2011imt_tanggap fungsional parasitoid (bab 5)
TRANSCRIPT

46
BAB V
TANGGAP FUNGSIONAL PARASITOID Anastatus dasyni FERR. (HYMENOPTERA: EUPELMIDAE)
TERHADAP TELUR KEPIK PENGISAP BUAH LADA
[Functional response of Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae) to eggs of pepper bug]
Abstrak
Anastatus dasyni adalah salah satu spesies parasitoid yang paling penting yang memarasit telur kepik pengisap buah lada Dasynus piperis. Studi dilakukan dengan tujuan untuk mengukur tanggap fungsional parasitoid di laboratorium. Parasitoid dipelihara secara individu di dalam tabung gelas (d = 1,5 cm; p = 17.0 cm) yang di dalamnya berisi telur kepik lada dengan kerapatan 1, 2, 3, 4, 6, 8, 10, 12, atau 14 telur D. piperis. Regresi logistik diterapkan untuk memeriksa bentuk tanggap fungsional. Regresi nonlinear digunakan untuk menduga laju pencarian seketika (a) dan masa penanganan inang (Th). Hasil penelitian mengungkapkan bahwa parasitoid A. dasyni memperlihatkan tanggap fungsional tipe II. Persamaan cakram Holling dan persamaan acak sama sesuainya terhadap data pengamatan dengan R2 = 0.91. Laju pencarian inang seketika dan masa penanganan inang berturut-turut adalah 0.049 ± 0.003 jam-1 dan 1.087 ± 0.135 jam untuk persamaan cakram, dan 0.194 ± 0.042 jam-1
Anastatus dasyni is one of the most important egg parasitoids of pepper bug, Dasynus piperis. This study was carried out to measure the functional response of the parasitoid in laboratory. The parasitoid was individually reared for 24 h in test tube (d=1.5 cm; l = 17.0 cm) containing either 1, 2, 3, 4, 6, 8, 10, 12, or 14 D. piperis eggs. A logistic regression analysis was performed to determine the type of functional response for the parasitoid. A nonlinear regression approach was used to estimate the instantaneous search rate (a) and handling time (Th). Results of the study showed that A. dasyni demonstrated a type II functional response. Holling’s disc equation and random equation fitted equally to the data with R
dan 2.011 ± 0.182 jam untuk persamaan acak. Jumlah maksimum telur yang dapat diparasit berdasarkan persamaan cakram dan persamaan acak masing-masing adalah 22 dan 12 butir. Mempertimbangan perilaku peneluran A. dasyni, persamaaan cakram tampaknya lebih sesuai untuk memerikan tanggap fungsional parasitoid.
Kata kunci: Anastatus dasyni, Dasynus piperis, parasitoid, tanggap fungsional
Abstract
2 value was 0.91. The instantaneous searching rate and handling time for Holling disc equation were 0.049 ± 0.003 h-1 and 1.087 ± 0.135 h, respectively, and for random equation 0.194 ± 0.042 h-1 and 2.011 ± 0.182 h, respectively. The maximum number of D. piperis eggs which were able to be parasitized based on the disc and random equations were 22 and 12 individuals, respectively. According to the oviposition behavior of A. dasyni, the disc

47
equation seemed to be more appropriate for describing the parasitoid functional response.
Key words: Anastatus dasyni, Dasynus piperis, parasitoid, functional response
Pendahuluan
Kepik pengisap buah lada, Dasynus piperis China (Hemiptera: Coreidae),
merupakan hama utama pada pertanaman lada. Laba et al. (2004) melaporkan
rataan tingkat serangan hama ini pada pertanaman lada di Bangka mencapai
36,8%. Di Lampung dilaporkan bahwa serangan kepik D. piperis mengakibatkan
penurunan hasil panen sebesar 15% (Suprapto & Thomas 1989). Untuk
mengendalikan hama ini, petani umumnya melakukan aplikasi insektisida setiap
bulan selama masa perbungaan hingga panen, atau sebanyak 10 kali aplikasi
selama setahun. Karena persyaratan yang ketat terhadap residu pestisida di
pasaran internasional, pengendalian hayati kini tengah dikembangkan sebagai
salah satu komponen utama pengendalian hama terpadu di perkebunan lada.
Salah satu musuh alami hama ini yang paling penting adalah parasitoid telur
Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae), dengan tingkat parasitisasi di
lapangan berkisar antara 75% hingga 84% (Deciyanto et al. 1993; Trisawa et al.
2007).
Kemampuan musuh alami mengatur populasi hama salah satunya
ditentukan oleh tanggap fungsional (Solomon 1949). Tanggap fungsional suatu
parasitoid didefinisikan sebagai hubungan antara banyaknya inang yang diparasit
dengan kerapatan inang. Dikenal ada tiga tipe tanggap fungsional yaitu I, II, dan
III (Holling 1959). Pada tanggap fungsional tipe I proporsi inang terparasit
bersifat konstan, sehingga hubungan antara banyaknya inang yang terparasit dan
kerapatan inang bersifat linear. Pada tanggap fungsional tipe II proporsi inang
yang terparasit menurun tajam dengan bertambahnya kerapatan inang. Pada
tanggap fungsional tipe III proporsi inang yang terparasit awalnya meningkat,
tetapi kemudian secara berangsur menurun dengan meningkatnya kerapatan inang.
Pengetahuan tentang tanggap fungsional dapat digunakan untuk menapis musuh
alami yang potensial dan memperkirakan potensi pengendalian hayati (Parella &
Horsburgh 1983; Houck & Strauss 1985).

48
Parameter penting dari tanggap fungsional adalah laju pencarian seketika
(a) dan masa penanganan inang (Th). Parasitoid yang baik adalah yang memiliki
nilai a yang tinggi dan nilai Th yang rendah (Hassell 2000). Informasi tentang
kedua parameter ini dapat memberikan pemahaman yang mendasar tentang
interaksi parasitoid-inang, sehingga diharapkan dapat memberikan landasan bagi
penyusunan strategi pengendalian hayati kepik lada dengan parasitoid A. dasyni.
Sehubungan dengan itu, penelitian ini bertujuan mengukur dan memerikan
tanggap fungsional A. dasyni terhadap peningkatan kerapatan inang telur kepik
buah lada.
Bahan dan Metode
Penelitian dilakukan sejak bulan April 2009 sampai dengan Juli 2009 di
laboratorium hama Balai Penelitian Tanaman Obat dan Aromatik Bogor, serta
laboratorium dan rumah kaca Balai Pengkajian Teknologi Pertanian Kepulauan
Bangka Belitung.
Pembiakan R. linearis
Serangga R. linearis yang diperoleh dari pertanaman kedelai dipelihara
dalam kurungan kain kasa (p = 35 cm, l = 35 cm, t = 75 cm) di laboratorium.
Serangga diberi pakan kacang panjang yang digantungkan pada kawat di bagian
atas kurungan. Pakan diganti setiap 2 hari sekali. Untuk tempat peneluran
serangga R. linearis, di dalam kurungan digantungkan untaian kain wol. Telur
yang diperoleh kemudian digunakan untuk pembiakan dan penelitian.
Pembiakan Parasitoid A. dasyni
Telur D. piperis dikumpulkan dari pertanaman lada di Bangka. Telur yang
terdapat pada daun, buah dan bagian tanaman yang lain diambil dan dipelihara
dalam tabung reaksi bergaris tengah 1.5 cm dan panjang 17.0 cm di laboratorium.
Tabung reaksi ditutup dengan kapas yang dibungkus kain kasa. Perkembangan
telur diamati sampai keluar imago A. dasyni. Imago A. dasyni dipindahkan ke
tabung reaksi lain yang berukuran sama dan diberi pakan madu 10%.
Telur R. linearis umur 3 hari hasil pembiakan di laboratorium diambil dan
digunakan untuk pembiakan A. dasyni. Pembiakan dilakukan dengan cara setiap
10 telur R. linearis dilem dengan lem kertas cair pada kertas karton (pias)

49
berukuran 1.0 cm x 5.0 cm. Pias telur kemudian dimasukkan ke dalam tabung
reaksi bergaris tengah 1.5 cm dan panjang 17.0 cm yang berisi sepasang parasitoid
A. dasyni umur 3 hari. Pias diambil setelah 24 jam dan diganti dengan pias yang
baru. Pias yang diambil dimasukkan ke dalam tabung reaksi bergaris tengah 1.5
cm dan panjang 17.0 cm. Pias diamati sampai parasitoid keluar. Parasitoid hasil
pembiakan digunakan untuk penelitian.
Pembiakan Telur D. piperis
Telur diperoleh dari pemeliharaan imago D. piperis pada bibit lada di
rumah kaca dan pertanaman lada di lapangan. Di rumah kaca, imago D. piperis
dipelihara dalam kurungan plastik milar bergaris tengah 18.0 cm dan tinggi 40.0
cm di rumah kaca. Kurungan tersebut menyungkup bibit lada dalam pot plastik
bergaris tengah 22.0 cm dan tinggi 14.5 cm. Imago D. piperis diberi pakan buah
lada umur 6 sampai 9 bulan yang digantungkan pada kawat di bagian atas
kurungan atau dilekatkan pada bibit lada. Pakan tersebut diganti setiap 2 hari.
Pemasukan serangga dan atau penggantian buah lada dilakukan melalui lubang
yang terdapat di bagian atas kurungan. Pada pemeliharaan di lapangan, serangga
dimasukkan ke dalam kurungan yang menyungkup cabang dan buah lada.
Tanggap Fungsional A. dasyni pada Kerapatan Inang
Telur D. piperis dilem dengan lem kertas cair pada pias karton berukuran
1.0 x 5.0 cm. Kerapatan telur yang diuji adalah 1, 2, 3, 4, 6, 8, 10, 12, dan 14
butir. Pias telur tersebut kemudian dimasukkan ke dalam tabung reaksi bergaris
tengah 1.5 cm dan panjang 17.0 cm. Dinding tabung reaksi dioles cairan madu
10% untuk pakan parasitoid. Setiap satu pasang parasitoid A. dasyni umur 3 hari
asal telur R. linearis dimasukkan ke dalam tabung reaksi sesuai perlakuan
kelimpahan inang yang diuji. Setelah 24 jam, telur D. piperis dikeluarkan dari
tabung reaksi dan dimasukkan ke dalam tabung reaksi lain yang berukuran sama,
kemudian diamati jumlah telur D. piperis yang diparasit. Percobaan diulang 10
kali dan disusun dalam rancangan acak lengkap.
Analisis Data
Pengaruh banyaknya inang tersedia terhadap rataan banyaknya inang
diparasit diperiksa dengan sidik ragam yang dilanjutkan dengan uji beda nyata

50
terkecil pada α = 0.05. Untuk menentukan tipe tanggap fungsional dilakukan
analisis regresi logistik antara kerapatan inang (N0) dengan proporsi inang yang
diparasit (Ne/N0) dengan persamaan sebagai berikut (Juliano 1993):
Ne exp (P0 + P1N0 + P2N02 + P3N0
3
= N
)
0 1 + exp (P0 + P1N0 + P2N02 + P3N0
3) dengan P0 adalah intersep, P1 koefisien linear, P2 koefisien kuadratik, dan P3
koefisien kubik. Analisis menggunakan prosedur CATMOD dari program SAS
(SAS Institute 1990). Jika koefisien linear P1 tidak berbeda nyata dengan 0,
model tanggap fungsional yang sesuai adalah tipe I. Bila koefisien linear P1
secara nyata < 0 model yang sesuai adalah tipe II, sedangkan bila P1 secara
nyata > 0 model yang sesuai adalah tipe III (Juliano 1993). Bila persamaan kubik
menghasilkan nilai P3 yang tidak nyata, maka disarankan parameter kubik
dikeluarkan dari persamaan dan kemudian dilakukan pengujian ulang terhadap
parameter yang tersisa. Hal yang sama berlaku pula bila persamaan kuadratik
menghasilkan nilai P2 yang tidak nyata.
Karena hasil analisis regresi logistik mengindikasikan tanggap fungsional
tipe II, maka analisis selanjutnya ditekankan pada pemeriksaan kesesuaian data
terhadap model tanggap fungsional tipe II. Untuk keperluan tersebut digunakan
model persamaan cakram dari Holling (1959) dan persamaan acak dari Rogers
(1972), sebagai berikut:
Persamaan cakram : Ne = aTNo/(1 + aThNo)
Persamaan acak : Ne = No {1-exp[a(ThNe – T)]}
dengan Ne adalah banyaknya inang yang diparasit, No banyaknya inang yang
disediakan, a laju pencarian seketika, T lama waktu inang terpapar pada
parasitoid, dan Th lama waktu penanganan inang.
Nilai penduga parameter (a dan Th) dari kedua model tersebut di atas
diperoleh melalui regresi non linear menggunakan prosedur PROC NLIN dari
SAS (SAS Institute 1990). Selanjutnya koefisien determinasi (R2 = 1 – (jumlah
kuadrat sisaan/jumlah kuadrat total terkoreksi) digunakan untuk memeriksa
kesesuaian model.

51
Hasil dan Pembahasan
Hasil analisis ragam menunjukkan bahwa kerapatan inang berpengaruh
sangat nyata (F = 110.09; db = 8, 81; P = < 0.0001) terhadap rataan banyaknya
inang yang diparasit (Tabel 5.1). Semakin meningkat kerapatan inang semakin
banyak inang yang diparasit, namun demikian proporsi inang yang diparasit
menurun. Pada kerapatan inang rendah (1 dan 2 butir) seluruh inang diparasit
(100%), kemudian proporsi inang yang diparasit menurun pada kerapatan inang
berikutnya. Pada kerapatan inang 12 dan 14 butir, rataan banyaknya telur yang
diparasit sekitar 9 butir atau proporsi yang terparasit 64-75%.
Tabel 5.1 Rataan inang yang diparasit (x ± SD) A. dasyni pada berbagai kerapatan inang
Kerapatan inang (butir)
Rataan inang yang diparasit *)
1
1.00 ± 0.00 a
2 2.00 ± 0.00 b
3 2.80 ± 0.42 c
4 3.70 ± 0.67 d
6 5.50 ± 0.71 e
8 7.00 ± 0.82 f
10 7.70 ± 1.34 f
12 9.00 ± 1.33 g
14 9.00 ± 1.49 g *)
Analisis regresi logistik dengan menggunakan persamaan kubik
menghasilkan nilai parameter kubik yang tidak nyata, begitu pula dengan model
kuadratik. Oleh karena itu, parameter P
Angka dalam kolom yang diikuti oleh huruf yang sama tidak berbeda nyata (uji beda nyata terkecil, α = 0.05) setelah ditransformasi ke √x + 0.5
3 dan P2 dikeluarkan dari persamaan dan
kemudian dilakukan pengujian ulang terhadap parameter sisanya. Hasil analisis
dengan menggunakan model tereduksi disajikan pada Tabel 5.2. Koefisien linear
(P1) yang bernilai negatif dan secara nyata < 0 menunjukkan bahwa proporsi
inang terparasit menurun dengan meningkatnya kerapatan inang seperti
ditunjukkan pada Gambar 5.1. Hal ini menunjukkan bahwa tanggap fungsional
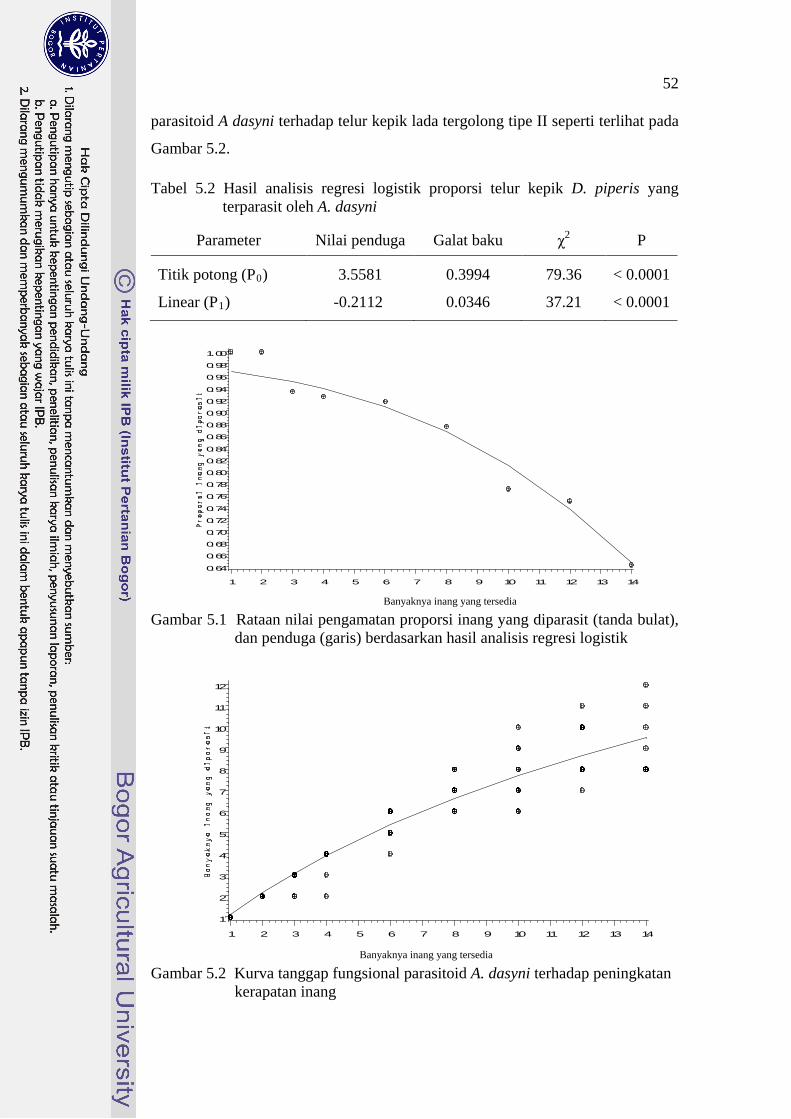
52
parasitoid A dasyni terhadap telur kepik lada tergolong tipe II seperti terlihat pada
Gambar 5.2. Tabel 5.2 Hasil analisis regresi logistik proporsi telur kepik D. piperis yang
terparasit oleh A. dasyni
Parameter
Nilai penduga
Galat baku
χ
P 2
Titik potong (P0
3.5581 )
0.3994
79.36
< 0.0001
Linear (P1 -0.2112 ) 0.0346 37.21 < 0.0001
0.64
0.66
0.68
0.70
0.72
0.74
0.76
0.78
0.80
0.82
0.84
0.86
0.88
0.90
0.92
0.94
0.96
0.98
1.00
Banyaknya inang yang tersedia
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Gambar 5.1 Rataan nilai pengamatan proporsi inang yang diparasit (tanda bulat),
dan penduga (garis) berdasarkan hasil analisis regresi logistik
1
2
3
4
5
6
7
8
9
10
11
12
Banyaknya inang yang tersedia
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Gambar 5.2 Kurva tanggap fungsional parasitoid A. dasyni terhadap peningkatan
kerapatan inang
Banyaknya inang yang tersedia
Banyaknya inang yang tersedia

53
Tanggap fungsional tipe II umum terdapat pada percobaan di laboratorium
atau kurungan di lapangan, dengan satu spesies inang atau mangsa yang
disediakan (Schenk & Baker 2002; Pervez & Omkar 2005). Hal ini terkait dengan
kemudahan parasitoid menemukan inangnya, karena parasitoid dan inangnya
berada dalam ruangan yang terbatas. Jika parasitoid berada pada area yang lebih
luas, maka parasitoid dapat memperlihatkan kurva tanggap fungsional tipe III
(sigmoid). Contoh kasus ini terdapat pada parasitoid Aphelinus thomsoni Graham
(Hymenoptera: Aphelinidae) yang memperlihatkan tanggap fungsional tipe II jika
dikurung bersama inangnya, dan memperlihatkan tanggap fungsional tipe III jika
parasitoid diberi kesempatan berpindah area percobaan (Collins et al. 1981).
Seperti disebutkan terdahulu, dalam penelitian ini tanggap fungsional tipe
II didekati dengan persamaan cakram dan persamaan acak. Berdasarkan kriteria
R2
Model tipe II
pada Tabel 5.3 tampak bahwa persamaan cakram maupun persamaan acak
memperlihatkan kesesuaian yang sama untuk digunakan memeriksa hubungan
antara kerapatan telur kepik lada dengan rataan banyaknya telur yang terparasit
oleh A. dasyni. Walaupun demikian, kedua persamaan ini menghasilkan penduga
a dan Th yang berbeda. Persamaan acak menghasilkan nilai penduga parameter
tanggap fungsional yang lebih tinggi daripada persamaan cakram.
Tabel 5.3 Nilai penduga parameter laju pencarian seketika (a) dan masa penanganan inang (Th) berdasarkan model tanggap fungsional tipe II
a (jam-1 Th (jam) ) R2
Persamaan cakram 0.049 ± 0.003 1.087 ± 0.135 0.91
Persamaan acak 0.194 ± 0.042 2.011 ± 0.182 0.91
Laju pencarian seketika (a) untuk parasitoid A. dasyni yang memarasit
telur kepik lada D. piperis menurut persamaan cakram adalah 0.049 ± 0.003 jam-1,
sedangkan menurut persamaan acak 0.194 ± 0.042 jam-1. Laju pencarian seketika
(a) menunjukkan proporsi dari total area yang dijelajahi parasitoid per unit waktu
jelajah, dalam hal ini adalah jam. Laju pencarian seketika ini menentukan
seberapa cepat kurva tanggap fungsional mencapai plato atas, dan merupakan
fungsi dari (1) jarak terjauh parasitoid mampu mendeteksi inang, (2) kecepatan

54
pergerakan dari parasitoid dan inang, dan (3) proporsi serangan yang berhasil
(Holling 1965).
Masa penanganan inang (Th) adalah lamanya waktu parasitoid mengenali,
mengejar, dan memarasit inang serta kegiatan lain yang terkait dengan parasitisasi
seperti membersihkan alat mulut dan beristirahat sebelum bergerak mencari inang
yang lain (Holling 1965). Nilai penduga Th pada persamaan cakram adalah 1.087
± 0.135 jam dan pada persamaan acak 2.011 ± 0.182 jam. Berdasarkan nilai Th
ini, dalam sehari parasitoid A. dasyni maksimum dapat memarasit sekitar 22 butir
telur menurut persamaan cakram dan 12 butir menurut persamaan acak.
Kemampuan parasitisasi ini perlu diketahui dalam rangka mengevaluasi potensi
suatu spesies parasitoid sebagai agens pengendalian hayati. Namun demikian,
hasil yang diperoleh di laboratorium tidak secara langsung dapat diterapkan pada
kondisi lapangan. Di lapangan banyak faktor yang mempengaruhi tanggap
fungsional (Parella & Horsburgh 1983; Messina & Hanks 1998) seperti fase
pertumbuhan tanaman, cuaca, inang alternatif, dan kompetisi dengan serangga
lain.
Menurut Rogers (1972), persamaan cakram dan persamaan acak dapat
digunakan untuk memerikan seperangkat data yang sama, tapi akan menghasilkan
nilai penduga a dan Th yang berbeda seperti terlihat di atas. Perbedaan ini
disebabkan oleh perbedaan asumsi dari kedua persamaan tersebut. Persamaan
cakram mengasumsikan banyaknya inang selalu tetap, dan pola pencarian inang
dilakukan secara sistematik yaitu inang yang sudah diparasit tidak akan
dikunjungi lagi oleh parasitoid. Persamaan acak mengasumsikan jumlah inang
berkurang sejalan dengan waktu karena sebagian telah terparasit, serta pola
pencarian inang dilakukan secara acak. Sebagai akibatnya, nilai penduga a dan
Th yang dihitung dengan persamaan acak lebih tinggi daripada yang dihitung
dengan persamaan cakram.
Persamaan cakram lebih tepat diterapkan untuk parasitoid yang mampu
membedakan status inang yaitu apakah inang telah terparasit atau belum,
sedangkan persamaan acak untuk parasitoid yang tidak memiliki kemampuan
membedakan status inang (Wang & Ferro 1998). Dalam penelitian selama ini
belum pernah dijumpai adanya fenomena superparasitisme pada parasitoid

55
A. dasyni. Hal ini diduga berkaitan dengan kemampuan parasitoid A. dasyni
untuk mengenali telur inang yang telah terparasit. Bila demikian halnya, maka
persamaan cakram mungkin lebih tepat digunakan untuk memerikan tanggap
fungsional parasitoid A. dasyni terhadap telur kepik lada.
Pengaturan populasi hama di alam dapat terjadi karena adanya mekanisme
terpaut kerapatan yang bersumber dari tanggap fungsional dan tanggap numerik.
Tanggap fungsional tipe III bersifat terpaut kerapatan pada selang kerapatan inang
tertentu, sehingga diperkirakan mampu memelihara kesetimbangan populasi.
Sementara tanggap fungsional tipe II bersifat terpaut kerapatan berkebalikan
(inverse density-dependent), yaitu proporsi inang yang terparasit menurun dengan
meningkatnya kerapatan inang. Keadaan ini dapat menciptakan ketidakstabilan
populasi. Hasil penelitian mendapatkan bahwa parasitoid A. dasyni
memperlihatkan tanggap fungsional tipe II, sehingga diperkirakan tidak mampu
mengatur populasi inangnya (Hassell 2000). Oleh karena itu, agar terjadi
kestabilan populasi, tanggap fungsional ini perlu dilengkapi dengan tanggap
numerik melalui pelepasan inundasi parasitoid.
Kesimpulan
Parasitoid A. dasyni memperlihatkan tanggap fungsional tipe II terhadap
peningkatan kerapatan inang. Berdasarkan nilai koefisien determinasi, persamaan
cakram dan persamaan acak sama sesuainya untuk memerikan tanggap fungsional
parasitoid. Jumlah maksimum telur inang yang dapat diparasit menurut
persamaan cakram dan persamaan acak berturut-turut 22 dan 12 butir per hari.
Karena parasitoid diduga mampu membedakan telur yang sudah diparasit dengan
yang belum, persamaan cakram mungkin lebih tepat untuk digunakan.
Daftar Pustaka
Collins MD, Ward SA, Dixon AFG. 1981. Handling time and the functional response of Aphelinus thomsoni, a predator and parasite of the aphid Drepanosiphum platanoidis. J Anim Ecol 50:479-487.
Deciyanto S, Trisawa IM, Muchyadi. 1993. Parasitism fluctuation of egg-parasitoids of pepper bug (Dasynus piperis China) in Bangka. J Spice Medic Crops 1(2):33-36.

56
Hassell MP. 2000. Host-parasitoid population dynamics. J Anim Ecol 69:543-566.
Holling CS. 1959. Some characteristics of simple types of predations and parasitism. Canad Entomol 91:385-398.
Holling CS. 1965. Functional response of predators to prey density and its role in mimicry and population regulation. Mem Entomol Soc Can 45(1):1-60.
Houck MA, Strauss RE. 1985. The comparative study of functional responses: experimental design and statistical interpretation. Can Entomol 115: 617-629.
Juliano SA. 1993. Non-linear curve fitting: predation and functional response curves. Di dalam: Scheiner SM, Gurevitch J, editor. Design and analysis of ecological experiments. New York: Chapman & Hall. hlm 158-183.
Laba IW, Kilin D, Trisawa IM. 2004. Tingkat kerusakan dan serangan hama buah lada, Dasynus piperis China pada pertanaman lada di Bangka. J Entomol Indonesia 1(1):34-40.
Messina FJ, Hanks JB. 1998. Host plant alters the shape of the functional response of an aphid predator (Coleoptera: Coccinellidae). Environ Entomol 27:1196-1202.
Parella MP, Horsburgh RL. 1983. Functional responses of the black hunter thrips, Leptothrips mali (Thysanoptera: Phlaeothripidae), to density of Panonuchus ulmi (Acari: Tetranychidae). Environ Entomol 12:429-433.
Pervez A, Omkar. 2005. Functional responses of coccinellid predators: an illustration of a logistic approach. J Insect Sci 5(5):1-6.
Rogers DJ. 1972. Random search and insect population models. J Anim Ecol 41: 569-583.
SAS Institute. 1990. SAS User’s guide. Ver 6 Ed 4 vol II. Cary (North Carolina): SAS Institute Inc.
Schenk D, Baker S. 2002. Functional response of a generalist insect predator to one of its prey species in the field. J Anim Ecol 71:524-531.
Solomon ME. 1949. The natural control of animal populations. J Anim Ecol 18: 1-35.
Suprapto, Thomas. 1989. Aspek biologi pengisap buah lada pada berbagai tingkat umur buah. Pembr Littri 14(4):119-125.
Trisawa IM, Rauf A, Kartosuwondo U. 2007. Biologi parasitoid Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae) pada telur Dasynus piperis China (Hemiptera: Coreidae). Hayati 14(3):81-86.

57
Wang B, Ferro DX. 1998. Functional responses of Trichogramma ostriniae
(Hymenoptera: Trichogrammatidae) to Ostrinia nubilalis (Lepidoptera: Pyralidae) under laboratory and field condition, Environ Entomol 27(3): 752-758.