prey selection by larval micromus tas'lnq,niae walker ... · larva m. tasmaniae menangkap...
TRANSCRIPT

Pertanika 8(3), 359 - 363 (1985)
Prey Selection by Larval Micromus tas'lnQ,niae Walker(Neuroptera: Hemerobiidae)
M.Y. HUSSEIN*Plant Protection Department,
iFaculty of AgricultU1;e,Universitz' Pertanian Malaysia,Serdang, Selangor, Malaysia.
Key words: Micromus tasmaniae; hemerobiidae; predation
ABSTRAK
Micromus tasmaniae ialah sejenis serangga pemangsa yang palz"ng kerap didapati sebagaimusuh semula jadi kepada kutu daun ubi kentang di Selatan Australia. Kemungkinan keupayaanlarva M. tasmaniae menangkap setiap instar kutu daun, Myzus persicae Sulzer, telah diukur padasuhu tetap 15°e, 20 0 e dan 25°e. Kemungkin(J,n keupayaan menangkap M. persicae oleh M.tasmaniae meningkat selepas setz"ap kali bersalin kulz"t dan menurun sedikit sejurus sebelum bersalinkulz"t terutama sekali pada suhu 15°e. Kecuali bagi larva yang terlalu kecil dan remaja, larvapemangsa berupaya menangkap dengan cekap mangsa yang kecil atau besar. Larva yang remajamenunjukkan sedikit taMat pemilz"han terhadap mangsa yang kecil, manakala larva yang lebih tuatz"dak menunjukkan pemilz"han langsung. Keberkesanan larva M. tasmaniae mungkin berbeza diantara musim dan iklz"m.
ABSTRACT .The brown lacewing, Micromus tasmaniae, is the most common predator of potato aphids in
South Australia. The probability of capture and prey preference of larval M. tasmaniae weremeasuredfor each instar of Myzus persicae Sulzer (the green peach aphid) at constant 15°e, 20° and25°e. The probabzUty of capture ofM. persicae by larval M. tasmaniae increased after each moultand declined just before moulting, especially at 15°e. Except for very small and young larvae, thepredator larvae were able to capture efficiently both small and large prey. Young larval predatorsshowed some preference for smaller prey~ whereas older larvae exhibited lz"ttle or no preference. Theeffectz"veness ofM. tasmaniae larvae may vary considerably between seasons and clz"mates.
INTRODUCTION
The Australian brown lacewing, Micromustasmaniae Walker is a common and abundantpredator of Myzus persicae (Sulzer) and otherpotato aphids in South Aus~ralia (Hussein,1982). Numerical changes in the populations ofM. persicae and M. tasmaniae have been studiedover a 2-year period at Milang and Adelaide,South Australia by Hussein (1982). Field ~ata
are difficult to interpret without understanding
the characteristics of the predator-prey interaction to provide guidance for the biologicalcontrol of M. persicae on potatoes. Field samplings of eggs, larvae and adults of M. tasmaniaeindicated that the brown lacewings were presentthroughout the year even when prey populationswere very low. Its ability to survive while mostother predators like the coccinellids, sryphidsand chrysopids fail, signifies that M. tasmaniaeprobably has high probability of capturing itsprey and has no preference for prey size or age.
• Part of the author's Ph.D. thesis submitted to the University of Adelaide, Adelaide, South Australia.
M.Y. HUSSEIN
To help interpret the field observations,additional information was obtained in laboratory experiments on the probability ofcapture, the prey preference by larval M.tasmaniae and the influence of temperature onthe predator's activity.
MATERIALS AND METHODS
An experiment was conducted in October1981 at Waite Agricultural Research Institute,Glen Osmond, South Australia using predatorsand aphids from their insectary cultures. It wasconducted at constant temperatures of 15, 20and 25°C under LD 12 : 12, and comprised fourtreatments of different numbers of M. persicaeinstars offered to each predator larva every daythroughout the experiment (Table 1).
The predator larvae were used in theexperiment within 24 hours of the eggs hatching,and the aphids were fed to them on potato leafdiscs (3 cm in diameter) inside small cages asdescribed by Hussein (1982). There were 4 larvae(replicates) at each treatment but because oflimited resources the experiment was run as 2blocks in time with 2 replicates in each block. Onanyone day, the aphids to be given in the different treatments were randomized amongst thepredator larvae.
Table 1 shows that during its first larvalinstar, each predator was given 15 aphids each
day; and in its second and third instar, eachlarva was given 35 aphids. The ratios of thenumbers of different aphid instars given as preywere converted to percentages of the total preypresented. The proportion of any aphid instareaten on one day was similarly estimated bytotalling the numbers of that instar eaten byeach of the 8 larvae in the 4 treatments at eachtemperature and dividing the total then by thetotal number of prey presented. The proportionswere then expressed as percentages.
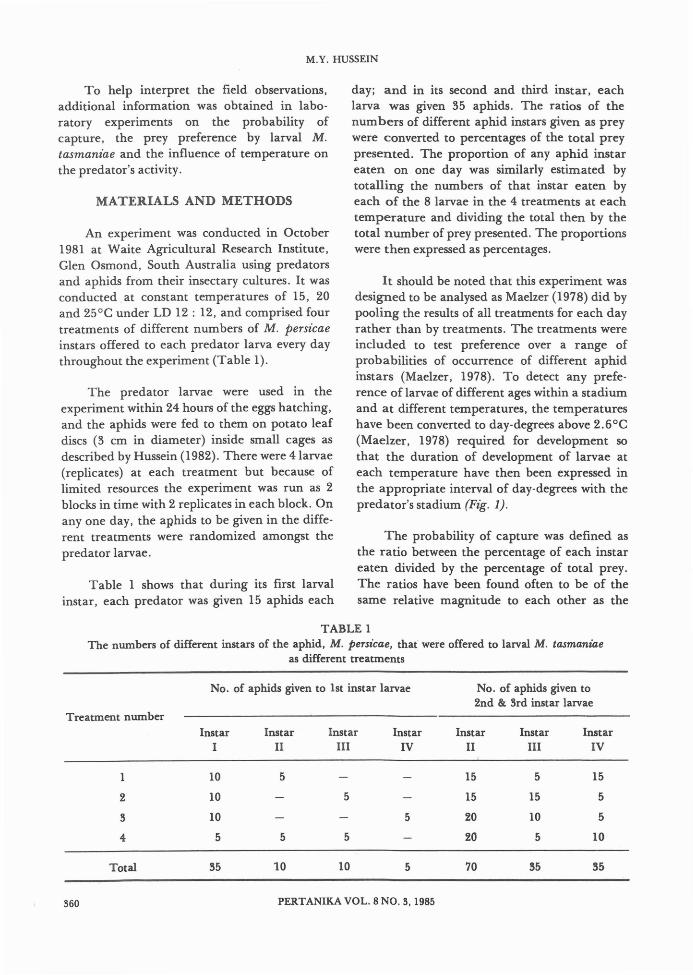
It should be noted that this experiment wasdesigned to be analysed as Maelzer (1978) did bypooling the results of all treatments for each dayrather than by treatments. The treatments wereincluded to test preference over a range ofprobabilities of occurrence of different aphidinstars (Maelzer, 1978). To detect any preference of larvae of different ages within a stadiumand at different temperatures, the temperatureshave been converted to day-degrees above 2.6°C(Maelzer, 1978) required for development sothat the duration of development of larvae ateach temperature have then been expressed inthe appropriate interval of day-degrees with thepredator's stadium (Fig. 1).
The probability of capture was defined asthe ratio between the percentage of each instareaten divided by the percentage of total prey.The ratios have been found often to be of thesame relative magnitude to each other as the
TABLE 1The numbers of different instars of the aphid, M. persicae, thai were offered to larval M. tasmaniae
as different treatments
No. of aphids given to 1st instar larvae No. of aphids given to2nd & grd instar larvae
Treatment numberInstar Instar Instar Instar Instar Instar Instar
I II III IV II III IV
1 10 5 15 5 15
2 10 5 15 15 5
g 10 5 20 10 5
4 5 5 5 20 5 10
Total g5 10 10 5 70 55 55
360 PERTANlKA VOL. 8 NO.3, 1985
PREY SELECTION BY LARVAL MICROMUS TA SMA NIAE WALKER
AlI'idNLDer-..... til 20 JD 40 10 .. 711 .. .. IlO iii l1il db '"
Hg. 1. ProbabzUties of capturing different instars ofM. persicae by larvae of M. tasmaniae atthree different temperatures. Arrows indicatethe end of the 3 larval stadia.
probabilities of capture estimated by Chesson(1978) and Maelzer (1978). For larvae to showpreference for certain prey instars, the percentages of prey eaten should be greater or less thanthe percentages of prey presented (Maelzer,1978).
It is well known that the previous experienceof a predator may affect hunger levels which inturn may cause significant relative changes inthe probability of capture (Hassell, 1976). Thismay be true when experiments are conductedover a very short period, say a few hours or so, asdid Dixon (1959) and Wratten (1973). However,in this study, estimates of relative probabilities ofcapture of M. persicae of different instars of M.tasmaniae were obtained over much longerperiods than those used to estimate probabilitiesof capture by successful encounters (Dixon,1959; Wratten, 1973; Chesson, 1974). Thelonger periods included periods of hunger andnon-hunger, so the hunger level is not likely tohave caused complications.
RESULTS
Probability ofCapture ofPrey
Figure 1 shows that at each temperature,the trend in the probability of capture of M.
persicae by larvae of M. tasmaniae increased to apeak after each larval moult and declined justbefore moulting. Similar trends have beenobserved by Dixon (1959) and Wratten (1973)with a coccinellid predator AdaNa bipunctata.In this study, the trend was most obvious at15°C. The apparent decreases in the probabilityof capture (at the day-degrees indicated by thearrows in Fig. 1) correspond to the decreases inslope of the larval growth and voracity curve.Again, these decreases are most obvious at 15°C.The data also indicate that the efficiency of M.tasmaniae larvae in capturing M. persicae increases with age and with each moult. Similarresults have been obtained for coccinellidpredator on sycamore aphids (Dixon, 1959 and1970), wheat aphid (Brown, 1972), lime aphids(Wratten, 1973) and rose aphid (Chesson, 1974;Maelzer, 1978), but the methods of estimatingprobability of capture were different. Dixon(1959 and 1970) also found that A. bzpunctataand A. decempunctata captured more smallaphids than large ones. However, in this study,except during the early stages of larval development, larvae of M. tasmaniae seemed to be ableto capture efficiently both small and large M.persiae. The lower efficiency of Adalia larvae incapturing large prey was because of effectiveescape responses the prey possessed such as kicking away the larvae when encountered (Wratten,1973).
The influence of temperature on the probability of capture by larval M. tasmaniae isshown also in Figure 1. The probability ofcapture of prey increases with temperatureprobably because of increased larval growth athigher temperatures. So too, the probability offirst instar of the aphid Macroszphum rosaebeing captured by second instar of the coccinellid species, Leis conformis increased markedly at 25°C (Chesson, 1974). The increasedvoracity with temperature recorded for coccinellid predators by Dunn (1952), Sundby (1966),Maelzer (1978) and others is probably due partlyat least, to increased probability of captureresulting from greater mobility of predatorlarvae as temperature increases. With Chrysopacarnea (Neuroptera: chrysopidae) larvae,
PERTANIKA VOL. 8 NO.3, 1985 361
M.Y. HUSSEIN
Sundby (1966) found 25% more" aphids wereconsumed at 21°C than at 16°C.
which shows very little or no preference for preysize is more likely to be a better candidate forbiological control of aphids.
Preference for P1'ey
.
DISCUSSION
Quite recently, hemerobiids have beenshown to have good potential for biologicalcontrol of aphids (Neuenschwander, 1975;Neuenschwander and Hagen, 1980; Hussein,1982). With respect to the feeding habits,hemerobiids are among the least voracious whencompared to coccinellids, syrphids and chrysopids (Neuenschwander et al., 1975). However,M. tasmanz"ae has. some advantages which arecommon to most hemerobiids over the otherspecies of aphid predators. The larvae of M.tasmaniae have a higher probability of capturingprey and are most efficient at low prey densitywhich is very useful in the control of aphids asa vector of virus diseases of crop plants.
The abundance of Hemerobz"us pacifz"cus(Neuroptera: Hemerobiidae) and M. tasmanz"aewas studied by Neuenschwander (1975) andHussein (1982), respectively. Neuenschwander etal. (1975), found in Coastal California, USA,numbers of adult H. pacifz"cus were higher inJanuary-February while in the valley, adultsresponded to the late spring (May) peak of theaphids occurring in the alfalfa fields. The lowernumbers of H. pacifz"cus adults in the valley wasattributed to heavy mortality suffered by theeggs, larvae, and pupae under high summer andearly autumn temperatures occurring fromJuly - September. Comparison between thephenologies of H. pacifz"cus in the two areassuggest that the coastal areas constitute a stablezone of permanent occupancy (Huffaker andMessenger, 1964), where the adult predatorsshow a high rate of aphid-density dependence.The abundance of M. tasmanz"ae and possiblyother hemerobiids under cooler conditions in thefield may be because they can be reproductivelyactive at extremely low temperatures (- 4°C)that is sufficient for the normal development oftheir immatures (Neuenschwander, 1975; Syrettand Penman, 1981; Hussein, 1982).
2O'C.' • -- . • . ...,,'c
• .. . • - • . •64& .. r •
+3:1 • 1..-__2S
-·-
C
......_"'l ----."".r---20~
!60~~ JO_ U oL.-_...L.-..:.l-_.L..._::L----'_..L-.----L.--J'-----'---I.._~--"-_
n:::1 I..
;I:l---.L..-..:J----I.--=--L..-..:;J..~.::J...._.J~--'-_'___.L-.J....~.......
I;JJ.~·-~~--JL---&-~
AjIIlid~ 'a •• la •• ,a •• , •••.oa•••••• io. a •••• u
Dor-..... • 20 3D 40 SO III 70 III 10 100 III 120 UO llD
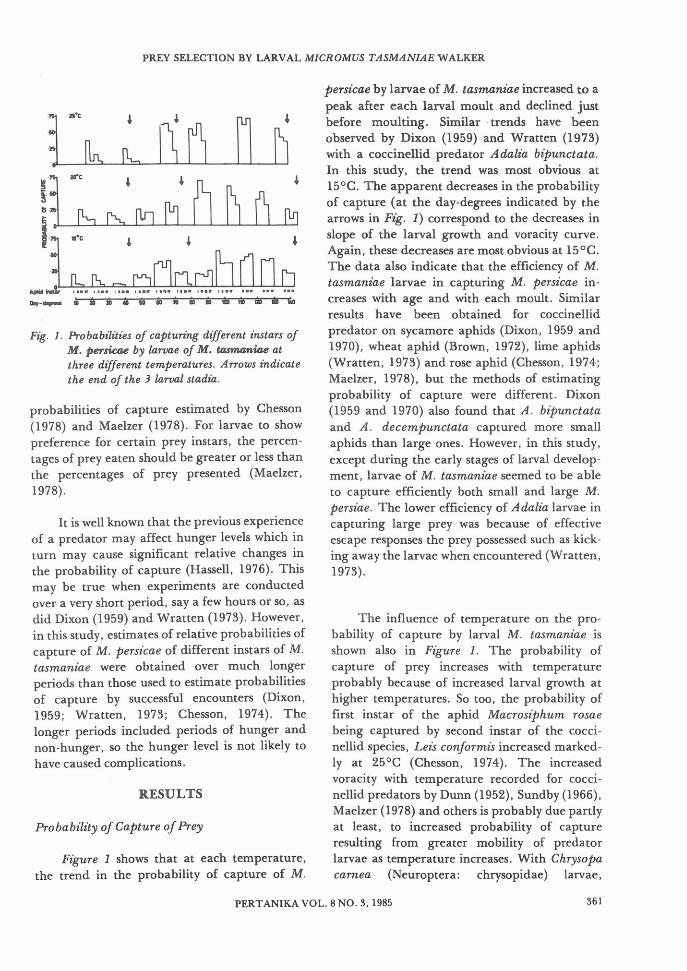
F£g. 2. Percentages of different instars ofM. persicae presented (0) and of thedifference from expected (_) (i. e. thedifferen'ce between percent presented andeaten by larvae of M. tasmaftiae) at threedifferent temperatu'res.
Whether or not the predator has a preference for some particular aphid instar willhave some influence on the change in size of preypopulations within which the predator is presented with a choice between more than one agegroup of prey. A predator like M. tasmanz"ae
Ftgure 2 illustrates the preference for eachaphid instar as percentage differences fromexpected (solid black areas) in relation to thepercentages of prey of each instar that werepresented to the predators. Young predatorlarvae showed some preference for smaller preyas illustrated by positive percentage differences,whereas older larvae of M. tasmanz"ae exhibitedno preference. Temperature seemed to have verylittle or no influence on prey preference. Bycontrast, studies on coccinellid predators haveshown that larvae changed their preference fordifferent aphid instars from day to day (Dixon,1959; Wratten, 1973; Maelzer, 1978), thechange being attributed to different probabilities of capture (Chesson, 1978).
362 PERTANIKA VOL. 8 NO.3, 1985

PREY SELECTION BY LARVAL MICROMUS TASMANIAE WALKER
ACKNOWLEDGEMENTS
The author wishes to thank the Waite Agricultural Institute, University of Adelaide, SouthAustralia and the Australian Universities International Development Programmes for fundingthe research.
REFERENCES
CHESSON, J. (1974): Selection predation. B. Ag. Sc.(Hons.) Thesis, University of Adelaide, Adelaide,South Australia. 52p.
CHESSON, J. (1978): Measuring preference in selectivepredation. Ecology. 59: 211- 215.
DIXON, A.F.G. (1959).: An experimental study of thesearching behaviour of the predatory coccinellidbeetle Adalla decempunctata.]. Anim. Ecol. 28:259 - 281.
DIXON, A.F.G. (1970): Factors limiting the effectiveness of the coccinellid beetle, A daNa bzpunctata(L.) as a predator of the sycamore aphid,Drepanoszphum plantanoides (Schr.).]. Anim.Ecol. 39: 739 -751.
DUNN, J.A. (1952): The effect of temperature on thepea aphid-ladybird relationship. Rep. Natn.Veg. Res. Sta. 1951, pp. 21- 23.
HASSELL, M.P. (1976): The dynamics of competitionand predation. Studies in BioI. No. 72, EdwardArnold. 68 p.
HUFFAKER, C.B. and MESSENGER, P.S. (1964): Theconcept and significance of natural control. In:Biological Control of Insect Pests and Weeds. Ed.P. DeBach. Reinhold Pub., New York. 844 p.
HUSSEIN, M. Y. (1982): The effects of natural enemiesof Myzus persicae Sulzer upon its populationtrends in potato crops in South Australia. Ph.D.Thesis. University of Adelaide, Adelaide, SouthAustralia. 257 p.
MAELZER, D.A. (1978): The growth and voracity oflarvae of Leis conformis (Boisd.) (Coleoptera:Cotcinellidae) fed on the rose aphid Macroszphum rosae (L.) (Homoptera: Aphididae) in thelaboratory. Aust.]. Zool. 26: 293 - 304.
NEUENSCHWANDER, P. (1975): Influence of temperature and humidity. on the immature stages, ofHemerobius pacificus. Environ. Entomol. 4:215 - 220.
NEUENSCHWANDER, P., HAGEN, K.S. and SMITH, R.F.(1975): Predation on aphids in California's alfalfafields. Hilgardia. 43: 53 - 78.
NEUENSCHWANDER, P. and HAGEN, K.S. (1980): Roleof the predator H emero bius pacificus in a noninsecticide treated artichoke field. Environ.Entomol. 9: 492 - 495.
SUNDBY, R.A. (1966): A comparative study ')n theefficiency of three predatory insects - Coccinellaseptempunctata (Coleop: Coccinellidae),Chrysopa carnea (Neuroptera: Chrysopidae) andSyrphus ribesii (Diptera: Syrphidae) at two different temperatures. Entomophage. 11: 395 - 404.
SYRETT, P. and PENMAN, D.R. (1981): Developmentalth~eshold temperatures for the brown lacewing,Mzcromus tasmaniae (Neuroptera: Hemerobiidae). N.Z.]. Zool. 8: 281- 283.
WRATTEN, S.D. (1973): The effectiveness of thecoccinellid Adalia bzpunctata (L.) as a predatorof the lime aphid, Encallzpterus tilliae L. ].Anim. Ecol. 42: 785 - 802.
(Received 1June, 1985)
PERTANIKA VOL. 8 NO.3, 1985 363