayatiadalah mudah diisolasi, mudah dibiakkan dan tumbuhcepat pada berbagai substrat. Trichoderma-PP1 danTrichoderma-PYK3 termasuk isolat yang mempunyaikemampuan yang rendah untuk tumbuh dan bertahanpada buah cabai. Bervariasinya kemampuan kesintasandari Trichoderma disebabkan genus ini terdiri dariberbagai strain dan spesies yang tersebar luas padaberbagai tanah dan substrat organik (Blaszczyk et al.,2011). Secara genetika masing-masing strain dan spesiesakan memiliki kemampuan tumbuh yang berbeda-bedadalam berbagai substrat. Chaisemsaeng et al. (2013)melaporkan bahwa berbagai isolat ragi berpotensimenghambat pertumbuhan C. capsici pada cabai danisolat PLN13 merupakan isolat yang terbaikkeberadaannya pada buah cabai.
Genus Paecilomyces mempunyai kemampuantumbuh dan bertahan yang lebih rendah dibandingdengan Trichoderma. Hal ini disebabkan Paecilomycesmempunyai kemampuan tumbuh yang lebih lambatdibanding dengan Trichoderma. Nurbailis et al. (2014)melaporkan bahwa Paecilomyces dapat memenuhicawan petri selama 14 hari sedangkan Trichodermahanya dalam waktu 6 hari.
Masa Inkubasi Colletotrichun gloeosporioides padaBuah Cabai. Masa inkubasi C. gloeosporioides padabuah cabai bervariasi antar perlakuan. Masa inkubasiyang paling cepat terdapat pada perlakuan kontrol yaitu
4 hari setelah inokulasi, sedangkan yang terlama terdapatpada perlakuan Paecilomyces-AG4 yaitu 9,25 hari(Tabel 3).
Persentase Buah Terinfeksi. Persentase buah yangterinfeksi C. gloeosporioides menunjukkan perbedaanyang nyata. Persentase buah yang terinfeksi yang palingrendah terdapat pada aplikasi Trichoderma-AG2 danTrichoderma-PP3 yaitu 12,50 dan 31,25%. Aplikasijamur antagonis Aspergillus-PP2, Trichoderma-PYK3dan Paecilomyces-AG4 tidak dapat menurunkanpersentase buah terinfeksi. Persentase buah cabai yangterinfeksi berkisar antara 75-93,75%. Beberapa isolatPaecilomyces (PP6, PYK4 dan PP7) dan TrichodermaPP1 tidak dapat menekan persentase buah terinfeksi,infeksi mencapai 100% sama infeksi tanpa aplikasi jamurantagonis (Tabel 3).
Keparahan Penyakit Antraknosa yang Disebabkanoleh C. gloeosporioides pada Buah Cabai.Keparahan penyakit antraknosa pada buah cabaimenunjukkan hasil yang berbeda nyata, keparahan yangterendah terdapat pada aplikasi Trichoderma-AG2danTrichoderma-PP3 dengan keparahan 31,25% dan38,75%. Aplikasi Paecilomyces spp. pada buah cabaitidak mampu menekan keparahan penyakit antraknosa,keparahan berkisar antara 87,50-93,75% (Tabel 3).
Potensi Jamur Antagonis dalam MenekanPenyakit Antraknosa yang Disebabkan oleh C.gloeosporioides. Jamur antagonis Trichoderma-PP3dan Trichoderma- AG2 mampu memperlambat masainkubasi, menekan persentase dan keparahan penyakit
Tabel 3. Potensi berbagai jamur antagonis dalam menekan penyakit antraknosa yang disebabkan oleh C.gloeosporioides
Aplikasi Jamur Antagonis
Masa Inkubasi
(HIS)
Buah terserang
(%)
Keparahan Penyakit
(%)
Trichoderma-PP3 6,00 31,25 bc 31,25 c Trichoderma-AG2 5,25 12,50 c 38,75 bc Aspergilus-PP2 5,25 81,25 a 86,50 ab Paecilomyces-PP6 6,70 100,00 a 97,50 a Trichoderma-PP1 7,50 100,00 a 98,75 a Trichoderma-PYK3 5,60 93,75 a 80,00 ab Paecilomyces-PYK4 7,00 100,00 a 88,75 a Paecilomyces-AG4 9,25 75,00 ab 93,75 a Paecilomyces-PP7 5,25 100,00 a 87,50 ab Kontrol 4,00 100,00 a 100,00 a

168 J. HPT Tropika Vol. 17, No. 2, 2017: 162 - 169
antraknosa. Isolat Aspergillus-PYK4, Paecilomycesspp (PP6, PP7, AG4 dan PYK4), Trichoderma-OPP1dan Trichoderma-PKY3 tidak mampu menekan tingkatserangan penyakit antraknosa pada cabai denganpersentase serangan di atas 75% dan keparahanpenyakit di atas 80%.
Isolat Trichoderma-PP3 dan Trichoderma-AG2mampu menekan perkembangan penyakit antraknosapada cabai. Propagul isolat Trichoderma yang beradapada buah cabai mampu menekan pertumbuhan C.gloeosporioides dengan berbagai mekanisme sepertipersaingan, mengeluarkan senyawa metabolit yangdapat menghambat pertumbuhan C. gloesporioidesataupun bersifat hiperparasit. Nurbailis et al. (2014)melaporkan bahwa Trichoderma spp. bersifat parasitterhadap miselium C. gloeosporoides. Menurut Wooet al. (2006), Trichoderma spp. merupakan jamurantagonis dan penghasil antibiotika yang kuat. Gliotoksinmerupakan antibiotika yang dihasilkan olehTrichoderma yang erat kaitannya dengan perannyasebagai agens pengendalian hayati patogen tanaman(Howell, 2006).
Isolat Aspergillus-PP2 pada buah cabai ternyatahanya mampu memperlambat masa inkubasi sedangkanpersentase dan keparahan penyakit antraknosa masihcukup tinggi yaitu 81,25% dan 86,50% walaupun tingkatkesintasan pada buah cabai cukup tinggi yaitu 91,67%(Tabel 2). Hal ini disebabkan karena Aspergillus tidakmampu menghambat pertumbuhan C. gloeosporioidesbaik dalam berkompetisi, antibiosis ataupun hiperparasit.Secara umum Aspergillus diketahui menghasilkanaflatoksin, ternyata toksin ini tidak mampu menghambatpertumbuhan C. gloeosporioides in vivo. MenurutAmaike & Keller (2011), A. flavus menghasilkanmikotoksin yang dikenal dengan aflatoksin yang toksikterhadap manusia dan hewan. Nurbailis et al. (2014)melaporkan bahwa Aspergillus tidak bersifat parasitterhadap C. gloeosporioides penyebab penyakitantraknosa pada buah cabai.
Kesintasan isolat Paecilomyces spp. pada buahcabai ternyata tidak mampu menekan persentase dankeparahan penyakit antraknosa. Pada pengujian in vitrodapat dideteksi bahwa jamur ini membentuk zona beningpada pertemuan koloni dengan C. gloeosporioides padapengujian biakan ganda (Nurbailis et al., 2014). Hal inimengindikasikan bahwa Paecilomyces menghasilkansuatu senyawa yang dapat menghambat pertumbuhanjamur patogen, ternyata senyawa ini tidak mampumenghambat pertumbuhan C. gloeosporioides in vivo.Penelitian Kalay et al. (2008) menunjukkan bahwa jamurPaecilomyces lilacianus dapat mengendalikan
nematoda Globodera rostochiensis pada tanamankentang in vitro.
SIMPULAN
Sembilan isolat jamur antagonis mampu tumbuhdan bertahan pada buah cabai. Kemampuan kesintasantertinggi terdapat pada Trichoderma-PP3 danTrichoderma-AG2 (95,83%) dan terendah terdapatpada Paecilomyces-PP7 (50%). Trichoderma-PP3 danTrichoderma-AG2 merupakan isolat yang terbaikdalam menekan penyakit antraknosa yang disebabkanoleh C. gloeosporioides.
SANWACANA
Penulis menyampaikan penghargaan danterimakasih kepada Direktorat Jenderal PendidikanTinggi Kementerian Riset, Teknologi dan PendidikanTinggi atas bantuan dana penelitian melalui skim HibahBersaing tahun 2015.
DAFTAR PUSTAKA
Amaike S & Keller NP. 2011. Aspergillus flavus. Annu.Rev. Phytopathol. 49: 107–133.
Basha H, Hemannavar V, Ramanujam B,Rangeshwaran R, & Sriram S. 2010. Screeningof chilli microflora and other biocontrol agentsfor their antagonistic effects on Colletotrichumspp. infecting chillies. J. Plant Prot. Sci. 2(1):38–44.
Blaszczyk L, Popie D, Chelkowski J, Koczyk G, SamuelsGJ, Sobieralski K, & Siwulski M. 2011. Speciesdiversity of Trichoderma in Poland. J. Appl.Genet. 52(2): 233–243.
Chaisemsaeng P, Mongkolthanaruk W, &Bunyatratchata W. 2013. Screening and potentialfor biological control of anthracnose disease(Colletotrichum capsici) on chili fruits by yeastisolates. J. Life Sci. Tech. 1(4): 202–204.
Herwidyarti KH, Ratih S, & Sembodo DRJ. 2013.Keparahan penyakit antraknosa pada cabai(Capsicum annuum) dan berbagai jenis gulma.J. Agrotek Tropika (1)1: 102–106.
Howell CR. 2006. Understanding the mechanismsemployed by Trichoderma virens to effectbiological control of cotton diseases.Phytopathology 96(2): 178–180.

Nurbailis et al. Kesintasan Jamur Antagonis 169
Kalay AM, Natasasmita S, Suganda T, & Simarmata T.2008. Uji parasitik beberapa spesies jamur tanahterhadap Globodera rostochiensis (Woll.) secarain vitro. J. Natur Indonesia (10)2: 73–75.
Leelavathi MS, Vani L, & Reena P. 2014. Antimicrobialactivity of Trichoderma harzianum againstbacteria and fungi. Int. J. Curr.Microbiol.App.Sci. 3(1): 96–103.
Montri P, Taylor PWJ, & Mongkolporn O. 2009.Pathotypes of Colletotrichum capsici, the causalagent of chili anthracnose, in Thailand. Plant Dis.93(1): 17–20.
Nurbailis, Martinius, & Azniza V. 2014.Keanekaragaman jamur pada rizosfer tanamancabai sistem konvensional dan organik danpotensinya sebagai agen pengendali hayatiColletotrichum gloeosporioides . J. HPTTropika 14(1): 16–24.
Ozbay N & Newman SE. 2004. Biological control withTrichoderma spp. with emphasis an T.harzianum. Pak. J. Biol. Sci. 4(7): 478–484.
Parey MA, Razdan VK, & Sofi TA. 2013. Comparativestudy of different fungi associated with fruit rotof chilli and screening of chilli germplasm againstColletotrichum capsici. Int. J. Agri. Crop Sci.5(7): 723–730.
Sharma M & Kulshrestha S. 2015. Colletotrichumgloeosporioides: an antracnose causing pathogenof fruits and vegetables. Biosci. Biotech. Res.Asia 12(2): 1233–1246.
Shovan LR, Bhuiyan MKA, Begum JA, & Pervez Z.2008. In vitro control of Colletotrichumdematium causing antracnose of soybean byfungicide, plant extract and Trichodermaharzianum. Int. J. Sustain. Crop. Prod. 3(3):10–17.
Than PP, Prihastuti H, Phoulivong S, Taylor PWJ, &Hyde KD. 2008. Chilli anthracnose diseasecaused by Colletotrichum species. J. ZhejiangUniv. Sci. B. 9(10): 764–778.
Vasanthakumari MM & Shivanna MB. 2013. Biologicalcontrol of anthracnose of chilli with rhizosphereand rhizoplane fungal isolates from grasses. Arch.Phytopathol. Plant Prot. 46(14): 1641–1666.
Well HD. 1986. Trichoderma as Biocontrol Agent. In:Mukerji KG & Garg KL (Eds.). Biocontrol ofPlant Disease. Vol. II. pp.72–82. CRC PressInc., Boca Raton, Florida.
Woo SL, Scala F, Ruocco M, & Lorito M. 2006. Themolecular biology of the interactions betweenTrichoderma spp., phytopathogenic fungi andplants. Phytopathology 96(2): 181–185.
Zadoks JC & Schein, RD. 1979. Epidemiology andPlant Disease Management. Oxford UniversityPress, New York.

Kusuma et al. Karekterisasi Morfologi 147
KARAKTERISASI MOLEKULER NUCLEOPOLYHEDROVIRUS (NPV)HYPOSIDRA TALACA WLK. DI PERKEBUNAN TEH
GUNUNG MAS BOGOR
R. Yayi Munara Kusumah, Lestia Revi, & Fitrianingrum Kurniawati
Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian BogorJl. Kamper Kampus IPB Dramaga, Bogor 16680
E-mail: [email protected]
ABSTRACT
Characterize molecular the Nucleopolyhedrovirus (NPV) Hyposidra talaca Wlk. of tea plantation at Gunung Mas Bogor.Hyposidra talaca is one of the most important pest in tea plantation, and generally attacks of leaves and shoots. This pestcause yield loss up to 40-100%. NPV can be pathogenic to the pest of H. talaca and can be developed as an alternativemeasure to control H. talaca in tea plantations and based management appears to be more ecofriendly and effective. However,information regarding characterisation molecular of HytaNPV is limited. The study conducted to characterize molecular theNPV of H. talaca by restriction nucleotide and amino acid, by using gens lef-8. Molecular identification used PolymeraseChain Reaction (PCR) consisted of DNA extraction, DNA amplication, and DNA electrophoresis. DNA amplication using genlef-8 showed positif result with approximately 770 bp. Gen lef-8 can identified HytaNPV. DNA sequance showed that isolateHytaNPV Bogor had high homology of pathogonic NPV of genus Helicoverpa from Brazil, Australia, Spanyol and Netherlandwith homology nucleotide and amino acid reached 98% and 100%. Based on philogeny tree of HytaNPV was one group withpathogenic NPV of genus Helicoverpa.
Key words: characterize NPV, gens lef-8, Hyposidra talaca, NPV
ABSTRAK
Karakterisasi molekuler nucleopolyhedrovirus (NPV) Hyposidra talaca wlk. di perkebunan teh Gunung Mas Bogor. LarvaHyposidra talaca termasuk hama penting pada tanaman teh yang menyerang daun yang masih muda atau pucuk daun, hamaini dapat menyebabkan kerugian mencapai 40-100% jika tidak dilakukan pengendalian. Salah satu teknik pengendalian H.talaca yang sejalan dengan prinsip PHT yaitu memanfaatkan agen hayati seperti nucleopolyhedrovirus (NPV) yang memilikiinang spesifik, tidak membahayakan lingkungan, dapat mengatasi masalah keresistansian hama terhadap insektisida, danselaras dengan komponen PHT lainnya. Namun, informasi mengenai karakterisasi molekuler HytaNPV masih sangat terbatas.Tujuan penelitian ini adalah mengetahui karakteristik molekuler melalui urutan nukleotida dan asam amino isolat NPV yangberasal dari H. talaca dengan menggunakan gen lef-8. Identifikasi molekuler dengan PCR terdiri beberapa tahap yaitu ekstraksi,amplifikasi dan elektroforesis DNA. Amplifikasi DNA HytaNPV menggunakan primer lef-8 berhasil mengisolasi gen lef-8HytaNPV dengan ukuran fragmen DNA sekitar 770 bp. Analisis urutan DNA berdasarkan gen lef-8 menunjukkan bahwa isolatHytaNPV Bogor tergolong ke dalam spesies virus yang memiliki hubungan kekerabatan yang dekat dengan isolat NPV yangmenyerang genus Helicoverpa yang berasal dari Brazil, Australia, Spanyol dan Belanda dengan nilai homologi nukleotida danasam amino sebesar 98% dan 100%. Berdasarkan analisis filogeni, isolat HytaNPV masuk dalam grup yang sama dengan NPVyang menyerang genus Helicoverpa.
Kata kunci: DNA, gen lef-8, Hyposidra talaca, karakterisitik molekuler, NPV
PENDAHULUAN
Teh (Camelia sinensis L.) merupakan tanamanperkebunan penting di Indonesia dan menjadi salah satupenyumbang devisa negara yang cukup besar. BPS(2016) melaporkan bahwa produksi teh di Indonesiahanya mencapai sekitar 145.86 kg teh kering per hektarper tahun pada tahun 2013. Sedangkan, produksi tehtahun 2014 yaitu sekitar 142.72 kg per hektar per tahun.
Angka produksi tersebut masih tergolong rendahdibandingkan dengan produksi negara penghasil tehlainnya. Turunnya produksi sangat mungkin disebabkanoleh turunnya produktivitas tanaman. Menurut Widayat(1996), rendahnya produktivitas teh di Indonesia lebihbanyak disebabkan oleh serangan organismepengganggu tanaman (OPT) khususnya hama. Hamayang biasa menyerang tanaman teh adalah kepik daunteh (Helopeltis antonii), wereng pucuk teh (Empoasca
J. HPT Tropika. ISSN 1411-7525Vol. 17, No. 2: 105 – 110, September 2017
J. HPT Tropika. ISSN 1411-7525Vol. 17, No. 2: 147 – 155, September 2017

148 J. HPT Tropika Vol. 17, No. 2, 2017: 147 - 155
sp.), ulat penggulung daun (Homona coffearia (L.)Nietn), ulat penggulung pucuk (Cydia leucostoma (L.)Meyr), dan ulat jengkal (Hyposidra talaca Wlk).
H. talaca merupakan hama penting pada tanamanteh (Ghosh et al., 2015). H. talaca umumnya menyerangdaun yang masih muda atau pucuk daun. Serangan beratdapat menyebabkan daun berlubang dan pucuk tanamangundul. Perkebunan teh milik PT. PerkebunanNusantara VIII Gunung Mas, Bogor sering menemuipermasalahan yang cukup serius akibat serangan hamaH. talaca (Pradana, 2013). Kehilangan hasil akibatserangan H. talaca cukup tinggi, pada musim kemaraularva H. talaca dapat mengakibatkan kerugian mencapai40-100% apabila tidak dilakukan pengendalian (Mulianiet al., 2011).
Pengendalian hama yang tepat dan bijaksanadapat mencegah kehilangan hasil dan menjaga kualitasserta keamanan produk. Undang-undang RI No. 12tahun 1992 mencanangkan pelaksanaan pengendalianhama terpadu (PHT) yang melarang penggunaan saranadan cara yang dapat mengganggu kesehatan manusiadan mencemari lingkungan (Widayat et al., 1996; Kunimi,2007).
Nucleopolyhedrovirus (NPV) merupakan salahsatu patogen serangga yang efektif sebagai agenpengendali serangga hama. NPV telah diuji efektif untukmengendalikan ulat grayak dan ulat pemakan polong(Pramono, 2000; Arifin, 2006). NPV juga efektif untukmengendalikan ulat jengkal (H. talaca) dan disebut jugaHytaNPV dengan persentase penurunan populasi hama50-100% dalam waktu 7-11 hari (Pradana, 2013).HytaNPV telah dilaporkan patogenik terhadap hama dilaboratorium (Mukhopadhyay et al., 2011). NPVmemiliki inang spesifik, tidak membahayakan lingkungan,dan sesuai dengan komponen PHT lainnya (Starnes etal., 1993).
NPV merupakan virus dari famili Baculoviridaeyang mampu menginfeksi lebih dari 400 spesiesserangga. Baculoviridae merupakan famili virus yangmampu menginfeksi serangga khususnya ordoLepidoptera (Blissard et al., 2000). Informasi mengenaisekuens nukeleotida HytaNPV asal Indonesia saat inibelum ada di situs GenBank. Penelitian ini bertujuanuntuk mengetahui karakteristik molekuler melalui urutannukleotida isolat NPV yang berasal dari H. talaca diPerkebunan teh Gunung Mas Bogor dengan gen lef-8.
METODE PENELITIAN
Tempat dan Waktu. Sampel isolat NPV diperoleh dariperkebunan teh Gunung Mas Jawa Barat. Perlakuan danpengolahan data dilakukan di Laboratorium Patologi
Serangga. Karakterisasi molekuler dilakukan diLaboratorium Virologi Tumbuhan, Departemen ProteksiTanaman, Fakultas Pertanian, Institut Pertanian Bogor.Penelitian dilaksanakan dari bulan Februari sampai Mei2016.
Isolasi NPV. Sampel cadaver ulat H. talaca yangmenunjukkan gejala terserang NPV diambil dariperkebunan teh Gunung Mas. Cadaver digerus denganmortar dalam larutan sodium deodecyl sulphate (SDS)0,1%, filtrat dituangkan ke dalam tabung mikro 2 ml,kemudian disentrifugasi dengan microsentrifuge SorvalBiofuge (Fresco) dengan kecepatan 2000 rpm selama 1menit pada suhu 4 °C. Pelet yang terbentuk dibuang,supernatan diambil dan disentrifugasi kembali dengankecepatan 5000 rpm selama 20 menit. Pelet yangterbentuk ditambahkan beberapa tetes aquabides (Cheng,1998).
Pemurnian NPV terhadap H. talaca. Pemurnian virusdilakukan dengan metode gradien sukrosa yang dibuatsecara kontinyu pada tabung mikro 2 ml (Grzywacz etal., 2011). Suspensi polihedra pada proses sebelumnyadi sentrifugasi dalam gradien sukrosa dengan kecepatan13.000 rpm selama 99 menit. Pita putih yang terbentukdiambil dengan menggunakan pipet pasteur kemudianditambahkan beberapa tetes aquabides. Suspensi tersebutdisentrifugasi kembali pada 7000 rpm selama 20 menit.Endapan yang terbentuk diambil dan diresuspensi denganaquabides. Polihedra hasil purifikasi diamati di bawahmikroskop cahaya.
Pemurnian Virion dan Ekstraksi DNA. Hasilpurifikasi sebelumnya diresuspensikan dengan larutansodium karbonat (Na2CO3, pH 11), dan dikocok pelanpada suhu 37 oC selama 30 menit, kemudiandisentrifugasi kembali menggunakan tabung mikro 2 mlpada 13.000 rpm selama 1 jam. Suspensi viriondimurnikan dengan metode yang sama pada pemurnianpolihedra (Cheng, 1998), selanjutnya dilakukan ekstraksiDNA dengan menggunakan Viral nucleic acid extractionkit II (Geneaid).
Proses ekstraksi DNA genom virus meliputibeberapa tahapan yaitu pertama tahap lisis yaitu Sebanyak200 µl virion murni dimasukkan ke dalam tabung mikrokemudian 400 µl VB lysis buffer ditambahkan. Suspensitersebut dikocok menggunakan vortex kemudiandiinkubasi pada suhu ruangan selama 10 menit.Selanjutnya tahap pengikatan asam nukleat, padasuspensi ditambahkan 450 µl AD buffer, kemudiandipindahkan ke tabung mikro yang di dalamnya terdapattabung kolom VB, kemudian disentrifugasi pada 13.000

Kusuma et al. Karekterisasi Morfologi 149
rpm selama 1 menit. Tahap pencucian: sebanyak 400 µlW1 Buffer ditambahkan pada tabung kolom VB,kemudian disentrifugasi pada kecepatan 13.000 rpmselama 1 menit. Pada tabung kolom VB ditambahkan600 µl wash buffer, kemudian disentrifugasi kembalipada 13.000 rpm selama 1 menit. Cairan yangtertampung pada dasar tabung mikro dibuang, kemudiandisentrifugasi kembali pada 13.000 rpm selama 3 menithingga dasar tabung kolom VB kering. Tahap terakhiryaitu elusi asam nukleat, tabung kolom VB dipindahkanke dalam tabung mikro yang baru, kemudianditambahkan 50 µl RNAase free water, dan didiamkanselama 3 menit. Setelah itu disentrifugasi padakecepatan 13.000 rpm selama 1 menit. Cairan akhir yangtertampung di dasar tabung mikro merupakan DNA hasilekstraksi. DNA kemudian di nano drop untuk melihatkonsentrasi DNA hasil ekstraksi.
Amplifikasi DNA NPV Menggunakan PCR .Amplifikasi DNA menggunakan primer Gen lef-8dengan urutan oligonukleotida primer forward 5’-ATGAATTGCAAACTCTCCGCCCAG-3’ danreverse 3’-TCGACTGCAGACC-GCCGAAGA-5’(Kaur et al., 2014). Untuk amplifikasi isolat HytaNPV,sebanyak 2 µl DNA hasil ekstraksi dicampur denganreagen yang terdiri atas 12,5 µl Taq DNA polymerase,8 µl ddH2O, 1 µl primer forward, 1 µl primer reverse,0,5 µl MgCl hingga volume akhir yang digunakan dalamamplifikasi sebanyak 25 µl untuk satu reaksi. Amplifikasidilakukan pada mesin PCR (gene amp system 9700).Satu siklus amplifikasi meliputi tiga tahapan utama yaitu:pradenaturasi 94 oC selama 5 menit, denaturasi 94 oCselama 1 menit, penempelan 61 oC selama 1 menit,ekstensi 72oC selama 1 menit, dan pasca ekstensi 72 oCselama 7 menit, dengan suhu akhir penyimpanan 4 oC.Siklus diulangi sebanyak 35 kali. Proses elektroforesisdilakukan dengan memasukan 5 µl DNA hasil amplifikasike dalam sumur gel agarose 1% yang direndam dalam
larutan Tris Borate EDTA (TBE) kemudian dialiri denganarus listrik 50 volt selama 50 menit. Selanjutnya gelagarosa 1% direndam dalam larutan Ethidium Bromida(EtBr) selama 30 menit. Visualisasi DNA dilakukandengan cara meletakkan gel agarose pada UV-transiluminator kemudian difoto dengan menggunakankamera.
Analisis Pengurutan DNA dan Pengurutan AsamAmino. DNA hasil amplifikasi dilakukan pengurutanfragmennya oleh 1st BASE Singapura melalui PT.Genetica Science. Urutan nukleotida sampeldibandingkan dengan urutan nukleotida NPV lain yangtelah dipublikasikan di situs National Centre forBiotechnology Information (NCBI) melalui programBLAST (Basic Local Alignment Search Tools). Dataurutan nukleotida yang terpilih dianalisis menggunakanprogram penjajaran dan ClustalW serta program Bioeditver 7.1.7 untuk mengetahui homologi nukleotida danprotein tiap sampel. Data asam amino dianalisismenggunakan situs www.expasy.com untukmenerjemahkan nukleotida menjadi asam amino. Dataurutan asam amino dianalisis menggunakan programBioedit ver 7.1.7.
Analisis Filogeni HytaNPV. Analisis filogeni dilakukanmenggunakan pendekatan ‘Neighbor-Joining’ denganBootstrap 1000x dengan program MEGA 6 (Tamuraet al., 2011).
HASIL DAN PEMBAHASAN
Pengambilan Sampel Hyposidra talaca yangTerserang NPV di Lapangan. Larva H. talaca yangmenunjukkan gejala terinfeksi NPV di perkebunan tehdiambil sebanyak-banyaknya. Larva yang terinfeksisering ditemukan menggantung dengan kedua tungkaisemu. Biasanya larva yang terinfeksi berwarna coklat
Gambar 1 Larva H. talaca yang sehat (A), Larva H. talaca yang terserang NPV (B)
B
A

150 J. HPT Tropika Vol. 17, No. 2, 2017: 147 - 155
kehitaman, jika bagian integumen pecah akanmengeluarkan lendir berwarna kecoklatan (Gambar 1).
Hasil Pemurian Virion NPV dan Nano Drop. Hasilsentrifugasi membentuk awan putih yang berkumpul dibagian tengah tabung yang merupakan kumpulan virionmurni HytaNPV. Hasil nano drop virion murnimenunjukkan bahwa konsentrasi DNA virion yaitusebesar 50%. Hasil tersebut dapat digunakan untukbahan perbanyakan DNA dengan menggunakan metodePCR.
Pengamatan Polihedra HytaNPV. PolihedraHytaNPV diamati menggunakan mikroskop cahayaperbesaran 400x berbentuk seperti butiran-butiran kecil(Gambar 2). Ukuran dan bentuk polihedra NPVbervariasi tergantung spesiesnya (Cheng, 1998). Adapunbentuk polihedra yang pernah dilaporkan yaitu kuboid,dodecahedral, tetrahedral dan bentuk tidak beraturan.Pada umumnya polihedra HytaNPV berbentuktetrahedral dengan diameter 0,476-0,274 (µm –SE) danmemiliki tipe multinukleotida (Lestari, 2008).
Karakterisasi Molekuler. Pengamatan karaktermolekuler dengan amplifikasi DNA HytaNPVmenggunakan primer forward dan reverse gen lef-8menunjukkan hasil yang baik. Hal itu dilihat dari
terbentuknya pita DNA sesuai dengan ukuran target yangdiinginkan yaitu sekitar 770 pb (pasang basa) (Gambar3).
Metode PCR (Polymerase Chain Reaction) sangatsensitif untuk mengklasifikasikan Baculovirus. Gen lef-8 merupakan salah satu gen transkripsi dalam genomBaculovirus yang merupakan gen ekspresi pada RNApolymerase yang cukup stabil mengkode subunit DNAdan menempelkan langsung pada RNA polymerase virussecara spesifik (Herniou et al., 2003). Gen late expressionfactor-8 (lef-8) merupakan gen yang banyak digunakanuntuk menganalisis DNA NPV Lepidoptera (Acharya &Gopinathan, 2002). Yu et al. (2013) telahmengarakterisasi gen late expression factor-10 (lef-10)dari Bombyx morii nucleopolyhedral virus.
Pengurutan Fragmen DNA HytaNPV. Fragmennukleotida HytaNPV Bogor dikirim dalam bentukkromatogram. Kromatogram HytaNPV menunjukkanhasil yang baik dengan tidak ada pengusutan yang terjadipada kromatogram. Hasil fragmen berisi nukleotidaforward dan reverse dicontig menggunakan softwarebioedit untuk mendapatkan sekuens lengkap dariHytaNPV.
Pengurutan DNA HytaNPV Bogor dilakukanuntuk menentukan persentase kemiripan berdasarkangen lef-8. Hasil BLAST diperoleh beberapa spesies NPV
Gambar 2. Polihedra HytaNPV dibawah mikroskop cahaya (400x)
Gambar 3. Hasil visualisasi DNA HytaNPV Bogor menggunakan primer gen lef-8 Marker (M), HytaNPV (1)
750 bp500 bp
250 bp
770 bp

Kusuma et al. Karekterisasi Morfologi 151
yang memiliki kemiripan yang cukup tinggi dengan isolatHytaNPV (Tabel 1). Parameter yang diamati pada hasilBLAST yaitu Query discovery yang berfungsi untukmengetahui persen dari total panjang dari urutannukleotida sampel yang cukup baik untuk disejajarkandengan urutan nukleotida yang dimiliki oleh GenBankNCBI dan Max/total score berfungsi untuk mengetahuipersen kesamaan antara urutan nukleotida sampel yangdisejajarkan dengan urutan nukleotida bank data(Agustin, 2013). Dua spesies dikatakan identik jikamemiliki persentase kemiripan >73% (Li et al., 2009).Besarnya tingkat keragaman di dalam suatu spesiesbergantung pada jumlah individu, penyebaran wilayahgeografis, dan sistem genetiknya (Elrod & Stansfield,2007). Isolat HytaNPV Bogor memiliki nilai kemiripantinggi dengan NPV yang menyerang genus Helicoverpadengan persentase kemiripan mencapai 99%. Nilaitersebut membuktikan bahwa sampel teridentifikasisebagai NPV.
Hasil BLAST berdasarkan gen lef-8 pada Tabel1 menunjukkan bahwa isolat HytaNPV Bogor memilikinilai max/total score yang tinggi dengan NPV yangmenyerang genus Helicoverpa yaitu berkisar 1362sampai 1380, untuk persentase kemiripan dan querydiscovery mencapai 99%. Nilai max/total score padaNPV yang menyerang genus lain seperti dari genusAgrotis, Spodoptera, Bombyx, dan Mamesta memilikinilai max/total score < 500, dengan persentasekemiripan dan query coverage sebesar < 76% dan98%. Hal ini menunjukkan semakin tinggi nilai max/total
score pada NPV yang menyerang suatu genus makapersentase kemiripannya semakin tinggi dengan sampelHytaNPV Bogor.
Homologi Nukleotida dan Asam Amino HytaNPV.Karakterisasi molekuler dari susunan dan organisasi genberfungsi menunjukkan hubungan evolusi diantaraanggota Baculoviridae. Sekuen polihedrin baiknukleotida maupun asam amino digunakan untukmengidentifikasi kekerabatan yang ditunjukkan olehprotein. Isolat HytaNPV dari Bogor memiliki nilaihomologi nukleotida dan nilai homologi asam amino yangberbeda dengan 12 NPV yang menyerang genus lainyang terdapat di Genbank (Tabel 2 dan 3). Nilaihomologi nukleotida dan asam amino tertinggi terdapatpada NPV yang menyerang genus Helicoverpa yaitusebesar 98% dan 100%.
Berdasarkan Tabel 2, homologi nukleotida isolatHytaNPV asal Bogor Jawa Barat menunjukkankekerabatan tinggi, yaitu sebesar 98% dengan homologinukleotida pada isolat HearNPV Australia (No. akses:JN584482.1), HearNPV Spanyol (No. akses:KJ701033.1), HeSNPV Australia (No. akses:KU738899.1), HezeNPV Brazil (No. akses:KM596835), HezeNPV Australia (No. akses:U67265.1),dan HezeNPV Belanda (No. akses: AF334030.1).
Selain keenam spesies Helicoverpa NPV,digunakan juga spesies pembanding dari NPV yangmenyerang spesies lainnya dari ordo Lepidoptera.Tujuannya yaitu untuk melihat jarak kekerabatan antara
Tabel 1. Hasil BLAST gen lef-8 genom penuh (www.ncbi.nlm.nih.gov)
Asal Kode akses Panjang
DNA (pb)
Persentase kemiripan
(%)
Query coverage
Max/total score
H. armigera NPV Australia JN584482.1 130992 99 99 1362/1362 H. armigera NPV Spanyol KJ701033.1 132265 99 99 1389/1389 H.SNPV Australia KU738899.1 130435 99 99 1380/1380 H. zea sNPV Brazil KM596835.1 129694 99 99 1380/1380 H. zea NPV Australia U67265.1 6517 99 99 1380/1380 H. zea NPV Belanda AF334030.1 130869 99 99 1380/1380 A. segetum NPV Poland DQ123842.1 147544 75 97 500/500 S. exigua NPV Belanda AF169823.1 135611 72 97 412/412 B. mori NPV Japan AB009987.1 3682 72 97 407/407 M. configurata NPV Canada U59461.2 155060 72 96 421/421 P. xylostella MNPV USA DQ457003.1 134417 71 97 389/389 S. litura NPV Cina AF325155 139342 70 98 345/345

152 J. HPT Tropika Vol. 17, No. 2, 2017: 147 - 155
Tabel 3. Persentase homologi asam amino HytaNPV Bogor, Jawa Barat, gen lef-8 dengan sampel dari GenBank
Asal Isolat
Persentase homologi (%)
1 2 3 4 5 6 7 8 9 10 11 12 13
1 ID 2 100 ID
3 100 100 ID 4 100 100 100 ID 5 100 100 100 100 ID 6 100 100 100 100 100 ID 7 100 100 100 100 100 100 ID 8 78.6 78.6 78.6 78.6 78.6 78.6 78.6 ID 9 77.4 77.4 77.4 77.4 77.4 77.4 77.4 86.6 ID
10 69.4 69.4 69.4 69.4 69.4 69.4 69.4 68.3 69.8 ID 11 76.2 76.2 76.2 76.2 76.2 76.2 76.2 83.7 80.3 63.3 ID 12 69 69 69 69 69 69 69 67.9 69.4 98.8 63 ID 13 67.6 67.6 67.6 67.6 67.6 67.6 67.6 66.5 67.6 66.1 61.2 66.1 ID
1. HytaNPV Bogor, 2. HearNPV Australia, 3. HearNPV Spanyol, 4.HeSNPV Australia, 5. HezeNPV Brazil, 6.HezeNPV Australia, 7. HezeNPV Belanda, 8. AgseNPV Poland, 9.SpexNPV Belanda, 10. BomoNPV Japan, 11.MacoNPV Canada, 12. PlxyMNPV USA, 13. SpliNPV Cina.
Tabel 2. Persentase homologi nukleotida HytaNPV Bogor, Jawa Barat, gen lef-8 dengan sampel dari GenBank
Asal Isolat
Persentase homologi (%)
1 2 3 4 5 6 7 8 9 10 11 12 13
1 ID
2 98.6 ID 3 98.6 100 ID 4 98.8 99.7 99.7 ID 5 98.6 100 100 99.7 ID 6 98.6 100 100 99.7 100 ID 7 98.8 99.7 99.7 100 99.7 99.7 ID 8 73 73.5 73.5 73.5 73.5 73.5 73.5 ID 9 70.5 70.6 70.6 70.7 70.6 70.6 70.7 79.4 ID
10 70.2 70.7 70.7 70.6 70.7 70.7 70.6 67.5 68 ID 11 71.5 71.4 71.4 71.5 71.4 71.4 71.5 75.8 75.5 66.5 ID 12 0.355 35.2 35.2 35.2 35.2 35.2 35.2 35 35.6 35.5 34.2 ID 13 0.363 35.9 35.9 35.9 35.9 35.9 35.9 34.7 35.3 35.2 33.7 67.8 ID
1. HytaNPV Bogor, 2. HearNPV Australia, 3. HearNPV Spanyol, 4.HeSNPV Australia, 5. HezeNPV Brazil, 6.HezeNPV Australia, 7. HezeNPV Belanda, 8. AgseNPV Poland, 9. SpexNPV Belandam, 10. BomoNPV Japan,11. MacoNPV Canada, 12. PlxyMNPV USA, 13. SpliNPV Cina.
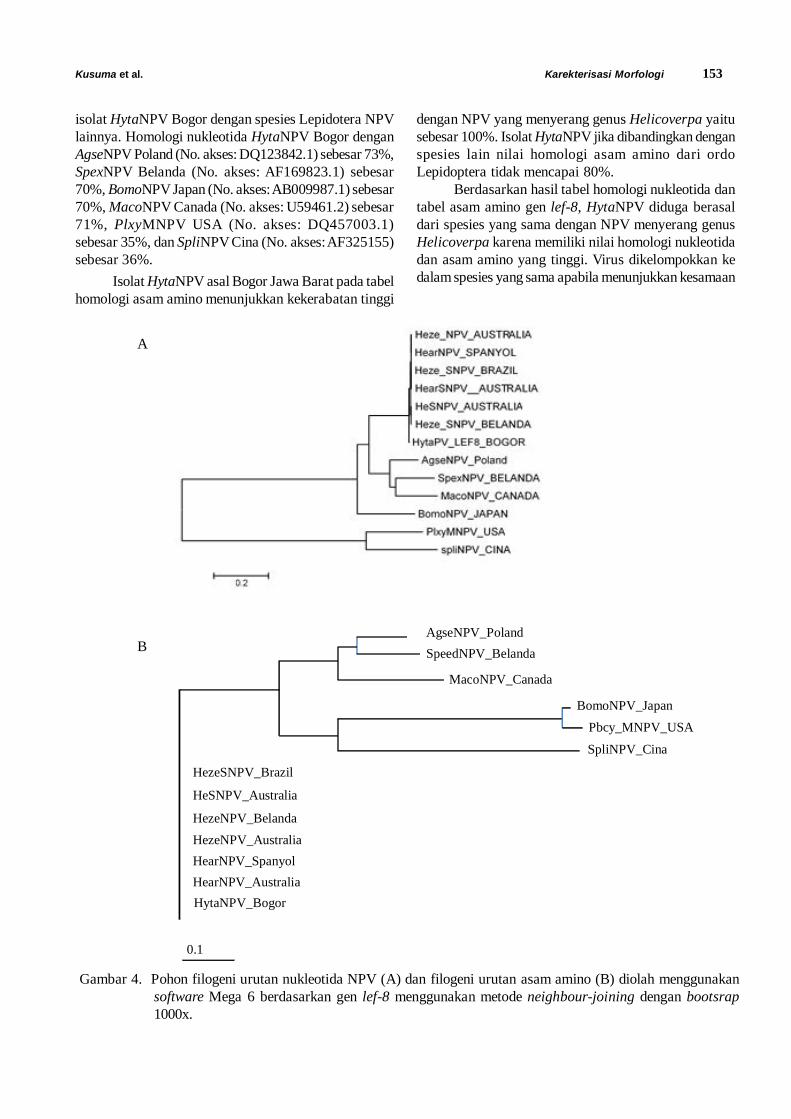
Kusuma et al. Karekterisasi Morfologi 153
isolat HytaNPV Bogor dengan spesies Lepidotera NPVlainnya. Homologi nukleotida HytaNPV Bogor denganAgseNPV Poland (No. akses: DQ123842.1) sebesar 73%,SpexNPV Belanda (No. akses: AF169823.1) sebesar70%, BomoNPV Japan (No. akses: AB009987.1) sebesar70%, MacoNPV Canada (No. akses: U59461.2) sebesar71%, PlxyMNPV USA (No. akses: DQ457003.1)sebesar 35%, dan SpliNPV Cina (No. akses: AF325155)sebesar 36%.
Isolat HytaNPV asal Bogor Jawa Barat pada tabelhomologi asam amino menunjukkan kekerabatan tinggi
dengan NPV yang menyerang genus Helicoverpa yaitusebesar 100%. Isolat HytaNPV jika dibandingkan denganspesies lain nilai homologi asam amino dari ordoLepidoptera tidak mencapai 80%.
Berdasarkan hasil tabel homologi nukleotida dantabel asam amino gen lef-8, HytaNPV diduga berasaldari spesies yang sama dengan NPV menyerang genusHelicoverpa karena memiliki nilai homologi nukleotidadan asam amino yang tinggi. Virus dikelompokkan kedalam spesies yang sama apabila menunjukkan kesamaan
Gambar 4. Pohon filogeni urutan nukleotida NPV (A) dan filogeni urutan asam amino (B) diolah menggunakansoftware Mega 6 berdasarkan gen lef-8 menggunakan metode neighbour-joining dengan bootsrap1000x.
A
B
HezeSNPV_Brazil
HeSNPV_Australia
HezeNPV_Belanda HezeNPV_Australia HearNPV_Spanyol HearNPV_Australia HytaNPV_Bogor
AgseNPV_Poland SpeedNPV_Belanda
MacoNPV_Canada
BomoNPV_Japan Pbcy_MNPV_USA
SpliNPV_Cina
0.1

154 J. HPT Tropika Vol. 17, No. 2, 2017: 147 - 155
sekuen nukleotida (tingkat homologi) di atas 89% (Kinget al., 2012).
Hasil tabel homologi urutan nukleotida dan urutanhomologi asam amino dari setiap isolat menunjukkanperbedaan. Homologi urutan nukelotida mempunyai nilaikekerabatan terbesar sebesar 98% sedangkan padahomologi urutan asam amino mempunyai nilai mencapai100%. Adanya perbedaan urutan basa nukleotida padaHytaNPV dengan NPV yang menyerang genusHelicoverpa yang tidak menimbulkan perbedaan padatranslasi dalam menghasilkan asam amino bisa terjadikarena adanya mutasi diam (silent mutation), yaituperubahan suatu pasang basa dalam gen yangmenimbulkan perubahan suatu kode genetik tetapi tidakmengakibatkan perubahan atau pergantian asam aminoyang dikode (Stansfield et al., 2003).
Analisis Filogeni HytaNPV. Konstruksi pohon filogeniurutan nukleotida dan pohon filogeni urutan asam amino(Gambar 4) dengan menggunakan metode Neighbor-Joining (NJ) menunjukkan sedikit perbedaan padapercabangan antara HytaNPV dengan NPV lainnya.HytaNPV dari Bogor pada pohon filogeni NJ beradadalam kelompok yang sama dengan NPV yangmenyerang genus Helicoverpa asal Australia, Belanda,Brazil, Spanyol, sedangkan NPV Lepidoptera dari genuslain berada pada kelompok yang terpisah.
SIMPULAN
Gen lef-8 berhasil mengidentifikasi HytaNPV,analisis urutan DNA berdasarkan gen lef-8 menunjukkanbahwa isolat HytaNPV Bogor tergolong ke dalamspesies virus yang memiliki hubungan kekerabatan yangdekat dengan isolat NPV yang menyerang genusHelicoverpa yang berasal dari Brazil, Australia,Spanyol, dan Belanda dengan nilai homologi nukleotidadan asam amino sebesar 98% dan 100%. Berdasarkananalisis filogeni, isolat HytaNPV masuk dalam grup yangsama dengan NPV yang menyerang genusHelicoverpa.
SANWACANA
Terimakasih kepada Perkebunan Gunung MasPTPN 8, Laboratorium Virologi Tumbuhan danLaboratorium Patologi Serangga, Departemen ProteksiTanaman, Fakultas Pertanian Institut Pertanian Bogoratas kerjasama serta fasilitas penelitian yang telahdiberikan selama penulis melaksanakan penelitian ini.
DAFTAR PUSTAKA
Acharya A & Gopinathan KP. 2002. Characterizationof late gene expression factors lef-9 and lef-8 fromBombyx mori nucleopolyhedrovirus. J. Gen Virol.83: 2015–2023.
Agustin HSW. 2013. Analisis potensi bakteri kitinolitikasal limbah udang dan kepiting kandidat penghasilenzim kitinase serta anti jamur Saprolegnia sp.menggunakan marka molekuler gen 16S rRNATesis. Universitas Padjajaran. Bandung.
Arifin M. 2006. Kompatibilitas SlNPV dengan HaNPVdalam pengendalian ulat grayak dan ulat pemakanpolong kedelai. Penelitian Pertanian TanamanPangan 25(1): 65–70.
Blissard G, Black B, Crook N, Keddie BA, Possee R,Rohrmann G, Theilmann D, & Volkman L. 2000.Family Baculoviridae. In: van Regenmortel MHV,Fauquet CM, Bishop DHL, Carstens EB, EstesMK, Lemon SM, McGeoch DJ, Maniloff J, MayoMA, Pringle CR, & Weckner RB (Eds.). VirusTaxonomy: Classification and Nomenclatureof Viruses. pp. 195–202. Seventh Report of theInternational Committee for the Taxonomy ofViruses. Academic Press, San Diego.
[BPS] Badan Pusat Statistik 2016. Produksi TanamanPerkebunan menurut Propinsi dan Jenis Tanaman,Indonesia (000 Ton), 2012-2015. BPS. Jakarta.
Cheng XW. 1998. Characterizatiom of twonuclearpolyhedrosis viruses from Thysanoplusiaorichalcea (L.) (Lepidoptera: Noctuidae) fromIndonesia. Dissertasion. Clemson University.Amerika Serikat.
Elrod SL & Stansfield WD. 2007. Genetika.Damaringtyas (penerjemah). Erlangga. Jakarta.
Ghosh B, Mukhopadhyay A, Das A, & Bahadur M.2015. Restriction endonuclease fragment analysisof Hyposidra talaca nucleopolyhedrovirusgenome. Int. J. Curr. Res. Aca. Rev. 3(8): 81–87.
Grzywacz D, Rabindra RJ, Brown M, Jones KA, &Parnell M. 2011. The Helicoverpa armigeraNPV Manual Production. FAO http://www.fao.org/docs/eims/upload/agrotech/2011/hanpvmanual-pt2. pdf. Diakses pada 3 April 2016.
Herniou EA, Luque T, Chen X, Vlak JM, WinstanleyD, Cory JS, & O’Reilly DR. 2003. Use of whole

Kusuma et al. Karekterisasi Morfologi 155
genome sequence data to infer Baculovirusphylogeny. J. Virol. 75(17): 8117–8126.
King AMQ, Adams MJ. Carstens EB, & Lefkowitz EJ.2012. Virus Taxonomy Classification andNomenclature of Viruses Nineth Report of theInternational Commite on Taxonomy of Viruses.Academic Press, San Diego.
Kaur B, Gupta VK, & Jindal V. 2014. Molecularfilogenetic analysis of Indian strains ofHelicoverpa armigera nucleopolyhedrovirus(HearNPV). Indian J. Biotechnol. 13: 186–194.
Kunimi Y. 2007. Current status and prospects onmicrobial control in Japan. J. Invertebr. Pathol.95: 181–186.
Lestari MP. 2008. Karakterisasi Morfologi danMolekuler Nucleopolyhedrovirus (NPV) padaHyposidra talaca Wlk (Lepidoptera:Geometridae). Skripsi. Institut Pertanian Bogor.Bogor.
Li Y, Yang BS, Wang H, Zia RX, Wang L, Zhang ZH,Qil L, & Liu YQ. 2009. Mitochondrial DNAanalysis reveals a low nucleotide diversity ofCaligula japonica in China. Afr. J. Biotec. 8(12):2707–2712.
Mukhopadhyay A, Khewa S, & De D. 2011.Characterist ics and virulence ofnucleopolyhedrovirus isolated from Hyposidratalaca (Lepidoptera: Geometridae), a pest of teain Darjeeling Terai, Indian. Int. J. Trop. Insect.Sci. 31(1–2): 13–19.
Muliani Y, Widayat W, & Solihin A. 2011. Penggunaanjamur entomopatogen Paecilomycesfumusoroseus Bain. (PFR) terhadap mortalitasulat jengkal (Hyposydra talaca Wlk.) hama padatanaman teh (Camellia sinensis (L). O. Kuntze).
Dalam: Intan A, Putra RE, Turmuktini T, MulianiY, Kantikowati E, Kinasih I, Meliansyah R, &Bari IN (Eds.). Hidup Sejahtera BersamaSerangga. Prosiding Seminar NasionalPerhimpunan Entomologi Indonesia (PEI)Cabang Bandung. pp. 449–453. Bandung, 16-17Februari 2011.
Pradana R. 2013. Pengelolaan Kebun dan UpayaPengendalian Ulat Jengkal dengan AplikasiHyposidra talaca Nucleopolyhedrovirus padaTanaman Teh di PT Perkebunan Nusantara VIIIGunung Mas Bogor Jawa Barat. Skripsi. InstitutPertanian Bogor. Bogor.
Pramono S. 2000. Efek penyimpanan terhadap virulensinuclear polyhedrosis virus yang diaplikasikan padatanaman kedelai terserang ulat grayak(Spodoptera litura Fabr.). J. HPT Tropika 1(1):29–32.
Stansfield WD, Colome JS, & Cano RJ. 2006. Schaum’sEasy Outlines Biologi Molekuler dan Sel.Fahmi V (Penerjemah). Erlangga. Jakarta.
Starnes RL, Liu CL, & Marrone PG. 1993. History,Use, and Future of Microbial Insecticides.American Entomologist, Amerika Serikat.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M,& Kumar S. 2011. MEGA5: MolecularEvolutionary. Cambridge University. AmerikaSerikat.
Widayat W. 1996. Hama-hama pada Tanaman Tehdan Cara Pengendaliannya. Balai PenelitianTeh dan Kina, Gambung. Bandung.
Yu W, Du YC, Quan YP, Nie ZM, Chen J, Lv ZB, &Zhang YZ. 2013. Characterization of late geneexpression factor LEF-10 of Bombyx morinucleopolyhedro virus. Virus Res. 175(1): 45–51.

Efrin et al. Aktivitas Insektisida Ekstrak Tithonia 185 J. HPT Tropika. ISSN 1411-7525Vol. 17, No. 2: 185 – 193, September 2017
AKTIVITAS INSEKTISIDA EKSTRAK TITHONIA DIVERSIFOLIA (HEMSL.)A. GRAY (ASTERACEAE) TERHADAP ULAT DAUN KUBIS
PLUTELLA XYLOSTELLA (L.) (LEPIDOPTERA: YPONOMEUTIDAE)
Efrin Firmansyah, Dadang, & Ruli Anwar
Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian BogorJl. Kamper Kampus IPB Dramaga, Bogor 16680
E-mail: [email protected]
ABSTRACT
Insecticidal activity of extracts of Tithonia diversifolia (Hemsl.) A. Gray (Asteraceae) against the diamondback mothPlutella xylostella (L.) (Lepidoptera: Yponomeutidae). The diamondback moth, Plutella xylostella (L.) (Lepidoptera:Yponomeutidae) is one of the most destructive pests on cruciferous plants. Generally farmers use synthetic insecticides tocontrol this pest. Intensive and excessive use of insecticides can cause some undesirable effects such as resistance, resurgence,and contamination of insecticide on environment. One effort to solve the problems caused by synthetic insecticides use is byutilization of botanical materials that are potential to be used as botanical insecticides, one of them is Tithonia diversifolia(Asteraceae). The purpose of this research was to study the insecticidal activity of T. diversifolia as a botanical insecticideagainst P. xylostella. The leaf residual and topical application methods were used to assess the mortality effect of the extracts.Insect mortality rate was analyzed using probit analysis to obtain LC50 and LC95 values. The results showed that the flowerextract had better activity on insect mortality than the leaf extract both in leaf residual and topical application treatments.
Key words: Botanical insecticides, environmentally friendly, mortality
ABSTRAK
Aktivitas insektisida ekstrak Tithonia diversifolia (Hemsl.) A. Gray (Asteraceae) terhadap ulat daun kubis Plutella xylostella(L.) (Lepidoptera: Yponomeutidae). Ulat daun kubis Plutella xylostella (L.) (Lepidoptera: Yponomeutidae) merupakan salahsatu hama yang merusak tanaman kubis-kubisan. Untuk mengendalikan hama tersebut petani umumnya menggunakan insektisidasintetik yang jika penggunaannya terus-menerus dapat menyebabkan resistensi, meningkatnya residu serta berpengaruhpada kesehatan lingkungan. Salah satu upaya yang dapat dilakukan untuk menyikapi masalah akibat penggunaan pestisidasintetik adalah dengan memanfaatkan bahan alami yang berpotensi untuk digunakan sebagai bahan baku insektisida nabatiyang salah satunya adalah Tithonia diversifolia (Asteraceae). Tujuan penelitian ini mempelajari aktivitas insektisida ekstrakmetanol daun dan bunga T. diversifolia terhadap ulat daun kubis P. xylostella. Dua metode uji digunakan untuk menilaipengaruh ekstrak T. diversifolia terhadap mortalitas larva P. xylostella yaitu metode residu pada daun dan metode aplikasitopikal. Tingkat mortalitas serangga diolah dengan menggunakan analisis probit untuk mendapatkan nilai LC50 dan LC95 darimasing-masing ekstrak. Hasil penelitian menunjukkan bahwa ekstrak bunga T. diversifolia memiliki efek mortalitas lebih tinggiterhadap P. xylostella daripada ekstrak daun baik pada perlakuan dengan metode residu pada daun maupun aplikasi topikal.
Kata kunci: insektisida nabati, mortalitas, ramah lingkungan
PENDAHULUAN
Tanaman kubis-kubisan (Brassicaceae) sepertisawi, kol kembang, brokoli, dan kubis merupakan jenistanaman sayuran yang banyak dibudidayakan oleh petaniIndonesia. Secara umum selama periode 2011-2015produksi tanaman kubis-kubisan mengalami fluktuasi(BPS, 2015). Beberapa faktor yang berkontribusi dalam
penurunan produksi tanaman di antaranya adalahpenurunan kesuburan lahan, berkurangnya lahanproduksi, dan adanya serangan organisme pengganggutanaman. Ulat daun kubis Plutella xylostella (L.)(Lepidoptera: Yponomeutidae) merupakan salah satuhama yang menyerang tanaman kubis-kubisan(Brassicaceae) di seluruh dunia (Sarfraz & Kaddie,2005). Kerusakan tanaman disebabkan oleh aktivitas

186 J. HPT Tropika Vol. 17 No. 2, 2017: 185 - 193
makan larva yang pada populasi tinggi dapat merusaktanaman secara berat kecuali pada bagian batangnya(Capinera, 2001).
Pengendalian secara kimia dengan insektisidasintetik merupakan teknik pengendalian yang palingumum dilakukan untuk mengendalikan P. xylostella diseluruh dunia (Syed et al., 2004). Penggunaaninsektisida sintetik selain efektif namun jikapenggunaannya secara terus menerus dapatmenyebabkan resistensi (Kau & Cheng, 2001). Kejadianresistensi hama P. xylostella terhadap insektisidapertama kali dilaporkan oleh Ankersmit (1953) yaituresistensi hama terhadap DDT di Indonesia. Selainresistensi, penggunaan insektisida sintetik yang terusmenerus secara tidak bijaksana juga berpengaruh padakesehatan lingkungan serta residu pada produk pertanianyang menimbulkan bahaya pada konsumen (Dadang &Prijono, 2008).
Salah satu upaya yang dapat dilakukan untukmeminimalisasi masalah akibat penggunaan pestisidasintetik adalah dengan mencari alternatif bahan alamiuntuk menggantikan penggunaan insektisida sintetik yangbanyak menimbulkan masalah lingkungan. Secara umuminsektisida nabati diartikan sebagai suatu insektisida yangbahan dasarnya adalah senyawa tumbuhan yang bersifatmudah terurai di alam (Mkenda et al., 2014), relatifaman terhadap organisme bukan sasaran termasukmusuh alami, dapat dipadukan dengan komponen lainpengendalian hama terpadu dan dapat memperlambatlaju resistensi (Dadang & Prijono, 2008). Berdasarkanhal tersebut penggunaan insektisida nabati diharapkanmampu mengurangi atau bahkan menggantikanpenggunaan pestisida sintetik di lapangan dan menjadibahan pengendali yang ramah lingkungan, aman, dansehat (Djunaedy, 2009).
Tithonia diversifolia (Asteraceae) yangmerupakan tumbuhan liar dan berpotensi menjadi gulmapada areal pertanian, belum banyak dimanfaatkan olehpetani-petani di Indonesia. Biasanya masyarakatmemanfaatkan tumbuhan ini sebagai pakan ternakruminansia. Pemanfaatan lainnya adalah sebagai pupukorganik (Pardono, 2011). Di luar negeri T. diversifoliaselain dimanfaatkan sebagai pakan ternak dan bahanpupuk organik, juga dimanfaatkan sebagai bahan bakuinsektisida nabati. T. diversifolia mengandungsenyawa-senyawa yang bersifat toksik terhadapserangga seperti flavonoid, tanin, dan triterpena yangberpotensi dapat dijadikan sebagai insektisida nabatiuntuk mengendalikan Atta cephalotes (Hymenoptera:Myrmicinae) (Castano-Quintana et al., 2013) dan larvaChlosyne lacinia (Lepidoptera: Nymphalidae)(Ambrosio et al., 2008). Hasil penelitian Carino &
Morallo-Rejesus (1982) menunjukkan tingkat toksikekstrak daun T. diversifolia terhadap P. xylostella24,47% lebih rendah dibandingkan dengan insektisidasintetik malathion, namun ekstrak tersebut lebih baik jikadibandingkan dengan tingkat toksiknya pada empatserangga lain (Dysdercus cingulatus Fab, Triboliumcastaneum Herbsr, Sitophilus zeamais Motsch, danSpodoptera exempta Walker). Di Indonesia,pemanfaatan T. diversifolia hanya sebagai pakanternak dan pupuk organik dan belum banyak dipelajaripotensinya sebagai pengendali hama tanaman atauinsektisida khususnya untuk mengendalikan ulat daunkubis P. xylostella. Oleh karena itu tujuan dari penelitianini untuk mengetahui aktivitas insektisida ekstrak metanoldaun dan bunga T. diversifolia terhadap larva P.xylostella.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilaksanakan diLaboratorium Fisiologi dan Toksikologi Serangga,Departemen Proteksi Tanaman, Institut Pertanian Bogordengan suhu rata-rata 26,6 oC dan rata-rata kelembapan67,5% dari September 2015 sampai Mei 2016.
Penanaman dan Pemeliharaan Brokoli. Tanamanbrokoli (Brassica oleraceae L. var. italica) yangdigunakan sebagai media untuk menguji pengaruhekstrak T. diversifolia terhadap larva P. xyostelladitanam di lahan dengan menggunakan polybag. Benihbrokoli disemai pada wadah penyemaian yang berisitanah dan pupuk kandang 2:1 (v/v) yang telah dicampurrata. Setelah berumur 3 minggu, sebanyak satu bibitditanam pada polybag berukuran besar (5 liter) yangtelah berisi campuran tanah dan pupuk kandang 2:1 (v/v). Pemupukan tambahan dilakukan ketika tanamanberumur 3-4 minggu melalui pemberian pupuk NPKdengan dosis 1 g/tanaman. Pemeliharaan tanamanbrokoli meliputi penyiraman yang dilakukan dua kalisehari, penyulaman terhadap tanaman yang mati,penyiangan gulma, dan pengendalian hama dilakukansecara mekanis. Daun dari tanaman berumur 2 bulanatau lebih digunakan sebagai pakan larva P. xylostella.
Pemeliharaan dan Perbanyakan Serangga Uji.Serangga yang digunakan sebagai hama sasaran adalahlarva P. xylostella instar III generasi kedua berdasarkanpemeliharaan di laboratorium. Imago seranggadidapatkan dari daerah Cipanas, Cianjur (6o44’58.81"LS dan 107o1’17.47" BT) yang dikumpulkan dandipelihara di laboratorium dengan cara dimasukkan kedalam kurungan kasa tempat pemeliharan (50 x 50 x 50

Efrin et al. Aktivitas Insektisida Ekstrak Tithonia 187
cm). Imago tersebut diberi pakan larutan madu 10% (v/v) yang diserapkan pada kapas yang digantungkan ditengah-tengah kurungan menggunakan seutas benang.Pada bagian dasar kurungan diletakkan daun brokolipada pot kecil berisi air sebagai tempat peletakan telur.Daun yang telah diletaki telur dipindahkan ke dalamkotak plastik (30 x 25 x 7 cm) tempat pemeliharaan.Telur-telur tersebut dipelihara hingga menetas kemudiansetelah menjadi larva, serangga diberi makan daunbrokoli bebas pestisida.
Ekstraksi Tumbuhan. Metode yang digunakan untukmendapatkan ekstrak T. diversifolia adalah metodemaserasi (Dadang & Prijono, 2011). Tanaman diperolehdari kebun pertanian organik di daerah Cisarua Bogor(6o41’20.30" LS dan 106o56’55.63 BT). Bagian tanamanyang digunakan sebagai bahan pembuatan ekstrak adalahbagian daun dan bunga. Setiap bahan tersebut dipotongmenjadi bagian-bagian kecil kemudian dikeringanginkanselama 7 hari di laboratorium, lalu dihaluskanmenggunakan blender secara terpisah hingga menjadiserbuk. Serbuk yang terbentuk diayak menggunakanpengayak kasa berjalinan 1 mm. Setiap serbuk yang telahdihasilkan direndam dalam pelarut metanol sebanyak duakali dengan perbandingan 1:10 (w/v) selama 48 jam.Rendaman kemudian disaring menggunakan corongBuchner yang dialasi kertas saring Whatman. Hasilsaringan kemudian diuapkan menggunakan rotaryevaporator pada suhu 50 oC dan tekanan 240 mbarsehingga diperoleh ekstrak kasar. Ekstrak yang diperolehdisimpan dalam lemari es (4 o C) hingga saat digunakan.
Uji Toksisitas Ekstrak terhadap LarvaP. xylostella. Setiap ekstrak diuji pada lima tarafkonsentrasi yang diharapkan dapat mengakibatkankematian serangga uji 5% hingga 95%, yang ditentukandengan uji pendahuluan. Konsentrasi ekstrak yangdigunakan untuk uji pendahuluan adalah 5%, 3%, 1%,0,5%, dan 0,1%. Setelah diperoleh data mortalitas hasiluji pendahuluan kemudian dilakukan analisis probit untukmemperoleh konsentrasi yang akan digunakan pada ujilanjutan. Konsentrasi uji lanjut ekstrak metanol daun T.diversifolia berturut-turut 0,02%, 0,04%, 0,09%, 0,22%,dan 0,97% serta untuk ekstrak bunga berturut-turut0,01%, 0,04%, 0,08%, 0,19%, dan 0,86%. Pembuatankonsentrasi larutan ekstrak dilakukan dengan metodepengenceran berseri (serial dilution). Setiap larutanekstrak mengandung metanol sebanyak 1% dan Tween-80 0,2%. Pengujian toksisitas dilakukan dengan duametode yaitu metode residu pada daun dan metodeaplikasi topikal.
Metode residu pada daun. Potongan daun brokoli (4cm x 4 cm) dicelupkan ke dalam sediaan ekstrak padakonsentrasi tertentu kemudian dikeringanginkan. Daunkontrol dicelupkan ke dalam 100 ml larutan air yangmengandung 1,2% metanol dan Tween 80 (5:1, v/v)(Dadang & Prijono, 2008). Sebanyak dua lembar daundimasukkan ke dalam cawan petri (diameter 9 cm), laludimasukkan 10 larva P. xylostella instar III. Setiapperlakuan dan kontrol diulang lima kali. Daun perlakuandan kontrol diganti setiap 24 jam setelah perlakuandengan daun tanpa perlakuan. Pengamatan mortalitaslarva dilakukan setiap 24 jam selama 6 hari. Persentasekematian untuk setiap konsentrasi ekstrak dianalisisdengan program analisis probit POLO-PC untukmenentukan hubungan konsentrasi/dosis dengankematian serangga uji (Finney, 1971).
Metode aplikasi topikal. Sebanyak 10 larva P.xylostella instar III dimasukkan ke dalam cawan petri(diameter 9 cm) yang telah dialasi tisu. Larva ditetesi 1µl larutan ekstrak pada masing-masing perlakuan ataukontrol yang mengandung air, metanol (1%) dan Tween80 (0,2%) pada bagian dorsal toraks menggunakanmicroapplicator. Setelah tetesan kering, dua lembardaun brokoli (4 x 4 cm) tanpa perlakuan diberikansebagai pakan. Setiap perlakuan dan kontrol diulangsebanyak lima kali. Pengamatan dilakukan setiap 24 jamselama 6 hari dengan menghitung jumlah larva yang mati.
Analisis Data. Data mortalitas ditabulasi pada programkomputer Microsoft Excel 2007 kemudian dianalisisprobit menggunakan program POLO-PC (LeOraSoftware, 1987) untuk mendapatkan LC50 dan LC95.
HASIL DAN PEMBAHASAN
Larva P. xylostella pada perlakuan ekstrak daunT. diversifolia dengan metode residu pada daun mulaimengalami kematian pada 48 jam setelah perlakuan (JSP)sampai 144 JSP kecuali pada perlakuan 0,02% dan0,04%, kematian larva mulai terjadi pada 72 JSP.Pengamatan 96 JSP menunjukkan kematian larva kurangdari 50% pada semua perlakuan. Persentase kematiantertinggi dan terendah yaitu masing-masing sebesar 48%dan 10% terjadi pada perlakuan dengan konsentrasi0,97% dan 0,02%. Pengamatan 144 JSP menunjukkanmortalitas larva 94%, 86%, 72%, 32%, dan 18% padakonsentrasi berturut-turut 0,97%, 0,22%, 0,09%, 0,04%,dan 0,02% (Gambar 1A).
Perkembangan mortalitas larva P. xylostellapada perlakuan ekstrak bunga T. diversifolia dengan

188 J. HPT Tropika Vol. 17 No. 2, 2017: 185 - 193
metode residu pada daun menunjukkan kematian larvadimulai sejak 24 JSP sampai 144 JSP kecuali padaperlakuan 0,01%, kematian larva mulai terjadi pada 48JSP. Pengamatan 96 JSP menunjukkan kematian larvamencapai 50% pada konsentrasi 0,86% dan 26% padakonsentrasi 0,01%. Pengamatan 144 JSP menunjukkanmortalitas larva mencapai 98%, 84%, 76%, 66%, dan44% pada konsentrasi berturut-turut 0,86%, 0,19%,0,08%, 0,04%, dan 0,01% (Gambar 1B).
Kematian larva P. xylostella pada perlakuanekstrak daun T. diversifolia dengan metode aplikasitopikal terjadi sejak 24 JSP sampai 144 JSP. Peningkatankematian terjadi pada setiap waktu pengamatan dan
peningkatan yang tinggi terjadi pada pengamatan 96 JSP,yang menunjukkan mortalitas mencapai 80%, 76%, 60%,52%, dan 42% pada dosis berturut-turut 9,7, 2,2, 0,9,0,4, dan 0,2 mg/larva. Pengamatan pada 144 JSPmenunjukkan kematian larva mencapai berturut-turut96%, 88%, 80%, 70%, dan 46% pada perlakuan dengankelima dosis tersebut (Gambar 2A).
Perkembangan mortalitas larva P. xylostella padaperlakuan ekstrak bunga T. diversifolia dengan metodeaplikasi topikal menunjukkan kematian larva mencapai46% pada dosis 8,6 mg/larva pada pengamatan 24 JSP.Perkembangan mortalitas pada perlakuan ini relatif lebihcepat daripada perlakuan dengan metode residu pada
Gambar 1. Perkembangan mortalitas larva P. xylostella pada perlakuan ekstrak daun (A) dan bunga (B) T.diversifolia dengan metode residu pada daun.

Efrin et al. Aktivitas Insektisida Ekstrak Tithonia 189
daun baik dengan metode residu pada daun maupunmetode aplikasi topikal. Pada 96 JSP kematian mencapai100%, 92%, 76%, 76% dan 50% pada dosis berturut-turut 8,6, 1,9, 0,8, 0,4, dan 0,1 mg/larva. Pengamatanpada 144 JSP menunjukkan kematian larva mencapaiberturut-turut 100%, 98%, 90%, 84%, dan 72% padaperlakuan dengan kelima dosis tersebut (Gambar 2B).
Berdasarkan analisis probit, ekstrak bunga lebihtoksik daripada ekstrak daun baik pada metode residupada daun maupun metode aplikasi topikal. Melaluimetode residu pada daun LC50 dan LC95 berturut-turut0,06% dan 0,64% untuk ekstrak daun serta 0,02% dan0,56% untuk ekstrak bunga. Sementara itu LD50 dan
LD95 ekstrak bunga yang diaplikasikan dengan metodeaplikasi topikal berturut-turut sebesar 0,04 dan 1,32 mg/larva yang lebih rendah daripada LD50 dan LD95 ekstrakdaun yang berturut-turut 0,17 dan 5,14 mg/larva (Tabel1).
Hasil penelitian ini menunjukkan terdapatpengaruh pemberian ekstrak daun dan bunga T.diversifolia terhadap mortalitas larva P. xylostella. Halini membuktikan bahwa tumbuhan T. diversifoliaberpotensi untuk dijadikan sebagai insektisida nabati(Isman, 2014). Beberapa penelitian lain yangmenunjukkan potensi tersebut di antaranya adalah Rusliet al. (2010) yang melaporkan bahwa T. diversifolia
Gambar 2. Perkembangan mortalitas larva P. xylostella pada perlakuan ekstrak daun (A) dan bunga (B) T.diversifolia dengan metode aplikasi topikal.

190 J. HPT Tropika Vol. 17 No. 2, 2017: 185 - 193
dapat digunakan sebagai insektisida untuk mengendalikanSpodoptera exigua. Ekstrak daun T. diversifoliadilaporkan berpotensi untuk dijadikan sebagai insektisidanabati untuk mengendalikan hama gudangCallosobruchus maculatus (Coleoptera: Bruchidae)(Adedire & Akinneye, 2004; Kolawole et al., 2014),larva nyamuk Aedes aegypti (Bernard et al., 2012),larva Chrysomya bezziana (Wardhana & Diana 2014),Macrotermes bellicosus (Oyedokun et al. 2011) danMacrotermes spp. (Osipitan & Oseyemi, 2012).Tithonia diversifolia juga dapat dimanfaatkan untukmengendalikan Sitophilus zeamais (Onekutu et al.,2015) dan Atta cephalotes (Hymenoptera: Formicidae)(Zanetti et al., 2014, Rodriguez et al., 2015).
Tithonia diversifolia diketahui mengandungbeberapa senyawa metabolit sekunder diantaranyaseskuiterpena, diterpena, monoterpena, dan kandunganalisiklik yang bersifat toksik yang berasal dari bagiandaun, batang, atau bunga (Mkenda et al., 2014; Mkenda& Ndakidemi, 2014). Tanaman ini juga diketahuimengandung senyawa glikosida, tanin, flavonoid, saponin,fenol, dan alkaloid (Otusanya & Ilori, 2012). Beberapasenyawa golongan sesquiterpen yang terdapat padaT. diversifolia adalah germakren sesquiterpena (1-asetiltagitinin-A) dan guanianana sesquiterpena (8-isobutiliriloksikumambranolida) (Kuo & Chen, 1998).
Hasil penelitian menunjukkan perlakuan ekstrakbunga dengan metode aplikasi topikal lebih toksikdaripada ekstrak daun dengan metode aplikasi topikaldan metode residu pada daun. Hasil tersebut sejalandengan penelitian Mukasa et al. (2008) yangmenunjukkan bahwa ekstrak bunga T. diversifolia lebihtoksik daripada ekstrak daun terhadap Cosmopolitessordidus (Germar) (Coleoptera: Curculionidae). Fouadet al. (2014) menyatakan ekstrak bunga T. diversifolia
dengan metode aplikasi topikal menjadi salah satu bahanalami yang berpotensi dijadikan sebagai insektisida nabatiuntuk mengendalikan hama Sitotroga cereallea(Lepidoptera: Gelechiidae). Hasil penelitian Carino &Morallo-Rejesus (1982) menunjukkan aplikasi topikalpada serangga uji yang memiliki tekstur kutikula halusseperti larva P. xylostella akan meningkatkan tingkattoksik ekstrak T. diversifolia yang diaplikasikan karenapenetrasi senyawa kimia pada kutikula serangga tersebutterjadi lebih efisien.
Ekstrak bunga T. diversifolia mengandungsenyawa fenol, tanin dan flavonoid (da Gama et al.,2014). Menurut Utami et al. (2010) senyawa fenol dapatberperan sebagai perangsang aktivitas makan yangmengakibatkan serangga uji mati karena banyakmemakan senyawa toksik pada daun perlakuan (dauncaisim). Hasil penelitian Mwanauta et al. (2014)menunjukkan beberapa senyawa monoterpena yangterkandung dalam T. diversifolia yaitu b-osimena, a-pinena, dan limonena. Menurut Ibrahim et al. (2001)senyawa monoterpena (limonene) bersifat toksik yangmasuk melalui lapisan kutikula (racun kontak), saluranpernapasan dan saluran pencernaan (racun perut) dapatmengakibatkan kematian serangga. Adapunseskuiterpena dan flavonoid mampu masuk ke dalammembran lipid bilayer yang mengakibatkan fluiditasmembran dan permeabilitas sel otot meningkat. Kondisiini akan melemahkan gerakan serangga dan berakhirdengan kematian (Cestari et al., 2004). Perbedaantingkat mortalitas yang ditimbulkan masing-masingperlakuan dapat diakibatkan oleh adanya perbedaankarakteristik dari senyawa metabolit sekunder yangterkandung di dalam masing-masing ekstrak termasukperbedaan dari cara kerja senyawa tersebut (War etal., 2013).
Tabel 1. Penduga parameter regresi probit hubungan antara konsentrasi atau dosis ekstrak T. diversifoliadengan mortalitas larva P. xylostella pada 144 JSP
a DR = ekstrak daun dengan metode residu pada daun, BR = ekstrak metanol bunga dengan metode residu padadaun, DT = ekstrak daun dengan metode aplikasi topikal. BT = ekstrak bunga dengan metode aplikasi topikal. b a=intersep garis regresi probit, c b= kemiringan regresi probit, d GB : galat baku. e LC : lethal concentration, f SK :selang kepercayaan, g LD= lethal dose.
Perlakuana ab ± GB bc ± GBd LC50e (SKf 95%) (%) LC95 (SK 95%) (%)
DR 1,95 ± 0,23 1,57 ± 0,19 0,06 (0,023-0,114) 0,64 (0,252-11,213)BR 1,92 ± 0,24 1,09 ± 0,18 0,02 (0,010-0,031) 0,56 (0,291-1,87)
LD50g (SK 95%)
(g/larva) LD95 (SK 95%)
(g/larva) DT 0,83 ± 0,10 1,04 ± 0,18 0,16 (0,07-0,27) 6,02 (3,07-21,07) BT 1,56 ± 0,16 1,17 ± 0,26 0,04 (0,015-0,095) 1,32 (0,72-6,65)

Efrin et al. Aktivitas Insektisida Ekstrak Tithonia 191
SIMPULAN
Secara umum baik ekstrak daun maupunekstrak bunga T. diversifolia mengakibatkan kematianlarva P. xylostella. Ekstrak bunga T. diversifoliadengan metode aplikasi topikal memilliki aktivitasmortalitas lebih tinggi dari pada ekstrak daun terhadaplarva P. xylostella baik pada perlakuan dengan metoderesidu pada daun maupun aplikasi topikal.
SANWACANA
Penulis mengucapkan terima kasih kepadaLembaga Pengelola Dana Keuangan (LPDP)Kementerian Keuangan RI atas dukungan seluruhpendanaan penelitian. Penulis juga mengucapkan terimakasih kepada Ir. Djoko Prijono, MAgr.Sc. sebagaisebagai Kepala Laboratorium Fisiologi dan ToksikologiSerangga, Departemen Proteksi Tanaman, InstitutPertanian Bogor.
DAFTAR PUSTAKA
Adedire CO & Akinneye JO. 2004. Biological activityof tree marigold, Tithonia diversifolia, oncowpea seed bruchid, Callosobruchusmaculatus (Coleoptera: Bruchidae). Ann. Appl.Biol. 144(2): 185–189.
Ambrósio SR, Oki Y, Heleno VCG, Chaves JS,Nascimento PGBD, Lichston JE, ConstantinoMG, Varanda EM, & Da Costa FB. 2008.Constituents of glandular trichomes of Tithoniadiversifolia: relationships to herbivory andantifeedant activity. Phytochemistry 69(10):2052–2060.
Ankersmit GW. 1953. DDT-resistance in Plutellamaculipennis (Curt.) (Lep.) in Java. Bull.Entomol. Res. 44(3): 421–425.
Bernard KL, David SK, Mark NO, Charles MK,Geoffrey RM, Charles M, Laban I, & Willy TK.2012. Larvicidal action of extracts from Tithoniadiversifolia against the dengue mosquito Aedesaegypti (Diptera: Culicidae). JBAPN 2(1): 46–49.
[BPS] Badan Pusat Statistik. 2015. Produksi sayurandi Indonesia, 2011-2015.http:// pertanian.go.id/D a t a 5 t a h u n / H o r t i A S E M 2 0 1 5 / 3 -Produksi%20Nasional%20Sayuran.pdf. Diaksestanggal 16 Mei 2016.
Capinera JL. 2001. Handbook of Vegetable Pests.Academic Press. San Diego.
Carino FA & Morallo-Rejesus B. 1982. Isolation andcharacterization of the insecticidal fraction fromTithonia diversifolia (A. Gray) leaves. AnnalsTrop. Res. 4(1): 1–11.
Castańo-Quintana K , Montoya-Lerma J, & Giraldo-Echeverri C. 2013. Toxicity of foliage extracts ofTithonia diversifolia (Asteraceae) on Attacephalotes (Hymenoptera: Myrmicinae)workers. Ind. Crops Prod. 44: 391–395.
Cestari IM, Sarti SJ, Waib CM, & Branco Jr AC. 2004.Evaluation of the potential insecticide activity ofTagetes minuta (Asteraceae) essential oil againstthe head lice Pediculus humanus capitis(Phthiraptera: Pediculidae). Neotrop. Entomol.33(6): 805–807.
Dadang & Prijono D. 2008. Insektisida Nabati:Prinsip, Pemanfaatan dan Pengembangan.Departemen Proteksi Tanaman, FakultasPertanian, Institut Pertanian Bogor.
Dadang & Prijono D. 2011. Pengembangan teknologiformulasi insektisida nabati untuk pengendalianhama sayuran dalam upaya menghasilkan produksayuran sehat. JIPI 6(2): 100–111.
da Gama RM, Guimarćes M, de Abreu LC, & Armando-Junior J. 2014. Phytochemical screening andantioxidant activity of ethanol extract of Tithoniadiversifolia (Hemsl) A. Gray dry flowers. AsianPac. J. Trop. Biomed. 4(9): 740–742.
Djunaedy A. 2009. Biopestisida sebagai pengendaliorganisme pengganggu tanaman (OPT) yangramah lingkungan. Embryo 6(1): 88–95.
Finney DJ. 1971. Probit Analysis, 3rd ed. CambridgeUniversity Press, Cambridge.
Fouad HA, Faroni LRDA, Tavares WS, Ribeiro RC,Freitas SS, & Zanuncio JC. 2014. Botanicalextracts of plants from the Brazilian Cerrado forthe integrated management of Sitotrogacerealella (Lepidoptera: Gelechiidae) in storedgrain. J. Stored Prod. Res. 57: 6–11.

192 J. HPT Tropika Vol. 17 No. 2, 2017: 185 - 193
Ibrahim MA, Kainulainen P, Aflatuni A, Tiilikkala K, &Holopainen JK. 2001. Insecticidal, repellent,antimicrodial activity and phytotoxicity of essentialoils: with special reference to limonene and itssuitability for control of insect pests. Agri. FoodSci. Finland 10: 243–259.
Isman MB. 2014. Botanical insecticides: a globalperspective. In: Biopesticide: State of the Artand the Future Opportunities. pp. 21-30. ACS.Washington, D.C.
Kau CH & Cheng EY. 2001. Insecticides resistence inPlutella xylostella L. J. Agric. Res. China.50(4): 80–89.
Kuo YH & Chen CH. 1998. Sesquiterpenes from theleaves of Tithonia diversifolia. J. Nat. Prod.61(6): 827–828.
Kolawole AO, Olajuyigbe FM, Ajele JO, & Adedire JO.2014. Activity of the antioxidant defense systemin a typical bioinsecticide-and syntheticinsecticide-treated cowpea storage beetleCallosobruchus maculatus F. (Coleoptera:Chrysomelidae). Int. J. Insect Sci. 6: 99–108.
LeOra Software. 1987. POLO-PC User’s Guide.LeOra Software. Petaluma.
Mkenda PA, Mtei K, & Ndakidemi PA. 2014. Pesticidalefficacy of Tephrosia vogelii and Tithoniadiversifolia against field insect pests of commonbeans [Phaseolus vulgaris L.] within Africanfarming communities. Afr. J. Appl. Agric. Sci.Technol. 2(1): 9–26.
Mkenda PA & Ndakidemi PA. 2014. Pesticidal efficacyof four botanical pesticides on survival, ovipositionand progeny development of bruchid,Callosobruchus maculatus in stored cowpea,Vigna unguiculata. IJPS 3(12): 1504–1523.
Mukasa D, Olila D, Tinzaara W, & Kagezi GH. 2008.Effects of Tithonia and Phytolacca extractsagainst the banana weevil, Cosmopolitessordidus (Germar) (Coleoptera: Curculionidae).AJABS 3(1): 24–29.
Mwanauta RW, Mtei KA, & Ndakidemi PA. 2014.Prospective bioactive compounds from Vernoniaamygdalina, Lippia javanica, Dysphaniaambrosioides and Tithonia diversifolia incontrolling legume insect pests. Agri Sci 5(12):1129–1139.
Onekutu A, Nwosu LC, & Abakpa RE. 2015.Comparative efficacy of aqueous extracts ofTithonia diversifolia (Asteraceae) and Vernoniaamygdalina (Asteraceae) leaves in themanagement of Sitophilus zeamais infestationin stored maize. IOSR-JPBS 10(3): 66–70.
Osipitan AA & Oseyemi AE. 2012. Evaluation of thebio-insecticidal potential of some tropical plantextracts against termites (Termitidae: Isoptera)in Ogun State, Nigeria. J. Entomol. 9: 257–265.
Otusanya O & Ilori O. 2012. Phytochemical screeningand the phytotoxic effects of aqueous extracts ofTithonia diversifolia (Hemsl) A. Gray. Int. J.Biol 4(3): 97–101.
Oyedokun AV, Anikwe JC, Okelana FA, Mokwunye IU,& Azeez OM. 2011. Pesticidal efficacy of threetropical herbal plants’ leaf extracts againstMacrotermes bellicosus, an emerging pest ofcocoa, Theobroma cacao L. J. Biopestic. 4(2):131–137.
Pardono. 2011. Potensi Chromolaena odorata danTithonia diversifolia sebagai sumber nutrisi bagitanaman berdasarkan kecepatan dekomposisinya.Agrovigor 4(2): 80–85.
Rodriguez J, Montoya-Lerma J, & Calle Z. 2015. Effectof Tithonia diversifolia mulch on Attacephalotes (Hymenoptera: Formicidae) nests. J.Insect Sci. 15(32): 1–7.
Rusli R, Arneti, & Permata SS. 2010. Pengujian ekstrakmetanol bunga kipat (Tithonia diversifolia A.Gray) (Asteraceae) untuk mengendalikanSpodoptera exigua Hubner (Lepidoptera:Noctuidae). J. Manggaro 11(1): 25–32.
Sarfraz M & Keddie BA. 2005. Conserving the efficacyof insecticides against Plutella xylostella (L.)(Lep., Plutellidae). J. Appl. Entomol. 129(3):149–157.

Efrin et al. Aktivitas Insektisida Ekstrak Tithonia 193
Syed TS, Abro GH, & Ahmed S. 2004. Efficacy ofdifferent insecticides against Plutella xylostellaunder field conditions. Pak. J. Biol. Sci. 7(1):10–13.
Utami S, Syaufina L, & Haneda NF. 2010. Daya racunekstrak kasar daun bintaro (Cerbera odollamGaertn.) terhadap larva Spodoptera lituraFabricius. JIPI 15(2): 96–100.
War AR, Paulraj MG, Husain B, Buhroo AA,Ignacimuthu S, & Sharma HC. 2013. Effect ofplant secondary metabolites on legume pod borer,Helicoverpa armigera. J. Pest. Sci. 86(3): 399–408.
Wardhana AH & Diana N. 2014. Aktivitas biolarvasidalekstrak metanol daun kipahit (Tithoniadiversifolia) terhadap larva lalat Chrysomyabezziana. JITV 19(1): 43–51.
Zanetti R, Zanuncio JC, Santos JC, Silva WLP, RibeiroGT, & Lemes PG. 2014. An overview ofintegrated management of leaf-cutting ants(Hymenoptera: Formicidae) in Brazilian forestplantations. Forests 5(3): 439–454.