properties of glutamate dehydrogenase in developing legume...
TRANSCRIPT

Pertanika 6(1),63-66 (1983)
Properties of glutamate dehydrogenase in developing legume fruit. II.The effects of divalent cations.
M.MARZIAHfabatan Biokimia dan Mikrobiologi, Fakulti Sains dan Pengajian Alam Sekitar,
Universiti Pertanian Malaysia, Serdang, Selangor, Malaysia.
Key words: Glutamate dehydrogenase, developing seeds; legume; manganese.
RINGKASAN
Aktiviti glutamate dehydrogenase dari ekstrak kasar bijian Lupinus anguistifolius yang direncatoleh EDTA boleh dipulih kembali samada separa atau sepenuhnya oleh kation-kation berdwivalensi yangditambah ke dalam campuran pengeraman. Mn2+ dan Ca 2
rr memberi rangsang yang tinggi sekali masingmasing 67% dan 66% dari kontrol, diikuti oleh Zn2+ (59%), Co2+ (55%), Mg2+ (53%) dan Cu2+ (49%).Enzim yang tak cukup manganum adalah lebih sensitif terhadap manganum dan lebih teransang padakepekatan manganum yang rendah dari enzim yang cukup manganum. Tetapi enzim yang tak cukup manganum mencapai aktiviti maksima sebanyak 91 % pada 1.4 mM Mn 2 + sementara enzim yang cukup manganummencapai aktiviti maksima 105% pada 2.0 mM Mn2+.
SUMMARY
The activity of EDTA-inhibited glutamate dehydrogenase from a crude seed extract of Lupinusangustifolius could be partially or fully restored by the addition of divalent cations to the incubationmixture. Mn2+ and Ca2+ gave the highest stimulations of 67% and 66% of the control, respectively; followedby Zn2+ (59%), Co2+ (55%), Mg2+ (53%) and Cu 2
+ (49%). The Mn-deficient enzyme was more sensitive tomanganese and more stimulated at a lower manganese concentration than the Mn-sufficient enzyme.However, Mn-deficient enzyme achieved a maximum activity of 91% at 1.4mM Mn2+ while Mn-sufficientenzyme achieved a maximum activity of 105% of control at 2.0mM.
INTRODUCTION
The importance of glutamate dehydrogenase(GDH) in amino acid metabolism especially indeveloping legume seeds has been widely emphasised. One of the interesting characteristicsof GDH is its response to certain metals. TheEDTA-inactivated GDH can either· can be fullyor partially reactivated by several metal ions, andGDH from different plant sources seem to respondto differe:lt metals (Pahlich and Joy, 1971);Yamasaki and Suzuki, 1969; Sheid et ai., 1980).
In the previous paper, a detailed study ofGDH inhibition by EDTA was reported. Thefollowing study was conducted to investigatethe effects of metal ions on GDH activity fromdeveloping seeds of Lupinus angustifolius. Knowledge on the metal ion response by the enzymewill provide a useful parameter in studying metalrequirements in plants and perhaps the interactionof different metals, especially the trace elements.
63
MATERIALS AND METHODS
General procedure
Plant materials and chemicalsMn-deficient seeds were obtained from plants
grown where no manganese fertilisers were appliedand the soil is inherently deficient in manganese.Similarly, the Mn-sufficient seeds from plantsgrown from plots with added manganese fertilisers.
The use and supply of chemicals were as inthe previous paper (Marziah, 1982).
Experimental detailsThe crude extract preparations and the
enzyme assays have been described elsewhere(Marziah, 1982). In order to prevent contamination from exogenous metalic cations all buffersand solutions were treated with Chelex-IOO.Glassware was soaked and washed with 0.2MEDTA and thoroughly rinsed with dou ble-deionisedwater.

M. MARZIAH
Crude Enzyme Studies
The effects of metal ions on GDH activityThe crude enzyme extract was preincubated
with ImM EDTA and the respective metal ionsfor 30 min. The final concentration of metal ions,used as chlorides and adjusted to pH 7.0 in thereaction mixture, was ImM.
Stimulation of GDH activity by manganeseThe stimulation studies were carried out both
with EDTA-treated and Chelex-treated enzymeextracts, Mn2+ was preincubated with the enzymeextracts for 3CJ min. before NH4 Cl was added toinitiate the reaction.
Responses of GDH activities in Mn-deficient andMn-sufficient seed extract to manganese.
The manganese concentration used rangedfrom 0.6mM - 3.0mM. The study was made onEDTA-preincubated enzyme extracts.
Mn2+ and Ca2 + seemed to activate the enzymefrom both the crude and purified extracts of mostof the tissues studied (Fawole and Boulter, 1977,Chou and Splitstoesser, 1972, Ehmke andHartmann, 1978).
The stimulation of GDH activity by Mg2 +,Co2+ and Zn2 + has also been reported (Yamasakiand Suzuki 1969). Chou and Splitstoesser (1972)reported full GDH stimulation by Cu2 + in apumpkin cotyledon extract.
In Table 2, NADH-GDH activity fromEDTA-pretreated enzyme was greatly stimulatedby Mn 2 +. The activity was 76% of the controlwhen the EDTA-treated enzyme was incubatedwith ImM Mn2+. A higher Mn2+ concentrationcaused inhibition. It is interesting to note that theNADPH-activity was not stimulated by Mn 2 +, orby other metal ions examined by earlier workers(Fawole and Boulter, 1977; King and Wu, 1970).
Table 2The effects of manganese concentrations on GDH
activities on EDTA-pretreated enzyme extract fromseeds of L. angustifolius cv. UniCrop.
Activity(Expressed as%of control)
RESULTS AND DISCUSSION
NADH-GDH could be partially activated bymetal ions (Table I) maximum stimulation wasachieved with Mn2+ (67%) and Ca2+ (66%) and to alesser extent Zn2+ (59%), Co2+ (55%) and Mg2+(53%). Cu 2 + caused stimula.tion of only 49%. !heresults were consistent wIth those of prevIOUSworkers who reported that GDH was activated bydivalent cations. (Ehmke and Hartmann, 1976;1978' Fawole and Boulter, 1977; King and Wu,1971: Pahlich and Joy, 1971; YanIasaki andSuzuki 1969).
Treatment(Final concentration)
NADHGDH
NADPHGDH
Table 1The effects of divalent cations on NADH-DDH
activity in the seeds of L. augustifolius cv. UniCrop.
Treatment(Final concentration)
NADH-DDHactivity
(Expressed as% of control)
~~rol 100
Control + 1.0 mM EDTA 6
0.25 mM Mn2+ + 1.0 mM EDTA 36
0.50 mM Mn2+ + 1.0 mM EDTA 58
1.00 mM Mn2+ + 1.0 mM EDTA 76
2.00 mM Mn2+ + 1.0 mM EDTA 67
3.00 mM Mn2+ + 1.0 mM EDTA 24
100303033
3034
30
Control
Control + 1.0 mM EDTA
1.0 mM Mn2+ + 1.0 mM EDTA
1.0 mM Ca2+ + 1.0 mM EDTA
1.0 mM Zn2+ + 1.0 mM EDTA
1.0 mM Co2+ + 1.0 mM EDTA
1.0 mM Mg 2+ + 1.0 mM EDTA
1.0 mM Cu2+ + 1.0 mM EDTA
100
20
67
66
59
55
53
49
64
Mn2+ was supplied as chloride and was pre-incubatedwith the EDTA-pretreated enzyme extract for 30 min.before reaction was initiated.
The Chelex-treated enzyme, however, wasstimulated by more than 100% by all the Mn2+concentrations used (Table 3). The marked increaseof the enzyme activity was probably not only dueto the stimulation effects of metal ions but alsodue to the removal of divalent cationic inhibitorsas proposed earlier.
GLUTAMATE DEHYDROGENASE IN DEVELOPING LEGUME FRUIT. II.
Table 3The effects of manganese concentrations on
NADH-GDH activity (after Chelex-treated) obtainedfrom seed extract of L. angustifolius cv. UniCrop.
Treatment(Final concentration)
NADH-GDH activity(Expressed as%of control
ions was added to the extraction medium orto the pre-assay mixture. It is highly likely that theGDH activity would be higher than what wasobtained in the control if the addition of metalions, such as Mn2 + or Ca2 +, were added. Generally,it appears that any metal that is present in theGDH molecule is not easily dissociable and consequently a higher concentration of EDTA isrequired to inactivate the enzyme.
Mn2+ was supplied as chloride and was pre-incubatedwith Chelex-treated enzyme extract for 30 min.
It should be emphasised that in all the metalion activation studies, no Mn 2 + or other metal
Joy (1969) first proposed that Ca2+ is involved in the aggregation or disaggregation of theenzyme. Ehmke and Hartmann (1976; 1978proposed that the metal ion Ca'+, governs onlythe NADH-reaction involving an equilibrium oftwo forms, and this is achieved by the relativeiyweak bonds between Ca2+ and the enzyme. Theyfurther suggested that the removal of Ca2+ andthe addition of Ca2+ may involve the alterationin the binding character or binding requirementsof the substrate NH;. Sheid et al., (1980) suggested that the role of Ca2 + is in governing areversible equilibrium between catalytically activeand inactive forms. It is likely that Mn 2 + and othermetal ions may be involved in similar mechanismsas proposed by Sheid et al., (1980).
120
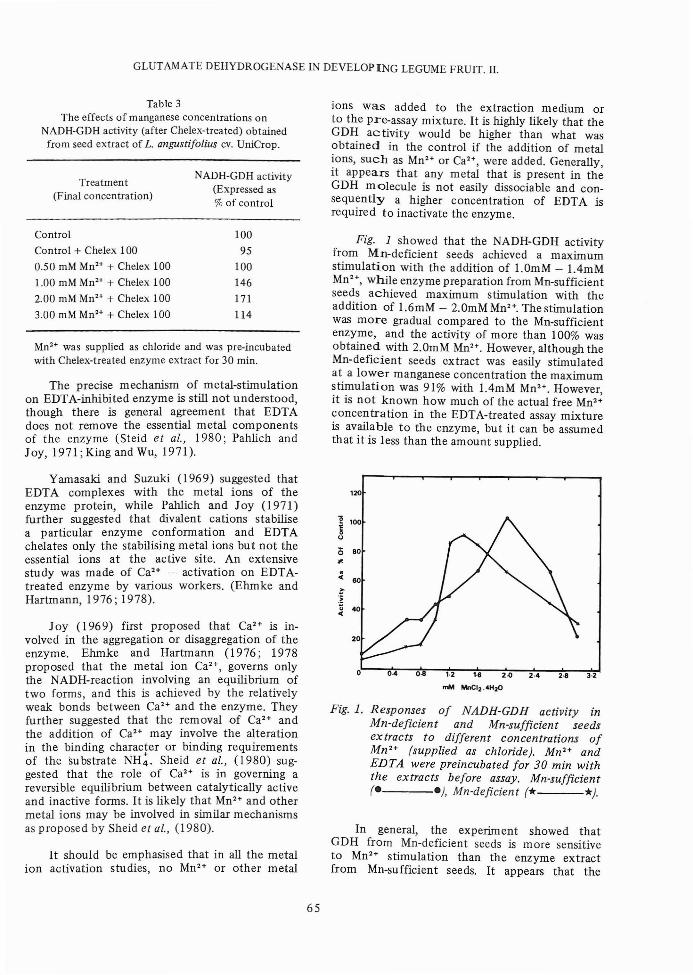
Fig. 1 showed that the NADH-GDH activityfrom Mn-deficient seeds achieved a maximumstimulation with the addition of 1.0mM - 1.4mMMn 2 +, while enzyme preparation from Mn-sufficientseeds achieved maximum stimulation with theaddition of 1.6mM - 2.0mM Mn2+. The stimulationwas more gradual compared to the Mn-sufficientenzyme, and the activity of more than 100% wasobtained with 2.0mM Mn2+. However, although theMn-deficient seeds extract was easily stimulatedat a lower manganese concentration the maximumstimulation was 91% with 1.4mM Mn2 +. However,it is not known how much of the actual free Mn2+concentration in the EDTA-treated assay mixtureis available to the enzyme, but it can be assumedthat it is less than the amount supplied.
O,L-_~..r--;:O.T---;1;':;'2~""'1;';;'8--t2~;;-~2.~4--:;2~-8-""'3;';-l.2
- MnCI2·4HzO
..i<.>
In general, the experiment showed thatGDH from Mn-deficient seeds is more sensitiveto Mn2+ stimulation than the enzyme extractfrom Mn-sufficient seeds. It appears that the
Fig. 1. Responses of NADH-GDH activity inMn-deficient and Mn-sufficient seedsex tracts to different concentrations ofMn2+ (supplied as chloride). Mn 2 + andEDTA were preincubated for 30 min withthe extracts before assay. Mn-sufficient(e e), Mn-deficient (* *).
100
95100
146
171114
Control
Control + Chelex 100
0.50 mM Mn2+ + Chelex 100
1.00 mM Mn2+ + Chelex 100
2.00 mM Mn2+ + Chelex 100
3.00 mM Mn2+ + Chelex 100
The precise mechanism of metal-stimulationon EDTA-inhibited enzyme is still not understood,though there is general agreement that EDTAdoes not remove the essential metal componentsof the enzyme (Steid et at., 1980; Pahlich andJoy, 1971; King and Wu, 1971).
Yamasaki and Suzuki (1969) suggested thatEDTA complexes with the metal ions of theenzyme protein, while Pahlich and Joy (1971)further suggested that divalent cations stabilisea particular enzyme conformation and EDTAchelates only the stabilising metal ions but not theessential ions at the active site. An extensivestudy was made of Ca2 + - activation on EDTAtreated enzyme by various workers. (Ehmke andHartmann, 1976; 1978).
65

M. MARZIAH
Mn-deficient enzyme, because of the low manganese availability in the extract, has to be efficientin its utilization or requirement of MnH. Thecase of stimulation at lower manganese concentration shows that the seeds that are Mn-deficienthave conditioned themselves to a low manganeseavailability, thus keeping its requirement to theminimum. Full stimulation of the enzyme activity,however, was not achieved because of the limitedMnH supply. The situation was not true inMn-sufficient seeds.
CONCLUSION
The EDTA-inactivated NADH-GDH in a crudeextract from developing lupin seeds was highlystimulated by MnH and CaH followed by Zn2 +,COH and MgH. The NADPH-GDH, however, wasnot at all stimulated. The Mn-deficient enzymeappeared to be more sensitive and stimulated thanthe Mn-sufficient enzyme. It is highly likely thatthe Mn-deficient enzyme has conditioned itselfto low manganese availability and it is moreefficient in its usage of manganese.
REFERENCES
EHMKE A. and HARTMANN T. (1976): Properties ofglutamate dehydrogenase from Lemna minor.Phytochem. 15: 1611-1617.
66
EEMKE A. and HARTMANN T. (1978): Control ofglutamate dehydrogenase from Lemna minor bydivalent metal ions. Phytochem. 17: 637-64l.
F AWOLE M.O and BOULTER D. (1977): Purificationand properties of glutamate dehydrogenase fromVigna unguiculata (L) Welp. Planta, 134: 97-102.
JOY K.W. (1969): N Metabolism of Lemna minor.Plant. Physiol. 44: 845-848.
KING J. and WU W.Y.F. (1971): Partial purification andkinetic properties of glutamate dehydrogenase fromsoybean cotyledons. Phytochem.. 10: 915-928.
MARZIAH, M (1982): Properties of glutamate dehydrogenase in developing legume fruit. 1. Effects ofChelating Agents. Pertanika. 5(2): 154-159.
P AHLICH E. and JOY K.W. (1971): Glutamate dehydrogenase from pea roots: Purification properties ofthe enzyme. Can. J. Biochem. 49: 127-138.
SHEID H., EHMKE A. and HARTMANN T. (1980):Plant NAD-dependent glutamate dehydrogenase.Purification, molecular properties, metal ion activation of the enzymes from Lemna minor and Pisumsativum. Z. Natur. 350: 213-221.
YAMASAKI K. and SUZUKI Y. (1969): Some propertiesof glutamate dehydrogenase from pea seedlings.Phytochem. 8: 963-969.
(Received 25 February 1982)