ii. tinjauan pustaka - repository.ipb.ac.id · variasi juga terjadi pada panjang rambut3. di daerah...
TRANSCRIPT

10
II. TINJAUAN PUSTAKA
2.1 Macan Tutul
2.1.1 Taksonomi
Macan tutul merupakan salah satu jenis kucing besar dari genus Panthera dalam
famili Felidae. Spesies macan tutul adalah Panthera pardus Linnaeus, 1758.
Berdasarkan analisis taksonomi moderen ada delapan atau sembilan sub spesies, salah
satunya adalah macan tutul jawa (Panthera pardus melas Cuvier, 1809) yang hanya
terdapat di Pulau Jawa1. Di beberapa daerah di Indonesia macan tutul dikenal dengan
nama lokal, misalnya di Jawa dikenal dengan nama macan, sima, macan tutul, seruni,
kombang, gogor, pogoh, bungbak; di Jawa Barat disebut meong hideung, kerud anjing,
rimau lalat, meong krut; di Madura dikenal dengan nama macan totol (Direktorat PPA,
1978).
Di Indonesia, macan tutul jawa sudah menjadi satwa yang dilindungi sejak tahun
1970 berdasarkan Keputusan Menteri Pertanian Nomor 421/Kpts/Um/8/1970. Pada
tahun 1999 status perlindungannya dipertegas lagi dengan Peraturan Pemerintah
Republik Indonesia Nomor 7 tahun 1999 tentang Pengawetan Tumbuhan dan Satwa.
Sejak tahun 1978 macan tutul telah dimasukkan ke dalam Red list IUCN
(International Union for Conservation of Nature & Natural Resources) dengan kategori
Vulnerable. Sejak itu kategorinya terus berubah, tahun 1988 menjadi Threatened, tahun
1994 menjadi Indeterminate, pada tahun 1996 menjadi Endangered dan pada tahun
2008 menjadi Critically Endangered (IUCN-The World Conservation Union, 1996;
Ario et al., 2008).
2.1.2 Deskripsi Fisik
Wilayah dan habitat memiliki pengaruh pada penampilan macan tutul. Di
Afrika, macan tutul yang hidup di daerah berbukit cenderung lebih besar daripada yang
hidup di dataran rendah (Guggisberg, 1975; Nowak, 1997). Secara umum ukuran tubuh
macan tutul jantan 20 – 40 % lebih besar daripada betina (Garman, 1997). Ukuran rata-
rata tubuh macan tutul yang hidup di Pulau Jawa disajikan pada Tabel 2.1 (Hoogerwerf,
1970).
1 http://www.lioncrusher.com/animal.asp?animal=57
11
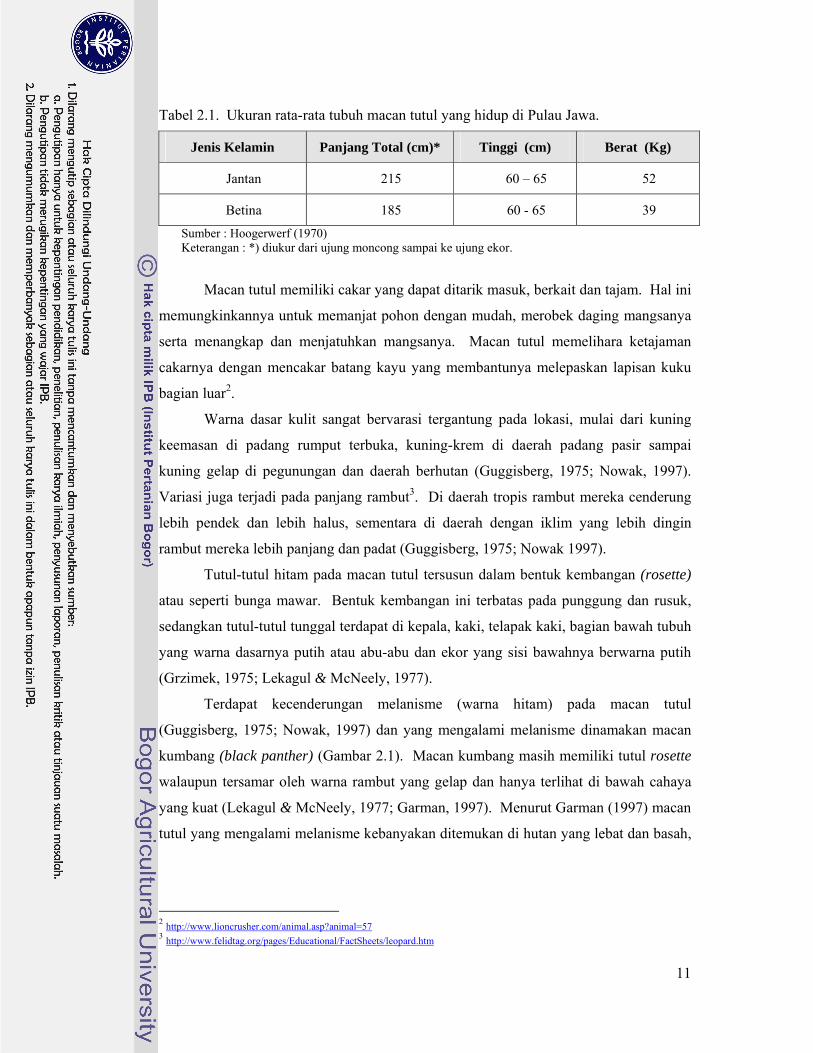
Tabel 2.1. Ukuran rata-rata tubuh macan tutul yang hidup di Pulau Jawa.
Jenis Kelamin Panjang Total (cm)* Tinggi (cm) Berat (Kg)
Jantan 215 60 – 65 52
Betina 185 60 - 65 39
Sumber : Hoogerwerf (1970) Keterangan : *) diukur dari ujung moncong sampai ke ujung ekor.
Macan tutul memiliki cakar yang dapat ditarik masuk, berkait dan tajam. Hal ini
memungkinkannya untuk memanjat pohon dengan mudah, merobek daging mangsanya
serta menangkap dan menjatuhkan mangsanya. Macan tutul memelihara ketajaman
cakarnya dengan mencakar batang kayu yang membantunya melepaskan lapisan kuku
bagian luar2.
Warna dasar kulit sangat bervarasi tergantung pada lokasi, mulai dari kuning
keemasan di padang rumput terbuka, kuning-krem di daerah padang pasir sampai
kuning gelap di pegunungan dan daerah berhutan (Guggisberg, 1975; Nowak, 1997).
Variasi juga terjadi pada panjang rambut3. Di daerah tropis rambut mereka cenderung
lebih pendek dan lebih halus, sementara di daerah dengan iklim yang lebih dingin
rambut mereka lebih panjang dan padat (Guggisberg, 1975; Nowak 1997).
Tutul-tutul hitam pada macan tutul tersusun dalam bentuk kembangan (rosette)
atau seperti bunga mawar. Bentuk kembangan ini terbatas pada punggung dan rusuk,
sedangkan tutul-tutul tunggal terdapat di kepala, kaki, telapak kaki, bagian bawah tubuh
yang warna dasarnya putih atau abu-abu dan ekor yang sisi bawahnya berwarna putih
(Grzimek, 1975; Lekagul & McNeely, 1977).
Terdapat kecenderungan melanisme (warna hitam) pada macan tutul
(Guggisberg, 1975; Nowak, 1997) dan yang mengalami melanisme dinamakan macan
kumbang (black panther) (Gambar 2.1). Macan kumbang masih memiliki tutul rosette
walaupun tersamar oleh warna rambut yang gelap dan hanya terlihat di bawah cahaya
yang kuat (Lekagul & McNeely, 1977; Garman, 1997). Menurut Garman (1997) macan
tutul yang mengalami melanisme kebanyakan ditemukan di hutan yang lebat dan basah,
2 http://www.lioncrusher.com/animal.asp?animal=57 3 http://www.felidtag.org/pages/Educational/FactSheets/leopard.htm

12
dimana warna ini bermanfaat dalam perburuan. Di Jawa sebagian besar macan tutul
mengalami melanisme4.
Robinson (1969) dalam Hoogerwerf (1970) menyatakan bahwa bila fase tutul
kawin dengan fase tutul maka perbandingan anak-anaknya adalah tiga tutul dan satu
kumbang, bila fase tutul kawin dengan fase kumbang maka perbandingan anak-anaknya
adalah satu tutul dan satu kumbang, dan bila fase kumbang kawin dengan fase kumbang
maka seluruh anaknya adalah kumbang.
Gambar 2.1. Kiri: Macan tutul yang mengalami melanisme; kanan: macan tutul
dengan pola warna normal (kanan).
2.1.3 Penyebaran Geografis
Macan tutul memiliki daerah penyebaran yang paling luas di antara jenis kucing
(Guggisberg, 1975; Lekagul & McNeely, 1977). Dari Afrika (melampaui Sahara
Tengah), macan tutul menyebar ke Asia Kecil, Afganistan, Turki, Iran, India, Srilanka,
Jawa, China termasuk China Utara (Manchuria), hingga Amar Ussuri (Grzimek, 1975;
Nowak, 1997; Sanderson, 1972). Ke arah utara macan tutul menyebar ke Rusia Timur
Jauh5.
Di Indonesia, macan tutul jawa (Panthera pardus melas) hanya ditemukan di
Pulau Jawa dan Pulau Kangean (Direktorat PPA, 1978; 1982). Sisa fosil yang
ditemukan menunjukkan umur satu juta tahun (Hemmer & Schutt, 1973). Van Helvoort
et al. (1985) memperkirakan macan tutul diintroduksi ke Pulau Kangean yang letaknya
4 http://www.felidtag.org/pages/Educational/factSheets/leopard.htm 5 http://www.felidtag.org/pages/Educational/FactSheets/leopard.htm
Sumber : UCN - The World Conservation Union (1996)
Sumber : http://globalcrossing.net/∼brendel/leopard.htm (2007)

13
lebih jauh dari Jawa dibandingkan Pulau Bali, di mana macan tutul tidak ada. Macan
tutul tidak terdapat di Sumatera, Kalimantan maupun Bali (Hoogerwerf, 1970).
Seidensticker (1986) berspekulasi bahwa macan tutul (dan harimau) mungkin tidak ada
di Pulau Borneo karena tidak adanya mangsa utama berupa ungulata besar dan macan
tutul tidak ada di Pulau Bali karena adanya harimau Bali dan tidak ada di Sumatra
karena melimpahnya anggota Felidae lainnya (tujuh spesies). Pada tahun 1996
ekspedisi yang dilakukan oleh Konservasi Satwa bagi Kehidupan (KSBK) di Cagar
Alam Pulau Sempu (Kabupaten Malang) menemukan macan kumbang di pulau seluas
877 ha tersebut (Surabaya Post Hot News, Selasa, 17/09/1996).
Sumber : http://commons.wikimedia.org/wiki/Image:Panthera_pardushistoric_distribution.gif
Gambar 2.2. Sejarah penyebaran macan tutul di dunia.
Di Jawa Tengah macan tutul terdapat di Randublatung, Pati, Kendal, Semarang,
Telawa, Gunung Muria dan Gunung Lawu (Hoogerwerf, 1970). Menurut Direktorat
Jenderal PHPA (1987) daerah penyebaran macan tutul di Jawa Tengah dan Daerah
Istimewa Yogyakarta adalah sebagai berikut: Pulau Nusa Kambangan, Batang,
Banjarnegara, Kendal, Cepu, Sragen, Kebasen, Notog, Jatilawang, Gunung Slamet,
Gunung Muria, Gunung Kidul, Gunung Merapi dan Kulon Progo.
Di Jawa Timur macan tutul terdapat di Taman Nasional Meru Betiri, TN.
Baluran, Tuban, Ponorogo, Padangan, Saradan, Jember, Blitar, Jatirogo, Madiun dan
Gundih. Di Jawa Barat macan tutul terdapat di Cianjur Selatan, Gunung Gede, Gunung
Pangrango, Cirebon dan Taman Nasional Ujung Kulon (Hoogerwerf, 1970). Suatu

14
penelitian yang dilakukan oleh tim LIPI, PHPA dan JICA berhasil memotret macan
tutul di TN. Gunung Halimun (Departemen Kehutanan, 1997).
Gunawan (1988) menemukan bukti keberadaan macan tutul berupa feces, jejak
dan bekas cakaran di pohon serta garukan di tanah di Cagar Alam (CA) Pringombo
(Kab. Banjarnegara), hutan jati BKPH Subah (Kab. Batang), Serang (Kab. Purbalingga)
dan CA. Nusa Kambangan Timur (Kab. Cilacap). Sementara Di Gunung Kidul tidak
berhasil diperoleh bukti keberadaan macan tutul.
2.1.4 Habitat
Macan tutul menempati berbagai tipe habitat dengan toleransi yang tinggi
terhadap variasi iklim dan makanan (Guggisberg, 1975; Lekagul & McNeely, 1977).
Macan tutul merupakan spesies yang sangat mudah beradaptasi. Mereka ditemukan di
setiap tipe hutan, savana, padang rumput, semak, setengah gurun, hutan hujan tropis
berawa, pegunungan yang terjal, hutan gugur yang kering, hutan konifer sampai sekitar
pemukiman (Cat Specialist Group, 2002).
Di Asia macan tutul terdapat di hampir semua tipe lingkungan6. Macan tutul
sangat tangguh menghadapi perkembangan pemukiman manusia, akibat meningkatnya
kepadatan populasi manusia di sekitar hampir seluruh habitatnya. Macan tutul masih
ditemukan di seluruh Jawa meskipun dalam jumlah yang sedikit, padahal pulau ini
merupakan salah satu pulau terpadat penduduknya di dunia (IUCN - The World
Conservation Union, 1996).
Macan tutul lebih toleran daripada harimau terhadap temperatur ekstrim dan
lingkungan yang kering (Santiapillai & Ramono, 1992), sebagai contoh, mereka lebih
umum di hutan monsoon tropika yang kering musiman daripada harimau, yang
tergantung pada sumber air permanen (Kleiman & Eisenberg, 1973; Sunquist, 1981;
Johnsingh, 1983; Rabinowitz, 1989).
Di Afrika macan tutul lebih menyukai semak yang tebal di lingkungan berbatu
dan hutan tepi sungai untuk habitat mereka7. Macan tutul sangat menyukai daerah yang
memiliki pohon untuk aktivitas berlindung dan mengintai karena mereka merupakan
pemanjat yang menakjubkan8.
6 http://www.felidtag.org/pages/Educational/factSheets/leopard.htm 7 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/Friedman/habitat.html 8 http://library.thinkquest.org/11234/leopard. html

15
Betina harus memiliki tempat untuk bersarang di dalam home range-nya
(Bailey, 1993). Tempat bersarang biasanya vegetasi tebal atau singkapan batu. Sarang
sangat penting untuk kelangsungan hidup anak-anaknya karena melindung mereka dari
pemangsa9.
2.1.5 Daerah Jelajah dan Teritori
Macan tutul jantan memiliki home range yang sering overlap dengan beberapa
home range betina. Home range macan tutul umumnya terpusat di sekitar badan air di
mana mangsa terkonsentrasi (Seidentsicker and Susan, 1991). Home range macan tutul
jantan lebih besar karena mangsanya biasanya lebih besar daripada mangsa macan tutul
betina (Sunquist, 2001). Ukuran home range macan tutul sangat bervarasi dan sangat
tergantung pada ketersediaan jumlah dan penyebaran satwa mangsa (IUCN - The World
Conservation Union, 1996).
Aspek-aspek seperti pelindung, perburuan serta penyebaran dan kelimpahan
mangsa adalah penting dalam menentukan ukuran jelajah karnivora pada umumnya,
khususnya Felidae. Semua daerah jelajah memiliki sedikitnya satu badan air dan
beberapa lainnya memiliki lebih dari satu badan air, tetapi macan tutul tampaknya tidak
menggunakan sungai baik secara eksklusif ataupun sebagai batas alam jelajahnya10.
Ukuran home range macan tutul jantan berkisar antara 30 – 78 km2 dan betina
23 – 33 km2 di kawasan yang dilindungi (Bailey, 1993). Home range mungkin jauh
lebih besar ketika ketersediaan makanan berkurang. Sebagai contoh, home range
berkisar antara 338 - 478 km2 ditemukan Norton & Lawson (1985) di dataran tinggi.
Bothma and Knight (1997) menemukan bahwa di Kalahari Selatan yang kering dan
miskin mangsa, rata-rata home range macan tutul jantan dewasa adalah 2.182 ± 492
km2 dan betina dewasa 489 ± 293 km2. Sementara di Taman Nasional Royal Chitwan,
Nepal, yang memiliki kepadatan populasi ungulata sangat tinggi, home range macan
tutul betina hanya 6 – 13 km2 dan di Taman Nasional Serengeti dan Tsavo, Afrika
Timur, teritori mereka berkisar antara 11 – 121 km2 (IUCN - The World Conservation
Union, 1996).
Penelitian pada sebuah ranch di Laikipia, Kenya seluas 200 km2 menunjukkan
bahwa macan tutul betina memiliki home range eksklusif rata-rata 14,0 km² dan 9 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2004/bunton/bunton.html#social 10 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/Friedman/socialsystem.html

16
beberapa diantaranya overlap dengan betina dewasa muda. Home range macan tutul
jantan rata-rata 32,8 km² dan tidak overlap antar sesama jantan tetapi overlap dengan
teritori-teritori betina (Mizutani & Jewell, 1998). Macan tutul muda tidak memiliki
home range tetap sampai mendapatkan home range karena yang dewasa mati11.
Macan tutul mempertahankan teritorinya dari individu lain sesama jenis
kelamin. Jantan dan betina menandai tertorinya dengan menyemprotkan urin dan
meninggalkan tanda cakaran pada batang pohon di pinggiran teritori mereka12. Menurut
Grzimek (1975) macan tutul tidak akan keluar dari teritorinya jika makanan cukup
tersedia dan mudah didapat.
2.1.6 Makanan dan Kebiasaan Makan
Menurut Prater (1965) dalam Hoogerwerf (1970), macan tutul akan membunuh
dan makan apa saja yang mudah ditangkapnya. Kebanyakan mangsa macan tutul
adalah satwa yang masih anak-anak (infant/juvenile) atau yang sudah tua karena
biasanya keadaannya lemah dan mudah ditangkap (Grzimek, 1975). Macan tutul lebih
menyukai ungulata dengan berat tubuh 20 sampai 50 kg13, tetapi kadang-kadang
berburu mangsa yang jauh lebih besar14.
Mangsa macan tutul di Jawa antara lain : babi hutan, kijang, rusa, monyet,
landak, lutung dan burung (Direktorat PPA, 1978). Menurut Bartels (1929) dalam
Hoogerwerf (1970) macan tutul memangsa teledu, musang dan owa abu-abu. Grzimek
(1975) menyatakan bahwa satwa-satwa kecil seperti kelinci, binatang pengerat, ikan dan
burung juga dimangsa macan tutul, bahkan juga buah-buahan yang manis.
Menurut Prater (1965) dalam Hoogerwerf (1970) macan tutul memangsa
binatang melata dan ketam, bahkan menurut Schaller (1969) dalam Lekagul & McNeely
(1977), macan tutul juga memangsa serangga. Westra (1931) dalam Hoogerwerf (1970)
menjumpai macan tutul memburu dan memangsa kelelawar. Sementara menurut
Direktorat PPA (1982) macan tutul memangsa penyu laut yang sedang atau baru selesai
bertelur di pantai. Di daerah yang kepadatan mangsanya rendah, macan tutul juga
11 http://www.catsurvivalstrust.org/leopard.htm 12 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_Information.htm 13 http://www.katzenseite.net/infos/habitat.htm 14 http://www.katzenseite.net/infos/habitat.htm

17
memakan landak, trenggiling, burung merak, ayam hutan, monyet di pohon dan anjing
yang tersesat di pinggiran desa15.
Bila di lingkungan hidupnya persediaan makanan sudah sangat berkurang,
macan tutul kadang-kadang masuk ke perkampungan di sekitar hutan dan memangsa
hewan ternak (Direktorat PPA, 1978). Hewan ternak yang sering dimangsa oleh macan
tutul adalah unggas dan kambing (Veevers-Carter, 1978). Seperti kebanyakan predator
besar, macan tutul dapat menjadi satwa kanibal (Hoogerwerf, 1970).
Menurut Goudriaan (1948) dalam Hoogerwerf (1970) macan tutul di Jawa
memakan korbannya mulai dari jantung, hati dan bagian-bagian lunak lainnya. Giginya
mengagumkan dan efisien untuk membunuh; taringnya membuat gigitan mematikan
dan merobek menembus kulit jangat, sementara molar-nya yang setajam pisau cukur
dan lidahnya yang kasar membuat daging cepat tertelan16.
Macan tutul kadang-kadang menyimpan sisa makanannya dengan cara
menutupinya dengan daun, ranting, rumput atau serasah. Sering pula sisa makanannya
disimpan di atas pohon untuk menghindari jangkauan binatang pemakan bangkai (Van
Dooren, 1949 dalam Hoogerwerf, 1970; Grzimek, 1975). Macan tutul mampu
mengangkat mangsa seberat 125 kg (2 – 3 kali beratnya) ke atas pohon setinggi 5,8 m
(Hamilton, 1976).
Menurut Goudriaan (1948) dalam Hoogerwerf (1970), macan tutul kembali ke
tempat penyimpanan sisa makanannya setelah dua atau tiga hari, bahkan kadang-kadang
lebih. Home (1927) dalam Hoogerwerf (1970) mengatakan bahwa macan tutul kembali
ke tempat penyimpanan sisa makanannya setelah lelah dan gagal dalam berburu.
Kebiasaan memakan bangkai yang disimpannya memungkinkan macan tutul mudah
keracunan (Bailey, 1993)
Bailey (1993) menemukan interval rata-rata antara pemangsaan ungulata
berkisar 7 – 13 hari dan konsumsi harian rata-rata macan tutul dewasa jantan adalah 3,5
kg dan betina 2,8 kg. Menurut Hart et al. (1996) komposisi makanan macan tutul terdiri
atas 53,5 % ungulata dan 25,4 % primata dengan rata-rata berat mangsa 24,6 kg.
Menurut Karanth & Melvin (1995) mangsa macan tutul berimbang antara ungulata dan
primata yaitu 89-98 %. Setelah makan, macan tutul biasanya mencari air untuk minum.
Macan tutul dapat bertahan hidup dengan baik pada musim kering yang panjang 15http://www.katzenseite.net/infos/habitat.htm 16 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/ Mammals/Big_5/Leopard/Leopard_Information.htm

18
walaupun hanya minum tiap 2 - 3 hari sekali (Grzimek, 1975). Macan tutul tidak
membutuhkan banyak air karena cairan yang terkandung pada mangsanya sudah cukup
untuknya17.
2.1.7 Kebiasaan dan Perilaku
a. Kebiasaan
Macan tutul termasuk satwa yang gemar mengembara dan kurang bersifat
menetap, tetapi suka kembali ke tempat persembunyiannya semula (Direktorat PPA,
1978). Macan tutul pemalu, cerdik dan berbahaya, khususnya ketika terluka18.
Seperti halnya harimau loreng (Panthera tigris), macan tutul biasanya hidup
menyendiri (soliter), kecuali pada musim kawin dan masa mengasuh anak. Di Jawa
tidak ada peneliti yang menyebutkan adanya kelompok macan tutul yang lebih dari dua
ekor atau seekor induk yang diikuti oleh lebih dari dua ekor anak (Hoogerwerf, 1970).
Walaupun mungkin merupakan pemangsa paling nokturnal, tetapi macan tutul
juga berburu di siang hari19. Di siang hari yang panas, macan tutul berteduh, baik di
pohon, gua maupun naungan batu. Pohon dan batu berfungsi ganda sebagai tempat
yang baik untuk mengamati areal perburuan dan untuk berlindung. Macan tutul
kadang-kadang berjemur matahari pagi20.
Macan tutul membuang kotoran (feces) tanpa disembunyikan, tetapi diletakkan
di tempat-tempat terbuka misalnya di atas batu-batu besar (Medway, 1975). Gunawan
(1988) mendapati kotoran macan tutul di tengah-tengah persimpangan jalan di hutan jati
Perum Perhutani di Subah (Kabupaten Batang) dan batas kawasan Cagar Alam
Pringombo (Kabupaten Banjarnegara).
Macan tutul adalah perenang yang baik tetapi tidak akan berendam dalam air
seperti harimau, bahkan macan tutul menghindari genangan air seperti kucing rumah
yang tidak senang menjadi basah21. Seperti halnya kucing besar lainnya, secara umum
macan tutul menghabiskan waktu sekitar dua per tiga waktu untuk istirahat dan
17 http://www.katzenseite.net/infos/habitat.htm 18 http://www.sa-venues.com/wildlife/wildlife_leopard.htm 19 http://www.dataid.com/junglegallery.htm 20 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big5/Leopard/Leopard_Information.htm 21 http://www.catsurvivalstrust.org/leopard.htm

19
mempelajari lingkungannya (Seidentsicker and Susan, 1991). Sebagian besar waktunya
untuk berbaring di pohon, di atas batu besar atau di sarangnya22.
Macan tutul adalah satwa arboreal, yang berarti mereka makan, tidur, kawin dan
memburu mangsanya dari atas pohon (Alderton, 1998). Mata mereka sangat spesialis
untuk melihat pada malam hari dengan lapisan pemantul di belakang matanya yang
disebut tapetum lucidum yang membuat cahaya melewati mata dua kali, menciptakan
image yang lebih cemerlang bahkan di cahaya yang redup (Kitchener, 1991). Seperti
halnya banyak kucing lainnya, macan tutul menggunakan kumisnya untuk merasakan
jalan mereka ketika melewati semak yang lebat di malam yang gelap (La Brasca,
2007).
b. Perilaku Berburu
Macan tutul merupakan pemburu soliter23. Pada umumnya macan tutul mencari
mangsa pada senja hingga malam hari, jarang mereka berburu pada siang hari (Grzimek,
1975). Menurut Goudriaan (1948) dalam Hoogerwerf (1970), waktu aktif macan tutul
mengadakan perburuan adalah antara pukul 15.00 sampai 20.00 dan antara pukul 03.00
sampai 06.00, jadi tidak selalu dalam keadaan gelap. Tidak ada kecenderungan yang
kuat baik pada aktivitas nokturnal maupun diurnal (Rabinowitz, 1989). Di Afrika macan
tutul berburu pada siang hari untuk menghindari kompetisi dengan singa dan hyena
(Guggisberg, 1975; Leyhausen & Tonkin, 1979). Dalam beberapa kasus, macan tutul di
Pulau Jawa juga berburu pada siang hari.
Macan tutul adalah pemburu dan penyergap yang berburu dengan indera
penglihatannya, suaranya dan penciumannya. Macan tutul mengincar atau mengintai
mangsanya dari atas pohon atau dari balik semak-semak (Direktorat PPA, 1978).
Ketika mengintai, macan tutul merundukkan badannya ke tanah dan ekornya horisontal,
sementara matanya melokalisir mangsanya menggunakan penglihatan malamnya yang
tajam, berdiam jika mangsanya menengok ke sekitar karena curiga; kemudian macan
tutul menyergap dengan tepat dan cepat24.
22 http://www.catsurvivalstrust.org/leopard.htm 23 http://library.thinkquest.org/11234/leopard.html 24 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_Information.htm

20
Macan tutul memburu mangsanya pada jarak pendek (umumnya kurang dari 30
m) dengan meloncat mangsanya disergap dan diterkam bagian tengkuknya25. Jika
mangsa tertangkap, lehernya digigit dan moncongnya dicakar dengan kaki depan serta
diserangnya sampai mangsa tidak berdaya (Direktorat PPA, 1978). Mangsa dibunuh
dengan mencekik atau menggigit bagian belakang kepala sehingga memutuskan saluran
syaraf tulang belakang26.
c. Perilaku Berkomunikasi
Macan tutul umumnya pendiam. Karakteristik suaranya paling banyak adalah
suara geraman parau, batuk serak berulang-ulang dalam interval, yang mirip dengan
suara gergaji mesin (chainsaw). Panggilan serak biasanya dikeluarkan oleh macan tutul
jantan untuk mengumumkan teritorinya yang akan dibalas oleh macan tutul lainnya, jika
ada individu lain di sekitarnya maka akan terus berulang-ulang mengeluarkan suara
tersebut sampai individu lain itu pergi. Macan tutul mempunyai suara individual yang
berbeda dan ini mungkin menguntungkan bagi satwa soliter seperti macan tutul untuk
mengenali satu dengan lainnya dari kejauhan melalui suara seperti juga mereka saling
menghindar satu sama lain. Dua macan tutul jantan teritorial akan selalu saling
menggeram. Macan tutul betina akan memanggil bila sedang oestrus. Macan tutul juga
dikenal mendengkur selama makan27.
Penandaan teritori oleh macan tutul juga merupakan cara yang penting dalam
komunikasi intra spesifik28. Batas-batas teritori secara teratur ditandai dengan urin,
feces, kemunculan/kehadiran, cakaran di tanah dan pohon29.
d. Perilaku Sosial
Sistem sosial merupakan cara adaptasi macan tutul, karena macan tutul
merupakan karnivora berukuran sedang sehingga tidak memiliki banyak pemangsa dan
dapat berburu sendiri dengan efisien. Cara berburu mereka membuat mereka harus
hidup menyendiri tanpa tergantung pada saudara kandung atau induknya untuk
keberhasilan perkembangbiakan. Tekanan seleksi utama yang membentuk sistem sosial
25 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_Information.htm 26 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_Information.htm 27 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_Information.htm 28 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_Information.htm 29 http://www.lioncrusher.com/animal.asp?animal=57

21
macan tutul adalah lingkungan. Keanekaragaman jenis mangsa macan tutul merupakan
penyumbang utama sistem sosial ini. Karena macan tutul tidak tergantung pada satu
jenis sumber makanan, maka jumlah pesaingnya sedikit30.
Dalam sistem sosial, macan tutul jantan mempertahankan teritorinya yang dapat
mencakup teritori dari dua atau tiga macan tutul betina. Macan tutul jantan
mempertahankan teritori dari jantan lain, dan betina mempertahankan teritori dari betina
lain. Tampaknya anak betina membangun teritorinya di dalam teritori induknya,
sementara anak jantan dikeluarkan dari teritori induk jantan sampai membentuk teritori
di luar tempat kelahirannya31.
Karena macan tutul soliter, pertemuan para dewasa dari jenis kelamin yang sama
umumnya jarang. Macan tutul dewasa dari jenis kelamin yang sama dan memiliki
daerah jelajah bersebelahan atau overlap biasanya saling menghindar, tetapi perkelahian
bisa terjadi khususnya untuk memperebutkan mangsa32.
Interaksi antara macan tutul dan spesies pemangsa besar lainnya sangat
kompleks (Bertram, 1982). Macan tutul cenderung menghindari daerah kekuasaan
harimau33. Keberadaan pohon atau batu sebagai tempat menyelamatkan diri
memungkinkan macan tutul dapat hidup bersama (co-exist) dengan pesaing-pesaing
besarnya. Macan tutul dapat masuk dalam teritori singa yang kosong (Bertram 1982).
Dimana ada harimau, macan tutul cenderung sedikit (Schaller, 1967; 1972; M.K.
Ranjitsinh pers. comm. dalam La Brasca, 2007), tetapi ini bukan aturan yang baku. Di
Taman Nasional Chitwan, Nepal, macan tutul dan harimau co-exist dengan cara berburu
pada waktu yang berbeda dan mangsa yang berbeda serta menggunakan komplek
vegetasi yang berbeda (Seidensticker, 1976). Macan tutul makan mangsa yang lebih
kecil (biasanya kurang dari 75 kg) (Seidensticker, 1976; Johnsingh, 1983), pembagian
mangsa juga terjadi antara singa dan macan tutul di Serengeti (Bertram, 1982) dan Gir
Forest (R. Chellam in litt., 1993 dalam La Brasca, 2007).
30 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2004/bunton/bunton.html#social 31 http://www.travelafricamag.com/content/view/176/56 32 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2004/bunton/bunton.html#social 33 http://www.catsurvivalstrust.org/leopard.htm

22
2.1.8 Perkembangbiakan dan perilaku berkembangbiak
a. Sistem Perkawinan dan Perilaku Kawin
Sistem perkawinan macan tutul adalah promiscuity yaitu jantan dan betina kawin
dengan lebih dari satu pasangan dan tidak ada ikatan jangka panjang34. Betina akan
kawin dengan jantan-jantan yang lebih tua yang memiliki home range overlap
dengannya. Sistem perkawinan ini berkembang karena betina dapat membesarkan
anak-anaknya tanpa bantuan dan perlindungan jantan35.
Macan tutul tidak memiliki musim berkembang biak khusus36. Jika musim
memungkinkan, macan tutul akan kawin sepanjang tahun (Guggisberg, 1975;
Kithchener, 1991; Nowak, 1997), tetapi puncaknya selama musim kelahiran mangsa
utamanya 37.
Seekor betina mengalami oestrus rata-rata tujuh hari (4 – 14 hari)38 dengan
siklus sekitar 46 hari39. Betina yang oestrus akan menarik perhatian jantan dangan
memanggil, dan akan meninggalkan tanda bau pada pohon atau semak-semak, ia juga
menjadi sering mengembara keluar dari home range-nya. Jantan dan betina membentuk
asosiasi sementara, dan seekor betina oestrus mungkin dikawini oleh beberapa jantan
dalam rentang waktu yang singkat40.
Jantan memiliki ritual perkawinan dengan betina-betina yang memiliki teritori
overlap dengan teritorinya (Estes, 1991). Jantan akan mengikuti betina yang birahi dan
berkelahi dengan jantan lain untuk mendapatkan hak kawin (Estes, 1991; Nowak,
1997). Macan tutul betina siap dikawini dalam interval 3-7 minggu dan periode ini
berlanjut sampai beberapa hari di mana terjadi perkawinan41.
Seekor betina mungkin dirayu oleh beberapa jantan. Jantan yang berhasil akan
menggigit bagian belakang leher betina dengan giginya, betina akan menampar jantan
ketika kopulasi telah sempurna. Kopulasi sangat sering, dari 70 sampai 100 kali
sehari42. Laman & Cheryl (1997) yang mengamati perilaku kawin macan tutul di
Taman Nasional Serengeti, Tanzania menemukan kopulasi sebanyak 13 kali selama satu 34 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html 35 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2004/bunton/bunton.html#social 36 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_ Information.htm 37 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html 38 http://www.catsurvivalstrust.org/leopard.htm 39 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html 40 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_ Information.htm 41 http://en.wikipedia.org/wiki/Indian_Leopard 42 http://www.catsurvivalstrust.org/leopard.htm

23
setengah jam pengamatan. Semua kopulasi tercatat dan semua dimulai dengan betina
berjalan mundur dan maju di depan jantan yang sedang beristirahat, menggosokkan
badannya dan menggoyang-goyangkan ekornya di wajah sang jantan. Jantan seringkali
menggigit betina di bagian tengkuknya selama interaksi ini. Perkawinan disertai dengan
suara geraman, baik dari jantan maupun betina. Puncaknya berlangsung rata-rata tiga
detik dengan interval rata-rata antara kopulasi 6,5 menit. Dalam kandang, kopulasi
tercatat 100 kali sehari (Kitichner, 1991) dan kopulasi yang terlihat bisa jadi merupakan
bagian dari percumbuan43.
Rata-rata lamanya waktu jantan dan betina dewasa bersama adalah 2,1 hari.
Ketika betina dan jantan menghabiskan waktu bersama, mereka akan kawin, jantan akan
mengikuti betina kemanapun pergi dan kadang-kadang mereka berbagi mangsa
buruan44. Perkawinan berlangsung selama dua atau tiga hari. Interval dengan
pekawinan berikutnya dua tahun45. Setelah musim kawin berakhir, jantan dan betina
akan berpisah46.
b. Masa Bunting dan Jumlah Anak Per Kelahiran
Macan tutul betina akan mencapai kematangan seksual pada umur rata-rata 33
bulan (30 – 36 bulan)47. Seekor betina, pertama melahirkan pada umur 2,5 tahun
sampai 3 tahun48. Masa kehamilan rata-rata 96 hari (90 – 112 hari)49. Laporan lain
menyebutkan masa kebuntingan macan tutul 3 - 5 bulan50.
Induk yang bunting mencari gua, celah batu besar, lubang pohon atau semak
belukar untuk melahirkan dan membuat sarang (Nowak, 1997; Guggisberg, 1975).
Anak-anak macan tutul dilahirkan dalam gua, lobang pohon, lubang tanah atau tempat
berlindung lain yang sesuai51. Macan tutul umumnya melahirkan dua anak per
kelahiran, kadang-kadang tiga atau empat,52 tetapi juga ada yang melaporkan sampai
43 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html 44 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2004/bunton/bunton.html#social 45 http://www.travelafricamag.com/content/ view/176/56 46 http://www.naturalia.org/ ZOO/AN_TERRA/e_leopardo.html 47 http://www.catsurvivalstrust.org/leopard.htm 48 http://en.wikipedia.org/wiki/Indian_Leopard 49 http://www.catsurvivalstrust.org/leopard.htm 50 http://www.naturalia.org/ ZOO/AN_TERRA/e_leopardo.html 51 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_
Information.htm 52 http://en.wikipedia.org/wiki/Indian_Leopard

24
enam ekor setiap kelahiran53. Rasio kelamin dalam setiap kelahiran adalah satu banding
satu54.
Angka kematian bayi tinggi sehingga hanya tersisa satu atau dua anak
(Guggisberg, 1975; Kithchener, 1991; Nowak, 1997). Menurut Garman (1997) angka
kematian anak macan tutul 40-50%, sehingga biasanya jarang dijumpai induk bersama
anak lebih dari 1 - 2 ekor55.
Anak-anak macan tutul lahir dalam keadaan mata tertutup dan lemah dengan
berat kurang dari dua pound56 atau sekitar 0,5 kg57. Anak macan tutul yang baru
dilahirkan berwarna keabu-abuan karena tutul rosette-nya belum tampak jelas58.
Rambutnya lebih panjang dan lebih lembut dibandingkan yang dewasa, warna kulitnya
agak abu-abu pucat dengan tutul-tutul kecil yang kurang jelas59. Mata anak macan tutul
terbuka setelah sekitar enam hari60 sampai 10 hari61. Anak-anak dapat berjalan setelah
13 hari62. Macan tutul beranak lagi setelah 15 bulan (jika anak-anaknya mati) sampai
lebih dua tahun. Rata-rata umur perkembangbiakan terakhir adalah 8,5 tahun63.
c. Pemeliharaan Anak
Macan tutul betina membesarkan anak-anaknya di tempat bersarang dan
mengajarinya berburu64. Mereka disembunyikan sekitar delapan minggu dan induknya
akan berhenti berpindah-pindah sampai anak-anak dapat bergabung mengikuti
perjalanan65.
Anak macan tutul disusui induknya selama tiga bulan atau lebih66. Mereka
dibimbing makan daging sampai kira-kira umur 10 bulan, ketika mereka begabung
dengan induk mereka pada perburuan67.
53 http://www.catsurvivalstrust.org//leopard.htm 54 http://www.travelafricamag.com/content/view/176/56 55 http://en.wikipedia.org/wiki/Leopard.htm 56 http://library.thinkquest.org/11234/leopard.html 57 http://www.catsurvivalstrust.org/leopard.htm 58 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html 59 http://www.catsurvivalstrust.org/leopard.htm 60 http://www.travelafricamag.com/content/view/176/56 61 http://www.catsurvivalstrust.org/leopard.htm 62 http://www.catsurvivalstrust.org/leopard.htm 63 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html 64 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2004/bunton/bunton.html#social 65 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html 66 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html

25
Anak-anak macan tutul disapih setelah tiga bulan dan mulai bergabung dengan
induknya berburu di mana mereka akan belajar untuk bertahan hidup68. Pada umur lima
bulan mereka dapat menangkap hewan kecil dan kebanyakan dapat mencari makan
sendiri pada umur satu tahun69.
Anak-anak macan tutul belajar dengan meniru tingkah laku induknya, dan
mereka biasanya membunuh mangsa utama pertamanya pada umur 11 bulan, walaupun
mereka dapat membunuh hewan kecil seperti garangan atau bangsa pengerat pada umur
sekitar empat bulan. Induknya mungkin membawa mangsa hidup atau mati untuk anak-
anaknya, di mana mereka belajar menerkamnya dan belajar menguasainya dengan
cakarnya70.
Betina mungkin pergi meninggalkan anak-anak untuk waktu lama, kadang-
kadang meninggalkan mereka sampai satu setengah hari sendirian71. Induk macan tutul
memindahkan anak-anaknya ke tempat berlindung baru setiap dua atau tiga hari72, cara
membawanya dengan menggigit tengkuknya dan bahkan bisa membawanya sambil
berenang73.
Ketika berpindah dengan anak-anaknya, ekor induk macan tutul melengkung
ke atas, menunjukkan warna keputihan bagian bawah ekor yang mungkin berperan
sebagai petunjuk bagi anak-anaknya di antara rumput yang tinggi. Pemangsaan pada
anak-anak macan tutul, khususnya yang dilakukan oleh macan tutul lain sangat umum
terjadi sehingga jarang lebih dari satu atau dua anak yang selamat dan dapat bertahan
hidup74.
Pada umur satu tahun, macan tutul muda mungkin dapat menjaga dirinya sendiri
tetapi masih bersama induknya sampai umur 18 – 24 bulan (Nowak, 1997; Guggisberg,
1975), ketika mulai perkembangbiakan berikutnya75. Setelah umur dua tahun menjadi
67 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_
Information.htm 68 http://library.thinkquest.org/11234/leopard.html 69 http://www.catsurvivalstrust.org/leopard.htm 70 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_
Information.htm 71 http://www.catsurvivalstrust.org/leopard.htm 72 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_
Information.htm 73 http://www.catsurvivalstrust.org/leopard.htm 74 http://www.ecotravel.co.za/Guides/Wildlife/Vertebrates/Mammals/Big_5/Leopard/Leopard_
Information.htm 75 http://en.wikipedia.org/wiki/Indian_Leopard

26
dewasa muda (sub-adult) anak-anak macan tutul akan meninggalkan induknya untuk
membangun teritori mereka sendiri76. Anak-anak menjadi dewasa secara seksual sekitar
dua setengah tahun77.
Pemencaran mungkin tertunda di wilayah yang melimpah mangsanya,
khususnya jika macan tutul lain menghuni habitat di sekitarnya78. Sistem pemencaran
macan tutul muda tampaknya fleksibel dan anak yang masih muda tidak selalu
meninggalkan tempat kelahirannya ketika sudah bisa mandiri. Tetapi sebaliknya
beberapa jantan muda meninggalkan tempat kelahirannya pada umur 15 – 16 bulan
(Seidensticker and Susan, 1991).
Macan tutul jantan tidak ambil bagian dalam membesarkan anak dan hanya
bertemu dengan betina untuk kawin79. Guggisberg (1975) melaporkan bahwa jantan
juga membantu betina yang mengasuh anak-anak seperti dengan memberi mereka hasil
buruan. Induk jantan mungkin dijauhkan dari anak-anak oleh betina, tetapi jantan
memberikan daging kepada anak-anaknya80.
2.1.9 Masa Hidup
Di berbagai negara macan tutul yang hidup dalam kandang dapat mencapai
umur 21 – 23 tahun. Di alam, umurnya tidak diketahui, tetapi diperkirakan antara tujuh
dan sembilan tahun (Guggisberg, 1975). Menurut Garman (1997) masa hidup macan
tutul antara 12-17 tahun.
2.2 Habitat, Relung, Daerah Jelajah dan Teritori
2.2.1 Habitat
Habitat adalah suatu tipe komunitas biotik atau kesatuan komunitas biotik di
mana seekor satwa atau populasi hidup (Bailey, 1984). Habitat adalah suatu unit
lingkungan, alami maupun tidak (meliputi iklim, makanan, cover dan air) dimana seekor
satwa, tumbuhan atau populasi secara alami dan normal hidup dan berkembang (Helms,
1998). Definisi habitat terbaru yang relevan untuk pengelola satwa liar datang dari Hall
et al. (1997) yaitu sumberdaya dan kondisi yang ada pada suatu tempat yang
76 http://library.thinkquest.org/11234/leopard.html 77 http://www.travelafricamag.com/content/view/176/56 78 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/friedman/Matingsystem.html 79 http://www.bio.davidson.edu/people/vecase/Behavior/Spring2004/bunton/bunton.html#social 80 http://www.catsurvivalstrust.org/leopard.htm

27
memberikan tempat hidup (occupancy), termasuk survival dan reproduksi suatu
organisme. Definisi ini berimplikasi bahwa habitat adalah sejumlah sumberdaya
spesifik yang dibutuhkan oleh suatu spesies (Hall et al., 1997).
Semua jenis satwa dapat hidup di suatu tempat hanya jika kebutuhan pokoknya
seperti makanan, air, dan cover tersedia dan jika satwa memiliki daya adaptasi yang
memungkinkannya menghadapi iklim yang ekstrim, kompetitor dan predator (Morrison
et al., 1992). Empat komponen dasar habitat adalah makanan, cover, air dan ruang
(Shaw, 1985).
Komponen habitat paling penting bagi satwa adalah makanan. Ketersediaan
(availability) makanan biasanya berubah menurut musim. Bagi karnivora atau jenis
pemangsa, ketersediaan makanan berarti ketersediaan satwa mangsa (Shaw, 1985).
Kuantitas dan kualitas makanan yang dibutuhkan oleh setiap satwaliar bervariasi
menurut spesies, jenis kelamin, kelas umur, fungsi fisiologis, musim, cuaca dan lokasi
geografis (Bailey, 1984). Karnivora mengeluarkan banyak energi untuk mencari,
memburu, menangkap dan membunuh mangsa, tetapi diimbangi dengan kandungan
energi yang tinggi dari satwa mangsanya. Karena kandungan nutrisi daging mangsa
yang lengkap dan mudah dicerna, spesies pemangsa jarang atau tidak pernah mengalami
kekurangan gizi dari mangsa alaminya (Shaw, 1985). Masalah nutrisi bagi karnivora
adalah masalah kuantitas dan ketersediaan (availability), bukan kualitas makanan
(Bailey, 1984).
Cover didefinisikan sebagai sumberdaya struktural dari lingkungan yang
mendukung perkembangbiakan (reproduksi) dan/atau daya hidup (survival) satwa
dengan menyediakan fungsi-fungsi alami untuk spesies tersebut (Bailey, 1984). Cover
biasanya digunakan untuk melarikan diri dari predator, walaupun predator yang
memburu mangsanya juga memerlukan cover untuk dapat mendekati mangsanya.
Cover juga memberikan perlindungan yang penting terhadap iklim yang keras, tempat
berteduh dari panas, angin dan hujan atau perlindungan dari udara malam yang dingin
(Shaw, 1985).
Vegetasi bukan satu-satunya komponen struktural dari lingkungan yang
mempengaruhi satwaliar. Badan air yang besar mungkin penting bagi itik penyelam,
tebing penting bagi kambing gunung, lereng ke arah selatan penting bagi rusa karena
lebih hangat dan makanannya tidak tertutup salju (Bailey, 1984). Cover dari vegetasi

28
seringkali lebih penting strukturnya daripada jenisnya. Sebagai pelindung, cover
mungkin lebih memberikan kesejahteraan satwa melalui fungsi-fungsi alaminya sebagai
tempat berkembang biak, makan, perjalanan, melarikan diri, bersarang dan beristirahat
(Bailey, 1984). Komponen struktural dari cover yang penting mungkin bentuk vegetasi
(rumput, semak, perdu, pohon), kerapatan vegetasi, kedalaman air, topografi, lereng dan
lain-lain. Komponen mikro-klimat yang penting mungkin temperatur, kelembaban,
angin, intensitas cahaya dan lain-lain (Bailey, 1984).
Air merupakan komponen habitat yang dibutuhkan dalam banyak proses kimia
dan fisik di dalam tubuh satwa. Air juga digunakan untuk pendinginan melalui
evaporasi di lingkungan yang panas (Bailey, 1984). Kebanyakan satwa memenuhi
kebutuhan airnya dengan minum dari air permukaan. Air dapat mempengaruhi satwa
secara tidak langsung melalui perubahan di dalam habitat (Shaw, 1985). Respon satwa
terhadap kelangkaan air ada tiga macam, menggali dasar sungai (seperti dilakukan
gajah), migrasi ke sumber air dan meninggalkan daerah jelajahnya yang kekeringan
selama musim kering dan berkumpul di sekitar sumber air. Hal ini dapat
menguntungkan bagi satwa predator tetapi juga dapat menjadi media penularan penyakit
dan parasit (Bailey, 1984).
Satwaliar secara individu membutuhkan berbagai ukuran ruang untuk
mendapatkan makanan, cover dan air dengan cukup serta untuk menemukan
pasangannya. Populasi satwaliar membutuhkan ruang yang lebih banyak. Ukuran luas
yang dibutuhkan oleh suatu spesies tergantung pada ukuran satwa (biasanya semakin
besar satwa, membutuhkan ruang semakin luas), makanan (karnivora membutuhkan
ruang lebih luas daripada herbivora) dan produktivitas serta keanekaragaman habitat
berkaitan dengan kebutuhan habitat dari spesies tersebut (Shaw, 1985).
2.2.2 Relung (Niche)
Satu hal terpenting dari konsep ekologi adalah relung (niche) atau peran yang
dimainkan oleh setiap spesies dalam habitat alaminya. Karena relung lebih berbicara
peran daripada tempat, maka hanya dapat digambarkan dengan interaksi antara spesies
dan lingkungannya. Bagian paling penting dari relung adalah pemisahan makanan,
walaupun relung lain juga penting seperti cara penggunaan cover, air, atau bahkan ruang
(Shaw, 1985).

29
Konsep relung ekologi berkaitan dengan evolusi, adaptasi dan pembatasan. Ahli
ekologi umumnya menggunakan dua definisi relung, satu lebih menekankan pada fungsi
satwa, yang kedua menekankan pada sumberdaya habitat. Secara fungsional, relung
ekologi adalah peran suatu spesies dalam komunitas biotiknya seperti yang dibedakan
oleh penyebaran geografis dan ekologis serta oleh serangkaian adaptasi yang
memisahkannya dari semua spesies lainnya. Fungsi makan biasanya menjadi pokok
bahasan dalam konsep ini. Dalam karnivora terdapat spesialis pemangsa mamalia
besar, mamalia kecil, burung, serangga dan sebagainya (Bailey, 1984).
Suatu relung ekologi adalah sekumpulan sumberdaya habitat (makanan, tipe
cover, air, dan lain-lain) yang digunakan oleh suatu spesies yang ditentukan oleh
wilayah geografis, ekologis dan adaptasinya. Sumberdaya makanan sering menjadi
penekanan dalam konsep niche ini, karena penggunaan makanan seringkali lebih mudah
diukur dibandingkan penggunaan sumberdaya habitat lainnya. Relung makan (feeding
niche) digunakan untuk membatasi definisi pada sumberdaya makanan (Bailey, 1984).
2.2.3 Daerah Jelajah dan Teritori
Daerah jelajah (home range) adalah daerah yang digunakan oleh individu satwa
untuk mendapatkan makanan, pasangan dan memelihara anak (Burt, 1943 dalam Shaw,
1985). Suatu home range adalah daerah yang dijelajahi oleh seekor satwa atau populasi
dalam aktivitas normal hariannya. Seekor satwa harus menemukan semua kebutuhan
habitatnya di dalam home range-nya, jika tidak, ia akan memperluas home range-nya.
Satwa penetap, mungkin hanya memiliki satu home range. Beberapa spesies biasanya
memiliki beberapa home range yang digunakan secara musiman. Rute perjalanan atau
migrasi antara home range bukan bagian dari home range karena pergerakan sepanjang
rute ini bukan aktivitas harian (Bailey, 1984).
Ukuran home range bervariasi di antara jenis satwa (Sanderson, 1966 dalam
Bailey, 1984). Umumnya karnivora memiliki home range lebih besar daripada
herbivora walaupun ukuran tubuhnya sama. Home range jantan sering kali lebih besar
daripada betina spesies yang sama. Home range dapat mengecil pada beberapa musim,
seperti masa mengasuh anak atau musim salju. Home range di habitat yang baik (kaya)
akan lebih kecil daripada di habitat yang buruk (miskin). Home range populasi dengan

30
kepadatan tinggi lebih kecil karena mereka menempati habitat yang baik dan karena
interaksi sosial mereka mungkin membatasi pergerakan (Bailey, 1984).
Teritori (territory) adalah bagian atau keseluruhan dari suatu home range yang
dipertahankan dari satwa lain, khususnya dari spesies yang sama. Teritori mungkin
dipertahanakan secara individual, pasangan yang sedang berkembang biak, atau
kelompok sosial sepanjang tahun atau secara musiman. Sifat teritorial penting dalam
pengaturan populasi pada beberapa spesies (Bailey, 1984). Schoener (1968) dalam
Shaw (1985) mendefinisikan secara sederhana bahwa teritori adalah daerah ekslusif,
bisa secara spasial seperti pada banyak burung atau temporal seperti pada banyak
mamalia karnivora
2.3 Teori Biogeografi Pulau
Teori Biogeografi pulau pertama dikemukakan oleh MacArthur & Wilson
(1967). Dalam teorinya mereka berusaha memprediksi jumlah spesies yang mungkin
akan bertahan pada suatu pulau yang baru tercipta. Dalam biogeografi pulau dipelajari
dan dijelaskan faktor-faktor yang mempengaruhi keragaman spesies dari suatu
komunitas tertentu. Dalam konteks ini, “pulau” dapat berupa areal habitat yang
dikelilingi oleh areal lain yang tidak sesuai untuk spesies dalam “pulau” tersebut; bukan
hanya pulau sesungguhnya yang dikelilingi lautan, tetapi juga gunung yang dikelilingi
oleh gurun pasir, danau yang dikelilingi daratan, dan fragment hutan yang dikelilingi
oleh lansekap yang terganggu oleh manusia.
Menurut teori biogeografi, jumlah spesis (jumlah equilibrium) yang terdapat di
suatu pulau ditentukan oleh dua faktor, yaitu jarak dari daratan utama dan ukuran pulau.
Keduanya akan mempengaruhi laju kepunahan di pulau dan tingkat imigrasi. Pulau-
pulau yang dekat dengan daratan utama kemungkinan menerima imigran dari daratan
utama lebih besar daripada pulau-pulau yang jauh dari daratan utama. Pada pulau-pulau
yang lebih kecil peluang kepunahan lebih besar daripada pulau-pulau besar. Pulau-
pulau besar memiliki jumlah spesies lebih banyak daripada pulau-pulau kecil
(MacArhtur & Wilson, 1967).
Menurut teori biogeografi pulau (MacArthur & Wilson, 1967), kekayaan spesies
suatu pulau tergantung pada:

31
a. Isolasi pulau, karena isolasi mempengaruhi laju kolonisasi. Pulau yang terisolasi
atau jauh, memiliki spesies yang lebih sedikit daripada pulau yang dekat dengan
sumber spesies yang mengkolonisasi. Pulau yang lebih jauh, lebih sedikit didatangi
pengkoloni dibandingkan pulau yang lebih dekat. Jika ada pulau-pulau di antara
sumber kolonisasi (daratan utama) dengan pulau, maka dapat berperan sebagai batu
loncatan (stepping stones) dan dapat meningkatkan laju kolonisasi pulau yang jauh.
Jika ada dua pulau dengan jarak yang sama dari sumber kolonisasi, maka pulau yang
lebih besar akan memiliki laju kolonisasi yang lebih tinggi karena adanya Target
Effect (The bigger targets are easier to hit).
b. Luas pulau, karena luas pulau mempengaruhi laju kepunahan. Pulau yang besar
memiliki jumlah spesies yang lebih banyak dibandingkan pulau yang kecil. Pulau
kecil memiliki ukuran populasi yang lebih kecil, lebih sedikit refugia dan memiliki
laju kepunahan lebih tinggi.
c. Dinamika kolonisasi dan kepunahan, kolonisasi menggantikan spesies yang
punah (species turnover).
d. Kekayaan spesies mencerminkan suatu kesetimbangan (equilibrium) antara
kolonisasi dan kepunahan.
Hubungan antara jumlah spesies dan luas pulau digambarkan dengan rumus
sebagai berikut: zcAS = ..................................................................... (Formula 2.1)
Dimana S = jumlah spesies, A = luas pulau, z dan c adalah konstanta yang diperlukan
untuk menyesuaikan data luas (dalam m2, km2, dll) dengan jumlah spesies. Hubungan
ini dapat dibuat linier dengan menggunakan log sehingga rumusnya menjadi :
zLogALogcLogS += ................................................ (Formula 2.2)
Laju kolonisasi lebih tinggi pada pulau yang dekat dengan daratan utama karena
lebih banyak spesies yang dapat menyeberang laut yang relatif dekat. Laju kepunahan
lebih besar pada pulau yang lebih kecil karena populasi-populasi berukuran lebih kecil
dan kemungkinan terkena penyakit dan kejadian merugikan lainnya yang lebih besar
yang dapat menghabiskan populasi atau menurunkannya sampai tingkat yang tidak
32
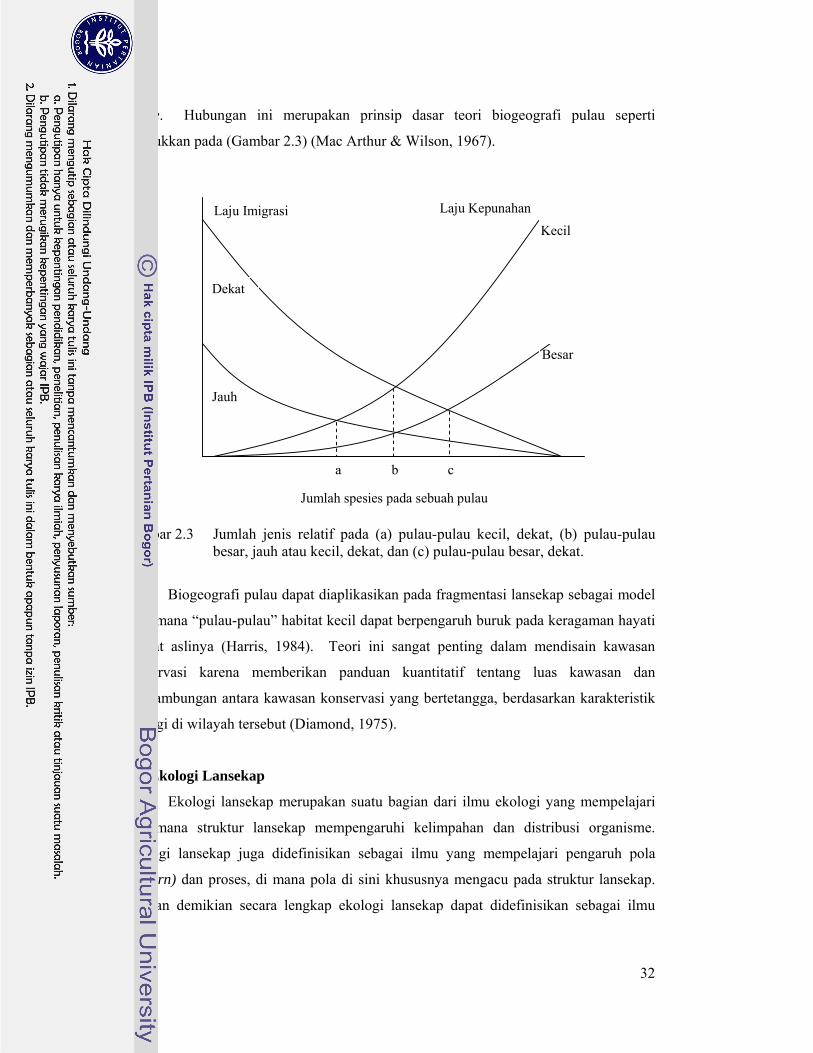
viable. Hubungan ini merupakan prinsip dasar teori biogeografi pulau seperti
dtunjukkan pada (Gambar 2.3) (Mac Arthur & Wilson, 1967).
Gambar 2.3 Jumlah jenis relatif pada (a) pulau-pulau kecil, dekat, (b) pulau-pulau
besar, jauh atau kecil, dekat, dan (c) pulau-pulau besar, dekat.
Biogeografi pulau dapat diaplikasikan pada fragmentasi lansekap sebagai model
bagaimana “pulau-pulau” habitat kecil dapat berpengaruh buruk pada keragaman hayati
habitat aslinya (Harris, 1984). Teori ini sangat penting dalam mendisain kawasan
konservasi karena memberikan panduan kuantitatif tentang luas kawasan dan
kesinambungan antara kawasan konservasi yang bertetangga, berdasarkan karakteristik
ekologi di wilayah tersebut (Diamond, 1975).
2.4 Ekologi Lansekap
Ekologi lansekap merupakan suatu bagian dari ilmu ekologi yang mempelajari
bagaimana struktur lansekap mempengaruhi kelimpahan dan distribusi organisme.
Ekologi lansekap juga didefinisikan sebagai ilmu yang mempelajari pengaruh pola
(pattern) dan proses, di mana pola di sini khususnya mengacu pada struktur lansekap.
Dengan demikian secara lengkap ekologi lansekap dapat didefinisikan sebagai ilmu
Laju Imigrasi Laju Kepunahan
Jumlah spesies pada sebuah pulau
Jauh
Dekat
Kecil
Besar
a b c

33
yang mempelajari bagaimana struktur lansekap mempengaruhi (memproses dan
membentuk) kelimpahan dan distribusi organisme81.
Ekologi lansekap menekanankan dinamika heterogenitas spasial-temporal dan
pengaruhnya pada proses-proses biotik dan abiotik pada lansekap (Forman & Godron,
1986; Turner & Gardner, 1991). Paradigma dinamika metapopulasi menghubungkan
proses populasi dengan karakteristik spasial lansekap dan menjadi bagian integral dari
ekologi lansekap (Singleton et al., 2002).
Teori biogeografi pulau dari MacArtur dan Wilson merupakan fokus dari
ekologi lansekap yang oleh Levin digunakan untuk menjelaskan model metapopulasi.
Dalam ekologi lansekap dapat dipelajari bagaimana fragmentasi habitat mempengaruhi
daya hidup suatu populasi (population viability). Dalam perkembangannya, ekologi
lansekap banyak menggunakan teknologi Sistem Informasi Geografis (SIG) dan banyak
data habitat yang tersedia (seperti citra satelit dan foto udara)82.
Ada tiga pendekatan pada ekologi spasial, pertama teori ekologi yang
mengasumsikan ruang homogen kontinu atau deskrit, kedua ekologi lansekap yang
bertujuan menganalisis struktur lansekap sesungguhnya yang kompleks dengan sedikit
penekanan pada pemodelan dinamika populasi dan ketiga ekologi metapopulasi yang
berada di tengah-tengah, membuat penyederhanaan asumsi bahwa habitat yang cocok
(suitable) untuk spesies target terjadi sebagai jejaring (network) dari kantong-kantong
habitat (habitat patches) yang ideal dengan luas, derajat isolasi dan kualitas yang
bervariasi serta tenggelam di tengah-tengah habitat yang tidak cocok (unsuitable) yang
seragam (Hanski, 1998).
2.4.1 Matrix
Matrix merupakan latar belakang (background) sistem ekologi dari suatu
lansekap dengan derajat konektivitas yang tinggi. Konektivitas adalah ukuran
bagaimana suatu koridor, jaringan (network) atau matrix terhubungkan atau
berkesinambungan (Forman, 1995). Sebagai contoh, suatu lansekap berhutan (matrix)
dengan lebih sedikit gap dalam tutupan hutan (patch terbuka) akan memiliki
konektivitas lebih tinggi. Koridor memiliki fungsi penting sebagai jalur (strip) dari
suatu tipe khusus dalam lansekap yang berbeda dengan tetangga di kedua sisinya 81 http://www.carleton.ca/l ands-ecol/whatisle.html 82 http://en.wikipedia.org/wiki/Landscape.ecology

34
(Forman, 1995). Suatu jaringan (network) merupakan suatu sistem hubungan antar
koridor, sementara mosaik menggambarkan pola (pattern) dari patch, koridor dan
matrix yang membentuk suatu lansekap dalam satu kesatuan (Forman, 1995).
2.4.2 Patch
Patch, merupakan suatu terminologi dasar dalam ekologi lansekap didefinisikan
sebagai sebuah area yang relatif homogen yang berbeda dengan sekelilingnya (Forman,
1995). Patch merupakan unit dasar dari lansekap yang berubah dan berfluktuasi, suatu
proses yang disebut dinamika patch. Patch memiliki bentuk tertentu dan konfigurasi
spasial, serta dapat digambarkan secara komposisi oleh variabel-variabel internalnya
seperti jumlah pohon, jumlah jenis pohon, tinggi pohon, atau ukuran-ukuran serupa
lainnya (Forman, 1995).
2.4.3 Edge
Patch bisa memiliki batas (boundary) yang jelas atau tidak jelas (kabur)
(Sanderson & Harris, 2000). Suatu zona yang tersusun atas edge-edge dari ekosistem
yang berdekatan (berbatasan) adalah boundary (Forman, 1995). Edge berarti bagian dari
suatu ekosistem yang berdekatan dengan garis kelilingnya (perimeter), di mana
pengaruh-pengaruh dari patch yang berdekatan dapat menyebabkan perbedaan
lingkungan antara interior suatu patch dengan tepiannya (edge). Efek tepi (edge effect)
ini meliputi pebedaan komposisi spesies atau kelimpahan di bagian luar patch (Forman,
1995). Sebagai contoh, ketika suatu lansekap merupakan sebuah mosaik dari tipe-tipe
yang jelas, seperti suatu hutan berdekatan dengan suatu padang rumput, edge-nya
adalah lokasi di mana kedua tipe tersebut bergabung. Dalam lansekap yang kontinyu,
seperti suatu hutan ke daerah berkayu yang terbuka, lokasi edge pastinya kabur dan
kadang-kadang dibedakan oleh gradien lokal melampaui suatu ambang batas, seperti
halnya suatu titik dimana penutupan pohon menurun di bawah 35 persen (Turner &
Gardner, 1991).
2.4.4 Koridor
Koridor merupakan komponen lansekap berbentuk strip atau jalur lahan yang
berbeda dengan matrix di sekitarnya. Koridor merupakan areal yang menghubungkan
antar patch-patch sehingga berperan sebagai lintasan atau saluran bagi organisme untuk

35
bertukar atau berpindah dari suatu patch ke patch yang lain. Koridor dapat menjadi
saluran untuk perpindahan, penghalang (barrier) atau penyaring (filter), misalnya untuk
aliran gen. Bentuk lain konektivitas habitat adalah batu loncatan (stepping stone) yaitu
satu atau lebih kantong habitat (habitat patches) yang secara ekologis terisolasi tapi
memberikan sumberdaya dan tempat pengungsian bagi satwa dalam perpindahan
menjelajahi suatu lansekap (Forman & Gordon, 1986).
2.4.5 Fragmentasi
Fragmentasi adalah proses pemecahan suatu habitat, ekosistem atau tipe land-
use menjadi bidang-bidang lahan yang lebih kecil. Fragmentasi juga merupakan suatu
hasil dimana proses fragmentasi mengubah atribut-atribut habitat dan karakteristik suatu
lansekap. Fragmentasi habitat mengubah konfigurasi spasial suatu kantong habitat
(habitat patch) besar dan menciptakan isolasi atau perenggangan hubungan antara
kantong-kantong habitat asli karena terselingi oleh mosaik yang luas atau tipe habitat
lain yang tidak sesuai bagi spesies yang ada (Wiens, 1995).
Franklin et al. (2002) mengembangkan definisi baru tantang fragmentasi sebagai
hasil (outcome) dan sebagai proses. Hasil (outcome) dari fragmentasi habitat adalah
diskontinuitas yang dihasilkan dari serangkaian mekanisme, di dalam distribusi spasial
suatu sumberdaya dan kondisi yang ada dalam suatu areal pada skala tertentu yang
mempengaruhi penghunian (occupancy), reproduksi atau survival suatu spesies. Proses
fragmentasi habitat didefinsikan sebagai serangkaian mekanisme yang mengakibatkan
diskontinuitas distribusi spasial suatu habitat. Ada empat komponen kunci dari dua
defininisi tersebut yaitu: (1) diskontinuitas, (2) mekanisme, (3) distribusi spasial dari
suatu sumberdaya dalam suatu areal, dan (4) atribut demografik (Franklin et al., 2002).
Konsep fragmentasi habitat diturunkan dari teori biogeografi pulau (MacArthur
& Wilson, 1967), di mana jumlah spesies meningkat dengan meningkatnya ukuran
pulau (Haila, 2002). Fragmentasi penting mendapat perhatian karena berpengaruh pada
kekayaan spesies dari komunitas, kecenderungan populasi beberapa spesies dan
keanekaragaman hayati ekosistem secara keseluruhan (Morrison et al., 1992).
Menurut Wilcove (1987) dalam Morrison et al. (1992) ada empat cara
fragmentasi dapat menyebabkan kepunahan lokal: (1) spesies mulai keluar dari kantong
habitat yang terlindungi; (2) kantong habitat gagal menyediakan habitat karena

36
pengurangan luas atau hilangnya heterogenitas internal; (3) fragmentasi menciptakan
populasi lebih kecil dan terisolasi yang memiliki resiko lebih besar terhadap bencana,
variabilitas demografik, kemunduran genetik atau disfungsi sosial; (4) fragmentasi dapat
mengganggu hubungan ekologis yang penting sehingga dapat menimbulkan sebab
sekunder kepunahan dari hilangnya spesies kunci dan pengaruh merugikan dari
lingkungan luar dan efek tepi (edge effect).
Fragmentasi umumnya terjadi melalui hilangnya habitat (habitat loss),
sebaliknya hilangnya habitat dapat dipandang sebagai akibat fragmentasi. Tetapi
fragmentasi dapat disertai hilangnya habitat (berkurangnya jumlah) seiring dengan
pemecahan atau pembagian kantong habitat besar menjadi kantong-kantong habitat
berukuran kecil dan lebih terisolasi (Hunter, 1997; Haila, 1999; Franklin et al., 2002;
Fahrig, 2003). Ketika hilangnya habitat dan fragmentasi dipandang secara terpisah,
hilangnya habitat memiliki konsekuensi lebih signifikan bagi kelangsungan hidup
(viability) spesies (Haila, 2002; Fahrig, 2003). Meskipun demikian, karena fragmentasi
dan hilangnya habitat terjadi bersamaan, maka sangat sulit untuk menentukan mana
yang lebih penting bagi perubahan habitat (Haila, 1999).
Fragmentasi bekerja dalam empat cara ketika hilangnya habitat dan fragmentasi
digabung untuk menggambarkan dan mengkategorikan prosesnya (Franklin et al., 2002;
Fahrig, 2003): (1) habitat hilang tanpa fragmentasi; (2) pengaruh kombinasi hilangnya
habitat dan pemecahan habitat menjadi patch-patch lebih kecil; (3) pemecahan habitat
menjadi patch-patch lebih kecil tanpa kehilangan habitat; dan (4) hilangnya habitat dan
pemecahan habitat menjadi patch-patch lebih kecil serta penurunan kualitas habitat.
Contoh ini berlaku untuk lansekap yang terdiri lebih dari satu habitat dan dikelilingi
oleh matrix dalam suatu kesatuan lansekap. Kasus pertama dan kedua berlaku ketika
lansekap keseluruhan berisi satu habitat dan tidak ada matrix di sekelilingnya. Dalam
kenyataan, kasus dua dan empat merupakan cara yang paling umum untuk habitat
terfragmentasi.
Fragmentasi habitat merupakan satu aspek dari tahapan proses yang secara
spasial dan temporal mengubah habitat dan lansekap yang diakibatkan oleh sebab-sebab
alami maupun antropogenik (Forman, 1995). Walaupun demikian, perubahan habitat
tidak dapat dihindari karena tidak ada habitat atau lansekap yang tetap (Forman, 1995).
Lanskap berubah melalui lima proses spasial dengan berbagai derajat overlap sepanjang

37
periode perubahan lahan (Forman, 1995), dan fragmentasi hanyalah satu outcome.
Proses ini dapat diakibatkan oleh penyebab alami dan antropogenik. Perforasi
(perforation) merupakan proses membuat lubang di dalam habitat. Pemotongan
(dissection) adalah pemotongan atau pembagian area menjadi habitat berbeda dengan
lebar yang relatif sama. Fragmentasi (fragmentation) adalah pemecahan habitat
menjadi potongan-potongan yang lebih kecil. Penyusutan (shrinkage) terjadi seiring
potongan habitat berlanjut dengan penurunan luas. Erosi habitat (attrition) adalah
proses dimana kantong habitat yang tersisa berangsur hilang karena degradasi habitat
atau suksesi
Fragmentasi dimulai dengan dissection ketika jalan, jaringan transmisi, sungai
dan fitur linear lainnya menjadi penghalang pergerakan. Kemudian diikuti perforation
ketika muncul kantong-kantong habitat (patches) kecil yang dibuat oleh manusia atau
sebab alami dan efek tepi menjadi nyata. Fragmentasi merupakan tahap ketiga yang
terjadi ketika kantong habitat yang lebih kecil meningkat frekuensinya dan berkurang
luasnya sampai pada tingkat di mana habitat yang terfragmentasi mulai mendominasi
lansekap. Attrition merupakan tahap akhir di mana lahan alami atau habitat asli tersisa
sebagai kantong yang kecil dan terisolasi di tengah-tengah lansekap yang sekarang
didominasi oleh suatu mosaik habitat yang telah berubah dan terfragmentasi. Proses
fragmentasi membuat habitat menjadi tidak sesuai atau memiliki kesesuaian rendah
bersamaan dengan berkurangnya kualitas habitat satwaliar. Sebaliknya, jika proses
gangguan mengubah mosaik habitat tetapi tidak ada perubahan kualitas habitat berarti
tidak terjadi fragmentasi, atau habitatnya berubah tetapi tidak terfragmentasi (Hunter,
1997)
Menurut Kupfer et al. (2004) ada empat cara primer fragmentasi hutan dapat
mempengaruhi keanekaragaman hayati, yaitu: (1) pengaruh perwakilan (sample effect);
(2) pengaruh luas (area effect); (3) pengaruh isolasi (isolation effect) dan (4) pengaruh
tepi (edge effect). Masing-masing pada gilirannya akan berpengaruh pada sebaran
populasi, komunitas dan proses ekosistem (Gambar 2.4).

.
Gambar 2
M
pengaruh
2003). A
dengan fr
(Franklin
Fr
positifnya
yang ber
Fragment
terbentuk
(3) habita
aktifitas n
merugika
2.4. Model
Mekanisme d
h ukuran patc
Ahli satwalia
fragmentasi h
n et al., 2002
ragmentasi h
a adalah m
rmanfaat, da
tasi member
k kantong ha
at-habitat tid
non kehutan
an spesies int
l konseptual
dan proses
ch; (2) peng
ar harus me
habitat dan m
2; Fahrig, 200
habitat dapa
meningkatkan
an meningk
rikan penga
abitat lebih k
dak lagi bersa
nan; dan (4)
terior (Barne
pengaruh fr
fragmentas
aruh tepi (ed
emperhatikan
masing-mas
03).
at dipandang
n keragaman
atkan edge
aruh negatif
kecil yang m
ambungan, k
) jumlah edg
es, 2000).
ragmentasi .
si menghasi
dge effect); d
n semua kar
ing memerlu
g dari segi
n habitat, m
yang disuk
ketika : (1
endorong pa
khususnya ji
ge meningka
.
ilkan tiga t
dan (3) peng
rena ketigan
ukan penang
positif dan
menciptakan
kai spesies
) ada habit
ada kepunah
ika fragment
at sehingga
tipe pengar
garuh isolasi
nya biasanya
ganan yang
negatif. P
penjajaran
satwaliar ge
at yang hila
an lokal dan
tasi disebabk
fragmentasi
38
ruh: (1)
(Fahrig
a terjadi
berbeda
engaruh
habitat
eneralis.
ang; (2)
n isolasi;
kan oleh
i habitat

39
2.4.6 Mengukur Fragmentasi
Indeks-indeks struktur lansekap seringkali digunakan untuk
mengkuantifikasikan fragmentasi habitat. Indeks-indeks telah dikembangkan untuk
mengukur tiga aspek struktur lansekap : (1) komposisi lansekap; (2) konfigurasi
lansekap; dan (3) bentuk-bentuk patch di dalam lansekap. Komposisi menunjukkan
jumlah dari tipe penutupan (cover) yang berbeda yang ditemukan dalam lansekap.
Konfigurasi menunjukkan bagaimana patch-patch dari tipe cover yang sama atau
berbeda tersusun di dalam lansekap dan hubungannya satu sama lain. Lanskap dengan
komposisi sama dapat memiliki konfigurasi yang berbeda, sehingga diperlukan
beberapa indeks aspek untuk menggambarkan suatu lansekap. Harus dicatat bahwa
beberapa indeks tidak sesuai benar untuk ketiga kategori tersebut83.
Program patch analyst yang kompatibel dengan ArcView 3.x cukup handal
untuk menghitung statistik fragmentasi, karena merupakan modifikasi dari program
Fragstats dan dapat digunakan untuk menghitung statistik spasial, baik file polygon
(seperti shape files) maupun file raster (seperti Arc grids) (Elkie et al., 1999). Empat
parameter fragmentasi yang akan diukur dalam penelitian ini adalah sebagai berikut:
(1) Class Area (CA)
Class area (luas kelas) bisa dihitung dari data vektor ataupun raster dan memiliki
satuan hektar. Nilai luas kelas berkisar > 0 sampai tak terhingga. Nilai Class area
akan mendekati 0 seiring tipe patch menjadi semakin jarang di dalam lansekap. Class
area = Total area (TA) ketika seluruh lansekap berisi satu tipe patch, yaitu ketika
seluruh citra berisi hanya satu patch. Class area sama dengan jumlah luas (m2) seluruh
patch dari semua tipe patch dibagi 10.000 (untuk konversi menjadi hektar) atau
diformulakan sebagai berikut:
⎟⎠⎞
⎜⎝⎛= ∑
= 000.101
1
n
jijaCA ............................................. Formula 2.3.
dimana aij luas patch ij (m2), dimana j = 1, ... , n patches, dan i = 1, ... , m or m’ tipe
patch (class).
83 http://chesapeake.towson.edu/landscape/forestfrag/

40
(2) Number of Patches (NumP)
Number of patches (jumlah patch) sama dengan jumlah patch dari semua tipe
patch (class). Jumlah patch berkisar dari satu sampai tak terbatas. Jumlah patch sama
dengan satu jika lansekap hanya berisi satu patch atau ketika kelas terdiri dari satu
patch. Jumlah patch tidak memiliki satuan dan formulanya adalah sebagai berikut:
niNP = ................................................... Formula 2.4.
dimana ni adalah jumlah patch dalam lansekap dari tipe patch (class) i.
(3) Total Edge (TE)
Total edge adalah jumlah panjang (m) dari semua segmen edge dari semua tipe
patch. Jika ada batas tepi (border) lansekap. TE mecakup segmen boundary lansekap
meliputi tipe patch dan hanya menggambarkan edge sebenarnya (disebut contrast
weight > 0). Jika suatu lansekap tidak memiliki border, TE mencakup proporsi segmen
boundary lansekap yang ditentukan oleh pemakai dari tipe patch yang dimaksud.
Tanpa memperhatikan apakah border lansekap ada atau tidak, TE mencakup proporsi
segmen edge background yang ditentukan oleh pemakai dari tipe patch dimaksud. TE =
0 ketika tidak ada edge kelas dari lansekap, yaitu ketika seluruh lansekap dan border
lansekap, jika ada, berisi tipe patch yang dimaksud dan ketentuan dari pemakai tidak
ada boundary lansekap dan background edge diperlakukan sebagai edge. TE bisa
dihitung dari data vaktor maupun raster dan satuannya adalah meter, dengan formula
sebagai berikut:
∑=
='
1
m
kjkeTE ............................................................ Formula 2.5.
dimana eik adalah panjang total (m) dari edge dalam lansekap antara tipe-tipe patch
(kelas-kelas) i dan k; mencakup segmen-segmen boundary lansekap yang
menggambarkan pohon edge (tree edge) hanya melibatkan tipe patch i; k = 1, ... , m
atau m’ tipe patch (kelas); m’ adalah jumlah tipe patch (kelas) yang ada di dalam
lansekap tidak termasuk border lansekap, jika ada.
(4) Edge Density (ED)
Edge density (kepadatan edge) sama dengan jumlah panjang (m) dari semua
segmen edge meliputi tipe patch yang dimaksud, dibagi dengan total luas lansekap (m2),

41
dikalikan dengan 10.000 (untuk konversi ke hektar). Jika ada border lansekap, ED
mencakup segmen boundary lansekap meliputi tipe patch dimaksud dan hanya
menggambarkan edge yang sebenarnya (disebut contrast weight > 0). Jika border
lansekap tidak ada, ED mencakup proporsi segmen boundary lansekap yang ditetapkan
oleh pemakai yang meliputi tipe patch dimaksud. Terlepas dari apakah suatu border
lansekap ada atau tidak, ED mencakup proporsi segmen edge background yang
ditetapkan oleh pemakai yang meliputi tipe patch dimaksud. ED bisa diperoleh dari
data vektor atau raster dengan satuan meter per hektar. Kisaran nilai ED ≥0 sampai tak
terhingga. Formula ED adalah sebagai berikut:
)100(
'
1
AED
m
kjke∑
== ............................................. Formula 2.6.
eik adalah panjang total (m) edge dalam lansekap antar tipe-tipe patch (kelas) i dan k;
meliputi segmen boundary lansekap hanya menggambarkan tree edge meliputi tipe
patch i; m’ adalah jumlah tipe patch (kelas) yang ada dalam lansekap, mencakup border
lansakap, jika ada; dan A adalah luas total lansekap.
2.5 Metapopulasi
Memahami dan memprediksi respon satwaliar terhadap konfigurasi patch dan
fragmentasi pada skala lansekap memerlukan pemahaman tentang dinamika
metapopulasi (Morrison et al. 1992). Levins (1970) merupakan orang pertama yang
menggunakan istilah metapopulasi dan memperkenalkan konsep metapopulasi sebagai
suatu populasi yang tersusun atas populasi-populasi lokal yang dianggap akan mati dan
dikolonisasi lagi secara lokal. Ia memperkenalkan model matematik untuk
menggambarkan metapopulasi:
dp/dt = m p (1 - p) - e p , ................................................. (Formula 2.7)
di mana p adalah proporsi (fraksi) pusat-pusat populasi (seperti pulau habitat atau
patch), m adalah laju migrasi (kolonisasi), dan e adalah laju dimana populasi lokal
menjadi punah.
Pada equilibrium p*=1 – e/m. Metapopulasi akan terjadi (yaitu p* > 0) hanya jika e <
m.

42
Suatu metapopulasi dapat hidup di suatu wilayah hanya jika laju rata-rata
kepunahan lebih kecil dari laju rata-rata migrasi. Populasi-populasi lokal terbangun
pada patch habitat tertentu yang bisa ditempati atau kosong pada suatu waktu.
Beberapa individu yang menyebar dapat meninggalkan suatu patch pergi untuk
mengkolonisasi suatu elemen kosong atau mengisi kembali (reinforce) suatu populasi
kecil. Populasi-populasi yang terbentuk di dalam sebuah patch dapat menghilang
megikuti kejadian lingkungan (kebakaran, pohon tumbang) atau kejadian demografik
(epidemik, penuaan). Model Levins sangat sederhana. Ia memberikan setiap patch
nilai yang sama sebagai sumber individu yang menyebar dan memiliki peluang
kepunahan yang sama, dan peluang keberhasilan dispersal adalah sama di semua patch
(Burel dan Baudry, 2003).
Model dan konsep Levins telah menjadi metapopulasi klasik. Definisi yang
lebih baru dibuat oleh Hanski dan Gilpin (1991), yaitu sekumpulan (set) populasi-
populasi lokal yang berinteraksi melalui perpindahan individu antar populasi-populasi
tersebut. Hanski dan Simberloff (1997) mendefinisikan metapopulasi sebagai
sekumpulan (set) populasi-populasi lokal di dalam beberapa area yang lebih besar, di
mana ditandai migrasi dari satu populasi lokal ke beberapa patch lain yang
memungkinkan.
Metapopulasi umumnya terjadi ketika kondisi lingkungan dan karakteristik
spesies memberikan pertukaran yang kurang sempurna dari sumber individu dan genetik
di antara sub populasi. Ini terjadi khususnya ketika habitat berada pada kondisi
heterogen pada suatu wilayah, provinsi atau lansekap yang menyebabkan isolasi parsial
individu yang berkembangbiak (Morrison et al., 1992).
Tidak semua kumpulan kantong-kantong populasi (patchy populations)
merupakan metapopulasi (Hanski & Simberloff, 1997), ada tiga tipe metapopulasi yang
berbeda menyimpang dari satu atau lebih asumsi metapopulasi model Levins (Harrison
& Taylor 1997) (Gambar 2.5). Asumsi metapopulasi model Levins adalah: (1) habitat
yang sesuai terjadi dalam patches diskrit di mana perkembangbiakan populasi lokal
mungkin ada; (2) semua populasi lokal memiliki resiko kepunahan yang besar. Jika
tidak, metapopulasi ”mainland-islands” dapat menjadi bukti karena keberadaan
metapopulasi akan tergantung hanya pada keberadaan populasi terbesar; (3) isolasi
patches tidak menghalangi rekolonisasi. Jika rekolonisasi tidak dapat terjadi,

43
metapopulasi akan berada pada keadaan non equilibrium dan dalam bahaya kepunahan;
(4) dinamika di antara populasi lokal tidak seluruhnya sempurna. Jika populasi lokal
mengalami proses-proses secara simultan, metapopulasi yang ada akan bergantung
hanya pada keberadaan populasi lokal dengan resiko kepunahan terkecil.
Gambar 2.5. Tipe-tipe metapopulasi.
Gambar 2.5 dapat dijelaskan sebagai berikut (Harrison & Taylor 1997):
Classic metapopulation: suatu jaringan besar dari patches kecil yang serupa, dengan
dinamika lokal terjadi pada skala waktu yang jauh lebih cepat dibandingkan
dinamika metapopulasi, dalam arti luas digunakan untuk sistem di mana semua
populasi lokal, meski mungkin mereka berbeda dalam ukuran, tapi memiliki satu
resiko kepunahan yang signifikan.
Mainland-island: sistem dari habitat patches (islands) berlokasi di dalam jarak
sebaran dari suatu habitat yang sangat besar (mainland) di mana populasi lokal tidak
akan pernah punah.
Nonequilibrium metapopulation: metapopulasi di mana laju kepunahan (jangka
panjang) melampaui laju kolonisasi atau sebaliknya; suatu kasus ekstrim di mana
“Classic” (Levins) metapopulation
Mainland – Island metapopulation
Nonequilibrium metapopulation Patchy Population
Keterangan : patch hitam berpenghuni, patch putih kosong Sumber: Hanski & Simberloff (1997); Harrison & Taylor (1997)

44
populasi-populasi lokal berlokasi sangat berjauhan satu sama lain sampai tidak ada
migrasi di antara mereka sehingga tidak ada peluang rekolonisasi. Sesungguhnya,
mereka mungkin tidak mengambarkan metapopulasi sama sekali, tetapi lebih
sebagai kumpulan populasi terisolasi yang tidak saling berinteraksi
Patchy population: suatu model metapopulasi di mana laju migrasi antar sub
populasi sangat tinggi sehingga dapat dikatakan secara efektif merupakan satu
populasi. Dalam patchy population, suatu individu mungkin merupakan bagian dari
lebih satu sub populasi sepanjang hidupnya.
Model metapopulasi memandang suatu populasi sebagai sejumlah sub populasi
deskrit di dalam kantong-kantong habitat (habitat patches) atau “pulau-pulau” yang
terhubungkan oleh dispersal. Seiring waktu, sub populasi bisa menjadi punah dan
dikolonisasi kembali, tetapi metapopulasi yang lebih besar dapat bertahan (Levins,
1969a; 1970). Paradigma dinamika metapopulasi juga serupa dengan model
biogeografi pulau dari MacArthur & Wilson (1967) dalam memandang kepunahan dan
kolonisasi (Hanski & Gilpin, 1991). Bedanya dalam biogeografi pulau ada daratan
utama sebagai sumber kolonisasi sedangkan dalam metapopulasi, kolonisasi terjadi di
antara sub populasi (Singleton et al., 2002).
Suatu metapopulasi merupakan kumpulan dari populasi-populasi kecil yang
menempati sejumlah kantong habitat (Gambar 2.6)84. Individu-individu secara tidak
teratur berpindah antar patch, dan populasi dapat menjadi punah di dalam patch
individual sebagai akibat dari peluang kejadian (chance events). Sebagai contoh, dua
jantan dalam suatu populasi dari enam betina dapat dimakan predator. Jika tidak ada
jantan lain yang imigrasi ke dalam patch tersebut dari patch lain, populasi di dalam
patch tersebut akan mengalami kepunahan. Populasi-populasi kecil dianggap rawan
mengalami tipe peluang kepunahan ini. Tetapi, karena individu-individu secara tidak
teratur berpindah antar patch, maka patch yang kosong akhirnya akan dikolonisasi dan
ditempati lagi di masa mendatang (Gambar 2.7)85.
84 http://chesapeake.towson.edu/landscape/forestfrag/effects.asp 85 http://chesapeake.towson.edu/landscape/forestfrag/effects.asp

45
Sumber : http://chesapeake.towson.edu/landscape/forestfrag/effects.asp
Gambar 2.6. Sekumpulan patch hutan yang ditempati oleh populasi-populasi penghuni
hutan. Tebal tipisnya panah menunjukkan laju pertukaran individu antar populasi (panah tebal menunjukkan lebih banyak pertukaran).
Sumber : http://chesapeake.towson.edu/landscape/forestfrag/effects.asp
Gambar 2.7. Penghunian sekumpulan patch hutan yang mendukung suatu metapopulasi
spesies penghuni hutan pada dua waktu yang berbeda. Patch hitam ditempati dan patch putih tidak ditempati.
Jika laju kolonisasi pada patch-patch kosong lebih tinggi dari laju kepunahan,
metapopulasi akan bertahan. Hal ini karena ketika beberapa patch mengalami
kepunahan, yang lainnya dikolonisasi. Akibatnya, perpindahan individu-individu antar
populasi mengikat semua populasi ke dalam suatu metapopulasi yang dapat bertahan
dalam lansekap yang terganggu86.
86 http://chesapeake.towson.edu/landscape/forestfrag/effects.asp

46
2.6. Seleksi Habitat
Satwaliar mungkin menilai dan memilih habitatnya yang cocok (sesuai)
sehingga dapat bertahan hidup dan berkembangbiak yang terbaik dengan melihat faktor-
faktor nutrisi dan struktural. Meskipun demikian kebanyakan studi tentang mekanisme
seleksi habitat menitik beratkan pada respon satwa terhadap faktor-faktor struktural
(Bailey, 1984).
Seleksi habitat merupakan spesialisasi, bagi suatu spesies memilih tipe habitat
tertentu berarti membatasi diri pada habitat tersebut dan akan mencapai adaptasi
terutama kecocokan dalam penggunaan sumberdaya makanan dan cover dari habitat
terpilih. Oleh karena itu seleksi habitat menyerupai isolasi relung dan diperkuat oleh
kompetisi interspesifik (Svardson, 1949 dalam Bailey, 1984). Seleksi habitat
bermanfaat untuk menunjukkan bagaimana gigihnya suatu jenis satwa setia pada
pilihannya terhadap struktur habitat tertentu. Meskipun belajar, kesan terhadap tempat
lahirnya mungkin mempengaruhi preferensi habitat, preferensi ini sangat mungkin
adalah bawaan sejak lahir (Bailey, 1984; Robinson dan Bolen, 1984).
Dalam seleksi habitat, beberapa vertebrata lebih mementingkan life form atau
fisiognomi habitat mereka, daripada keberadaan spesies tumbuhan tertentu. Satwaliar
mungkin hanya tergantung pada kebutuhan cover (aspek struktural dari lingkungannya)
seperti halnya ketergantungannya pada kebutuhuan mereka akan makanan tertentu.
Beberapa spesies adalah generalis, tak tarbatasi oleh ketersediaan bentuk-bentuk cover,
sedangkan jenis lainnya spesialis, memiliki kebutuhan cover tertentu saja (Bailey, 1984)
Evolusi preferensi habitat ditentukan oleh struktur morfologi dan fungsi-fungsi
tingkah laku, kemampuannya memperoleh makanan dan lindungan (shelter) dengan
sukses di habitatnya. Faktor-faktor yang mendorong untuk memilih suatu habitat antara
lain adalah ciri struktural dari lansekap, peluang mencari makan dan bersarang atau
keberadaan spesies lain (Cody, 1985). Studi seleksi habitat di mana menekankan pada
ekologi, tingkah laku dan fisiologi akan bermanfaat jika dapat menunjukkan, misalnya,
bahwa ciri-ciri habitat tertentu menentukan perbedaan fisiologi antara spesies yang
sekerabat atau bahwa suatu spesies mengembangkan tingkah laku tertentu dalam
mencari makan sehubungan dengan ciri-ciri struktural tertentu dari habitatnya (Cody,
1985).

47
Memahami seleksi habitat merupakan dasar yang penting untuk mengerti sejarah
alami satwaliar (Manly et al., 1993). Dengan demikian tidak mengherankan banyak
penelitian dilakukan bertujuan untuk mengembangkan perhitungan analisis habitat (Neu
et al., 1974; Johnson, 1980; Marcum & Lohftsgaarden, 1980; Aebischer et al., 1993;
McCracken et al., 1998). Seleksi habitat ditentukan berdasarkan penggunaan habitat
yang tercatat dan penggunaan habitat yang diharapkan sebagaimana ditetapkan sebagai
sebuah model null (Johnson, 1980). Jika habitat yang digunakan relatif lebih besar dari
yang diharapkan maka habitat tersebut dianggap “sangat disukai” dan jika habitat yang
digunakan relatif lebih sedikit dari yang diharapkan maka habitat tersebut dianggap
sebagai “kurang disukai”.
Lebih lanjut, habitat yang digunakan lebih dari yang diharapkan disebut sebagai
“habitat yang disukai” (Preferred habitat) dan habitat yang kurang digunakan dari yang
diharapkan disebut sebagai habitat yang dihindari (avoided habitat) (Neu et al., 1974).
Banyak perhitungan seleksi habitat mengandalkan klasifikasi lokasi satwaliar
berdasarkan tipe habitatnya dan menghitung proporsi penggunaannya (Neu et al., 1974;
Johnson, 1980; Aebischer et al., 1993). Teknik seperti ini disebut sebagai “pendekatan
klasifikasi” (classification approaches). Sebaliknya, metode yang didasarkan pada
penghitungan jarak Euclidean dari lokasi satwaliar ke habitat fiturnya disebut sebagai
“pendekatan jarak” (distance based approaches) (Krebs, 1999).
Metode Neu telah digunakan untuk menghitung indeks seleksi habitat yang
kemudian distandarisasi untuk studi perbandingan. Metode ini menggunakan proporsi
lokasi ditemukannya satwaliar di dalam tipe-tipe habitat yang diteliti dibandingkan dengan
proporsi ketersediaan (availability) tipe-tipe habitat tersebut (Manly et al., 1993).
Seleksi habitat diukur menggunakan fungsi-fungsi seleksi sumberdaya (Manly et
al., 2002) untuk memperkirakan peluang suatu unit contoh digunakan oleh seekor
satwa, sebagai fungsi dari variabel vegetasi habitat (Sawyer et al., 2009). Pengujian
signifikansi menggunakan uji Chi-square (χ2) (Fleis, 1981). Chundawat (1990)
menentukan preferensi habitat macan tutul salju (Panthera uncia) dengan uji Chi
Square seperti yang disarankan Neu et al. (1974).

48
2.7. Kesesuaian Habitat (Habitat Suitability)
Habitat satwaliar terus-menerus mengalami perubahan (dinamis), baik oleh
sebab alami maupun akibat kegiatan manusia. Perubahan habitat banyak disebabkan
oleh kegiatan manusia yang mengubah tutupan lahan (land cover) atau penggunaan
lahan (land use) seperti pengembangan pertanian, pemukiman, industri, jaringan
tranportasi, jaringan listrik dan lain-lain (Bureau of Land Management, 2004).
Pengetahuan hubungan antara penyebaran satwa dengan habitatnya memegang
peranan sangat penting dalam merancang pengelolaan spesies terancam punah (Lecis &
Noris, 2004). Sementara pengetahuan tentang preferensi habitat juga penting untuk
mengetahui distribusi spasial satwa dalam habitat-habitat yang sesuai (Osborne et al.,
2001). Habitat yang sesuai (suitable) adalah habitat yang dengan kondisinya mampu
menyediakan kebutuhan hidup suatu spesies (Juntti & Rumble, 2006). Kesesuaian
habitat saat ini sudah banyak dibuat dalam suatu model menggunakan piranti lunak
berbasis komputer. Model kesesuaian habitat (habitat suitability) merupakan metode
yang efisien dan murah serta cepat untuk menilai kualitas habitat satwaliar (Brooks
1997; Schamberger et al., 1982; Cole & Smith, 1983).
Geographic Information System (GIS), merupakan alat yang handal untuk
menilai dan mengevaluasi pola-pola lansekap dan perubahannya pada wilayah yang luas
(Sessions et al., 1994). GIS banyak diaplikasikan untuk menilai fragmentasi habitat dan
membuat pemodelan distribusi spesies (Apan, 1996; Jorge & Garcia 1997; He et al.,
1998). Model-model kesesuaian habitat telah banyak digunakan untuk mengevaluasi
habitat satwaliar serta dampak kegiatan manajemen dan pembangunan sejak awal tahun
1980-an. Model-model tersebut didasarkan pada hubungan fungsional antara satwaliar
dan variabel-variabel habitat (U.S. Fish and Wildlife Service, 1980; 1981).
Pemodelan spasial adalah prosedur analitik yang diaplikasikan dengan Sistim
Informasi Geografis (SIG), di mana serangkaian prosedur mensimulasikan kondisi
dunia nyata ke dalam SIG dengan menggunakan hubungan spasial dari fitur-fitur
geografis (AGI, 2010). Pembuatan pemodelan spasial bertujuan membantu pengambil
keputusan ataupun analis untuk memahami, menggambarkan dan memperkirakan
bagaimana suatu proses bekerja dalam dunia nyata melalui penyederhanaan fenomena
maupun fitur (Jaya, 2007). Pemodelan spasial kesesuaian habitat secara kuantitaif
sangat penting untuk pengelolaan populasi satwaliar dan perencanaan strategi

49
konservasi pada skala lansekap (U.S. Fish and Wildlife Service, 1980; 1981; Osborne et
al., 2001) serta prioritasisasi upaya-upaya konservasi dengan lebih efisien dan efektif
(Kushwaha et al., 2004).
Kesesuaian habitat dinilai untuk setiap fitur spasial dalam wilayah yang diteliti
untuk setiap periode waktu. Salah satu pendekatan umum untuk evaluasi kuantitatif
habitat adalah dengan membuat model indeks kesesuaian habitat (Habitat Suitability
Index) untuk membangun nilai dari suatu habitat tertentu berdasarkan pada preferensi
satwa terhadap penggunaan komponen-komponen habitat (U.S. Fish and Wildlife
Service, 1981; Rickers et al,. 1995). McLellan et al. (1995) mengembangkan suatu
model HSI untuk caribou gunung berdasarkan elevasi, slope, land cover atau tipe
habitat dan umur tegakan dengan notasi sebagai berikut:
HSItotal = (HSIelevation * HSIslope * HSIhabitat unit * HSIstand age)1/4 ................. (Formula 2.8)
Kesesuaian (suitability) didefinisikan sebagai kemampuan habitat dalam
kondisinya saat ini untuk menyediakan kebutuhan hidup suatu spesies. Ini merupakan
suatu perkiraan bagaimana suatu kondisi habitat sekarang memberikan kebutuhan hidup
tertentu suatu spesies. Rangking kesesuaian diberikan kepada setiap struktur dari setiap
tipe vegetasi. Struktur-struktur dengan rangking kesesuaian tertingi menggambarkan
kemampuannya untuk beberapa ekosistem tertentu. Untuk membuat kesesuaian, maka
diberikan rangking (rating) pada struktur habitat untuk potensinya mendukung spesies
tertentu pada musim tertentu. Hal tersebut menggambarkan suatu proporsi dari habitat
terbaik yang mencerminkan harapan penggunaan habitat oleh spesies satwaliar (Juntti
and Rumble, 2006). Secara numerik, indeks kesesuaian habitat (HSI) dibuat oleh U.S.
Fish and Wildlife Service (1980) sebagai berikut :
........................... (Formula 2.9)
Jika kondisi-kondisi aktual identik dengan kondisi-kondisi optimum, maka HSI
sama dengan satu. Nilai indeks kesesuaian habitat biasanya dihitung menggunakan
formula matematis yang menggambarkan hubungan hipotetik di antara SI individual.
Hubungan satwaliar-habitat dapat didukung data empiris, pendapat ahli atau keduanya
(U.S. Fish and Wildlife Service 1980; 1981). Secara tradisional model HSI diterapkan
pada suatu contoh lokasi dalam tipe-tipe penutupan lahan atau tipe-tipe vegetasi
HSI = Study Area Habitat Conditions Optimum Habitat Conditions

50
dominan. Kualitas habitat pada suatu area merupakan penjumlahan unit-unit habitat
yang menggambarkan hasil dari rata-rata nilai HSI di setiap tipe vegetasi dan area dari
suatu lahan pada tipe vegetasi tersebut, dijumlahkan seluruh area studi (Larson et al.,
2003).