research article reversal of ampicillin resistance in mrsa...
TRANSCRIPT

Research ArticleReversal of Ampicillin Resistance in MRSA via Inhibition ofPenicillin-Binding Protein 2a by Acalypha wilkesiana
Carolina Santiago Ee Leen Pang Kuan-Hon Lim Hwei-San Loh and Kang Nee Ting
Faculty of Science University of Nottingham Malaysia Campus Jalan Broga 43500 Semenyih Selangor Malaysia
Correspondence should be addressed to Kang Nee Ting kang-neetingnottinghamedumy
Received 26 February 2014 Revised 4 June 2014 Accepted 18 June 2014 Published 30 June 2014
Academic Editor Ranadhir Chakraborty
Copyright copy 2014 Carolina Santiago et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
The inhibitory activity of a semipure fraction from the plantAcalypha wilkesiana assigned as 9EA-FC-B alone and in combinationwith ampicillin was studied against methicillin-resistant Staphylococcus aureus (MRSA) In addition effects of the combinationtreatment onPBP2a expressionwere investigatedMicrodilution assaywas used to determine theminimal inhibitory concentrations(MIC) Synergistic effects of 9EA-FC-B with ampicillin were determined using the fractional inhibitory concentration (FIC) indexand kinetic growth curve assayWestern blot experiments were carried out to study the PBP2a expression in treatedMRSA culturesThe results showed a synergistic effect between ampicillin and 9EA-FC-B treatment with the lowest FIC index of 019 (synergismle 05) The presence of 9EA-FC-B reduced the MIC of ampicillin from 50 to 156 120583gmLminus1 When ampicillin and 9EA-FC-B werecombined at subinhibitory level the kinetic growth curves were suppressed The antibacterial effect of 9EA-FC-B and ampicillinwas shown to be synergistic The synergism is due the ability of 9EA-FC-B to suppress the activity of PBP2a thus restoring thesusceptibility of MRSA to ampicillin Corilagin was postulated to be the constituent responsible for the synergistic activity showedby 9EA-FC-B
1 Introduction
Infection caused by methicillin-resistance Staphylococcusaureus (MRSA) is a world-wide health problemThese infec-tions are predominantly observed among immunocompro-mised patients in hospitals However in recent years therehave been an increasing number of fatalities failed treatmentand healthcare costs Progressive escalation of antibioticresistance in MRSA has resulted in a limited option fortreatment [1]Thus there is an immediate need for alternativetherapies to control the spread of illness caused by MRSA
A key strategy in combating a resistant microorganism isto suppress its resistance factorThere are at least two mecha-nisms that Staphylococci can evade beta-lactam toxicitywhichare by synthesizing the penicillin-binding protein 2a (PBP2a)and 120573-lactamases [2] In normal circumstances Staphylococ-cus aureus strains produce penicillin-binding proteins (PBPs)for synthesis of bacterial cell wall However when exposedto lethal concentration of beta-lactams antibiotics resistantS aureus strains produce PBP2a which has unusually low
binding affinity to beta-lactams [3] PBP2a in turn replacesthe function of normal PBPs (blocked by beta-lactams) in theresistant strains for cell wall biosynthesis [4] This confersMRSA resistance to the entire beta-lactam family Hencesuppression of PBP2a production is a promising approachto overcome MRSArsquos resistance By doing so there is apossibility to restore the susceptibility of MRSA to beta-lactam antibiotics [5]
It has been observed that extracts fractions or metabo-lites of plant origin are able to inhibit production of PBP2awhen they are used in combination with current availableantibiotics In a review article [6] combinations of drugssuch as beta-lactams and beta-lactamase inhibitors of plantorigin have been shown to exhibit synergistic activitiesagainst antibiotic resistant microorganisms The article alsohighlights plants as a source of small molecule antibioticsand synergism observed in natural products with antibi-otics against growth of bacteria fungus and mycobacteriaUsing combination treatment as a strategy to tackle resistantmicroorganism has also demonstrated several successes at
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 965348 7 pageshttpdxdoiorg1011552014965348
2 BioMed Research International
experimental level For an example combination of corila-gin from the plant Arctostaphylos uva-ursi with oxacillinsuccessfully inhibited production of PBP2a in MRSA TheMIC of oxacillin and other tested beta-lactams was reducedbetween 100-fold and 2000-fold strongly suggesting thatcombination treatment can potentially be an alternativemethod to combat virulence of MRSA [7]
A wilkesiana is a medicinal plant which has widelybeen utilized for treating bacterial and fungal infections[8] In some cases the plant is also used to treat malariagastrointestinal problems [9] and potentially cancers [1011] Previously we found anti-MRSA and other antibacterialactivities in the ethyl acetate and ethanol extracts of Awilkesiana [12] In continuation to our earlier findings wehave now embarked to further investigate the effects of thesemipure extracts ofAwilkesiana in the reversal of ampicillinresistance in MRSA
2 Methods
21 Plant Extraction The plant material was collected fromBroga SelangorMalaysia (September 2010) Voucher sampleis deposited in the herbariumof Faculty of ScienceUniversityof Nottingham Malaysia Campus and assigned as UNMC9 The dried plant material (36 kg) consisting of the wholeplant was subjected to sequential extraction using n-hexanefollowed by ethyl acetate and finally 95 ethanol [13 14]
22 Isolation of Bioactive Fraction 9EA-FC-B The ethylacetate extract of A wilkesiana (9EA) was fractionatedby using vacuum liquid chromatography (silica gel) Thesolvent system used for elution was n-hexane (He) withincreasing amount of chloroform (CHCl
3
) and CHCl3
withincreasing amount of methanol (MeOH) [HeCHCl
3
(1 1)rarr CHCl
3
rarr CHCl3
MeOH (97 3 vv) rarr CHCl3
MeOH(95 5 vv) rarr CHCl
3
MeOH (93 7 vv) rarr CHCl3
MeOH(90 10 vv) rarr CHCl
3
MeOH (85 15 vv)] Fractionsobtainedwere further fractionated via preparative centrifugalthin layer chromatography (silica gel) using a similar solventsystem The semipurified fractions were then tested for anti-MRSA activity
23 Microorganism and Growth Conditions Methicillin sen-sitive S aureusATCC 11632 (MSSA) was grown in tryptic soybroth (TSB) (Hi-Media India) at 37∘C for 24 h with a shakingmode of 220 rpm Aliquot from this suspension was streakedon tryptic soy agar (TSA) (Hi-Media India) and incubatedat 37∘C for another 24 h Two to four single colonies fromthe TSA plate were inoculated in 10mL of Muller Hintonbroth (MHB) (Hi-Media India) and allowed to grow at 37∘Cuntil they reached exponential stage (2 times 108 CFUmLminus1)The suspension was then used for microbroth dilution assayMRSAATCC 43300 was grown similarly except all themediaused was supplemented with 2 sodium chloride (NaCl)(Merck Germany) and incubation temperature was at 35∘CBacterial stocks were kept at minus80∘C in TSB added with 10(vv) glycerol (Sigma USA)
24 Test Samples The crude ethyl acetate extract of Awilkesiana 9EA and a bioactive fraction derived from it 9EA-FC-B (identified from previous experiments) were dissolvedin dimethyl sulfoxide (DMSO) (Sigma USA) at stock con-centration of 100mgmLminus1 Further dilution was carried outusing media and the final concentration of DMSO in themedia did not exceed 1 Our earlier study has reported thelack of solvent (DMSO) effect in the test samples [14] Antibi-otics for susceptibility testing were prepared at 10mgmLminus1in sterile distilled water Tested antibiotics were ampicilin(Amresco USA) oxacillin (Discovery Fine Chemicals UK)and methicillin (Sigma USA)
25 Determination ofMIC MICs of antibiotics crude extract9EA and the active fraction 9EA-FC-B against MRSA andMSSA were determined via microdilution assay with a 96-well plate Test samples were tested in a twofold serialdilution Antibiotics were tested with concentrations rangingfrom 019 to 100 120583gmLminus1 and plant extract samples from 009to 12mgmLminus1
MSSA andMRSAbroth cultures were grown inMHB andMHB + 2NaCl respectively until an exponential stage (2 times108 CFUmLminus1) was reached (see Section 23) These brothcultures were diluted to correspond to final inoculums of5 times 105 CFUmLminus1 upon inoculation into eachwell containingtwofold serial dilutions of test sampleMedia used in the assaywere MHB for MSSA and MHB +2 NaCl for MRSA Inthe final incubation step plates were incubated for 24 h at37∘C forMSSA and at 35∘C forMRSA General guidelines forthis experiment were obtained from Clinical and LaboratoryStandards Institute 2007 [15] with recommendations adaptedfrom several other studies [16ndash18]
26 Synergistic Studies
261 Growth Curves Assay MRSA was grown in a 96-wellplate in the presence of following subinhibitory concentra-tions of 9EA-FC-B 14 times MIC (075mgmLminus1) 18 times MIC(038mgmLminus1) and 116 timesMIC (019mgmLminus1) in combina-tion with subinhibitory concentrations of ampicillin rangingfrom 12 times MIC to 164 times MIC (25 to 078 120583gmLminus1) underaerobic condition Cell growth was monitored by readingoptical density (OD) values at 600nmat indicated timepointsfor 24 h Reading was monitored by using Varioskan FlashMultimode Reader (Thermo Scientific USA)
262 FIC Index Interpretation FIC index for the com-bination treatments in synergy growth curves assay wascalculated The formula used was FIC ampicillin = MIC ofampicillin in combinationMIC ampicillin alone FIC plantextract = MIC of plant extract in combinationMIC of plantextract alone and FIC index = FIC ampicillin + FIC plantextract The combination was defined as synergy if the FICindex was le05 indifference was defined gt05 but le40 andantagonism was defined as when the FIC index was gt4 [19]
BioMed Research International 3
27 Protein Extraction MRSA was grown in MHB + 2NaCl in the presence of subinhibitory concentrations of 9EA-FC-B until late exponential phase The bacterial lysates wereprepared in an extraction buffer containing Tris and EDTAand culture supernatants were harvested via centrifugation4340 g at 4∘C for 10 min The collected pellets were thentreated with 150mgmLminus1 lysozyme DNase and proteininhibitors cocktail before being subjected to 2 h incubationat 37∘C To enhance cell disruption 15-minute sonication inice-bucket was done Following 15min of centrifugation at13 850 g the pellets were obtained as the insoluble proteinextracts that were harvested in elution buffer containingTris urea and sodium dihydrogen phosphate Protein con-centrations were measured using Pierce 660 nm proteinassay
28 SDS-PAGE and Western Blot Assay Extracted protein(3 120583gmLminus1) was stained with 4X lithium dodecyl sulphate(LDS) sample buffer and subjected to sodium dodecyl sul-fate (SDS)mdashpolyacrylamide (12) gel electrophoresis run at120V Upon completion the gel was stained in CoomassieBlue staining solution until a clear background was obtainedfor scanning with GS-800 calibrated densitometer (Bio-RadUSA) In western blot analyses electrophoresed gels weretransferred to BioTrace NT nitrocellulose transfer mem-brane (Pall USA) Membranes were incubated overnightat 4∘C in gelatin from cold water fish skin (blockingagent) (Sigma USA) The production of PBP2a fromMRSA was detected by probing the membranes with mouseanti-PBP2a primary antibody (Denka Seiken Japan) andantiglyceraldehyde 3-phosphate dehydrogenase (GAPDH)(Thermo Scientific USA) with a dilution factor of 1 10000The same membranes were hybridized with anti-mousehorseradish peroxidase-linked secondary antibody (AbcamUK) diluted to 1 10000 to facilitate colorimetric detectionwith 331015840551015840-tetramethylbenzidine (TMB) substrate (NacalaiTesque Japan) Assay response was recorded using GS-800 calibrated densitometer (Bio-Rad USA) Densitometricquantification of western blot images was done using ImageJ 138 programme (Windows version of NIH Image) Resultswere scored in percentage of expression () normalized toGAPDH control
3 Results
31 Anti-MRSA Activity of Antibiotics and A wilkesianaExtract The MIC values of ampicillin crude extract of Awilkesiana 9EA and fraction 9EA-FC-B for MRSA ATCC43300 and MSSA 11632 are presented in Table 1 The MICvalue of ampicillin against MRSA notably confirmed theresistance of the studied strain whileMIC value of ampicillinagainst MSSA revealed the susceptibility of this strain tothe antibiotic Crude extract 9EA demonstrated antibacterialactivity against both the resistant and sensitive strains testedwith a lower MIC value observed for MSSA The anti-MRSAactivity of fraction 9EA-FC-B was fourfold more effectivethan crude extract 9EA
Table 1 MIC values of ampicillin crude extract 9EA and fraction9EA-FC-B against MRSA and MSSA
StrainMIC
Ampicillin(120583gmLminus1)
9EA(mgmLminus1)
9EA-FC-B(mgmLminus1)
MRSA 50 12 3MSSA 625 6 3Values represent triplicates of three independent experiments
32 Synergistic Effects of 9EA-FC-B with Ampicillin on MRSAGrowth Curves The synergistic effects of 9EA-FC-B withampicillin at subinhibitory concentrations on the growth ofMRSA are shown in Figure 1 The growth inhibitory effectsshown by both ampicillin and 9EA-FC-B on MRSA areconcentration dependent while the curve for the untreatedMRSA culture (control) showed an exponential growthSuppression of MRSA growth was detected when the MRSAcultures were treatedwith 9EA-FC-B alone at 14timesMIC 18timesMICand 116timesMIC (not shown in graph)The growth curvessuggested enhanced growth inhibitory effects when MRSAwas treated with ampicillin at subinhibitory concentrationsin the presence of 9EA-FC-B also at subinhibitory concen-trations The inhibitory effect was observed at as low as 132times MIC ampicillin in the presence of 14 times MIC 9EA-FC-BSimilar inhibitory effect was also achieved in the presenceof 18 times MIC of 9EA-FC-B in combination with 116 times MICampicillin
33 Synergistic Effects of 9EA-FC-B with Ampicillin Based onFIC Index The FIC indices for the tested combinations arepresented in Table 2 Synergistic effects were observed when9EA-FC-B was introduced in the treatment at 14 times MIC18 times MIC and 116 times MIC with ampicillin at subinhibitoryconcentrations (14 to 132 times MIC) In the presence of 14times MIC 9EA-FC-B synergistic effects were observed for thewidest range of subinhibitory concentrations of ampicillinand the range gradually decreased as the subinhibitoryconcentration of 9EA-FC-B was lowered
Analysis of the FIC indices revealed the new MIC valuesof ampicillin in the presence of 9EA-FC-B at subinhibitorylevel (Table 3) 9EA-FC-B at subinhibitory concentrations hasenhanced the activity of ampicillin by up to 32-fold againstMRSA for example MIC of ampicillin alone is 50 120583gmLminus1while the MIC of ampicillin in the presence of 9EA-FC-B at14 timesMIC is 156 120583gmLminus1 It is notable that theMIC values ofampicillin (forMRSA) in the presence of 14 timesMIC 9EA-FC-B and 18 timesMIC 9EA-FC-B are lower than the MIC value ofampicillin for MSSA that is 625 120583gmLminus1
34 Expression of PBP2a Expression of PBP2a was detectedat 76 kDaThe percentage of PBP2a expression in tested treat-ments is summarized in Figure 2 GAPDH which served asan internal control was detected in all treatments (results notshown)The presence of PBP2a band was detected for MRSAcultures that were grown at subinhibitory concentrations ofampicillin (116 times MIC and 132 times MIC) in western blot
4 BioMed Research International
0
01
02
03
04
05
06
07
08
0 2 4 6 8 10 12 14 16 18 20 22 24
Opt
ical
den
sity
at600
nmIncubation time (h)
0
01
02
03
04
05
06
07
08
0 2 4 6 8 10 12 14 16 18 20 22 24
Opt
ical
den
sity
at600
nm
Incubation time (h)
Control MRSA132 times MIC AMP
Control MRSA116 times MIC AMP
09
14 times MIC 9EA-FC-B
18 times MIC 9EA-FC-B
132 times MIC AMP + 14 times MIC 9EA-FC-B
116 times MIC AMP + 18 times MIC 9EA-FC-B
Figure 1 Effects of subinhibitory concentration of ampicillin alone 9EA-FC-B alone and combination treatment on the growth of MRSACell growth wasmeasured by usingOD at 600 nm at indicated time pointsThe curves represent triplicates of three independent experimentsError bars show the standard deviation (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 2 FIC indices of some combinations of ampicillin and 9EA-FC-B for MRSA
Ampicillin (120583gmLminus1) 9EA-FC-B (mgmLminus1)14 timesMIC (075) 18 timesMIC (038) 116 timesMIC (019)
12 timesMIC (25) 075 065 05614 timesMIC (125) 043 036 03118 timesMIC (625) 038 025 mdash116 timesMIC (3125) 031 019 mdash132 timesMIC (1563) 028 mdash mdash164 timesMIC (0781) mdash mdash mdashValues represent triplicates of three independent experiments Index interpretation le05 = synergy gt05 but le40 = indifference and gt4 = antagonism (MIC =minimum inhibitory concentrations mdash = no activity)
experiment Both western blot and quantitative densitomet-ric analysis showed that these cultures have higher expressionof PBP2a compared to the untreatedMRSA culture (control)Inhibition of PBP2a expression was observed when MRSAcultures were exposed to subinhibitory concentrations of9EA-FC-B where no PBP2a bandwas seen at 14 timesMIC 9EA-FC-B and only a very low expression (39) was detected at18 times MIC 9EA-FC-B Likewise the MRSA culture did not
show the presence of PBP2a when grown in the presence of132 timesMIC ampicillin + 14 timesMIC 9EA-FC-B
4 Discussion
The scope of the study is to explore the potential of usingactive plant extracts or fractions to combat resistance in
BioMed Research International 5
L 1 2 3 4 5 6 7
76kDa PBP2a80kDa
(a)
MRS
A
MSS
A
116times
MIC
AM
P
132times
MIC
AM
P
14times
MIC
9EA
-FC-
B
18times
MIC
9EA
-FC-
B
0
10
20
30
40
50
60
70
80
90
100
Expr
essio
n of
PBP
2a (
)
14
MIC
9EA
-F-C
-B132times
MIC
AM
P+
(b)
Figure 2 Expression of penicillin-binding protein 2a (PBP2a) of MRSA cultures grown in the presence of subinhibitory concentrations of9EA-FC-B and ampicillin (a)Western blot image lane L molecular mass markers 1 control MRSA 2 control MSSA 3 116 timesMICAMP 4132 timesMICAMP 5 14timesMIC 9EA-FC-B 6 18timesMIC 9EA-FC-B 7 132 timesMICAMP+ 14timesMIC 9EA-FC-B (b) Quantitative densitometricanalysis of PBP2a expression ofMRSA cultures grown in the presence of ampicillin alone 9EA-FC-B alone and in combinations normalizedto GAPDH loading control (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 3 MIC values of ampicillin in combination with subin-hibitory concentrations of 9EA-FC-B for MRSA
Treatment MIC (120583gmLminus1) of ampicillinAmpicillin alone 50With 9EA-FC-B at 14 timesMIC 156With 9EA-FC-B at 18 timesMIC 313With 9EA-FC-B at 116 timesMIC 125Values represent triplicates of three independent experiments (MIC =minimum inhibitory concentrations)
MRSA For the first time in 2006 US Food and DrugAdministration (FDA 2006) has approved a special green teaextract containing a proprietary mixture of phytochemicals(the active ingredient listed as Polyphenon E) [20] as aprescription drug for the topical (external) treatment ofgenital warts caused by the human papilloma virus (HPV)This example reinforces the efforts to study medicinal plantextracts for possible application in clinical practice Thepresent result established the antimicrobial activity of frac-tion 9EA-FC-B obtained from the ethyl acetate crude extractof A wilkesiana It also demonstrated synergism betweenthe fraction 9EA-FC-B and ampicillin in overcoming resis-tance of MRSA by inhibiting production of PBP2a In thepresence of subinhibitory concentrations of 9EA-FC-B theMIC of ampicillin was reduced by as much as 32-fold from50120583gmLminus1 to 156 120583gmLminus1 indicating that MRSA becamemore sensitive to ampicillin when fraction 9EA-FC-B was
introduced in the treatment Based on these findings wepredict that the active constituents from fraction 9EA-FC-B may potentially be used for combating MRSArsquos virulenceSeveral researches have exploited the synergistic effects ofnatural products for drug development [21ndash23] The mostapt example that is closely related to the present study isthe synergistic effects of corilagin and tellimagrandin I incombination with beta-lactam antibiotics on antibacterialactivity against MRSA by inactivation of PBP2a [24]
Results of bacterial growth curve experiment indicatedthat combination of 9EA-FC-B with ampicillin (both agentsat subinhibitory concentrations) distinctly suppressed thegrowth of MRSA in contrast to MRSA cultures that weretreated with either 9EA-FC-B or ampicillin alone GenerallyMIC of ampicillin reduced when subinhibitory concentra-tion of 9EA-FC-B increased in the combination treatmentFrom the kinetic growth curves we were able to deducethat the antimicrobial action of the ampicillin and 9EA-FC-B combination was observed at the beginning of theexponential phase A very minimal bacterial growth wasseen with increase in incubation hours Instead of growingrapidly during the exponential stage the graph portrayedlow growth of bacterial cells (approximately fourfold lower)with extended lag phase compared to the untreated MRSA(control) An extended lag phase was also detected forMRSAtreatedwith 9EA-FC-B alone Lag phase is the particular stagewhen bacteria equilibrate to adapt to the new environment byundergoing macromolecular repair and synthesis of cellulargrowth through DNA replications [25] Hence we deduced
6 BioMed Research International
that a lengthy lag phase observed in our experiment isdue to the inhibition of DNA replications that delays thecellular growth process The prolonged lag phase of MRSAobserved in this experiment is reminiscent of the action offluoroquinolones that caused inhibition of DNA replicationinMRSA leading to a longer lag phase [26] A potent antimi-crobial action was also identified at the exponential phase inwhich bacterial cells were prevented from growing rapidlyin the presence of 9EA-FC-B alone and in combination withampicillin This phenomenon showed probable interferencein cell division which involves multiple rounds of DNAsynthesis that are controlled by a variety of gene regulatoryprocesses [27 28] The plausible mechanism of action of9EA-FC-B observed at the exponential phase is thereforeassociated with the interruption of cell division that causesmembrane derangements and failure inmembrane functions
As for interpretation of the FIC indices lower indicesindicate better synergism [19] Based on the FIC indicesobtained eight out of the 18 combinations tested showedsynergistic effects The presence of 9EA-FC-B significantlyenhanced the potency of ampicillin by up to 32-fold (MICreduced from 50 120583gmLminus1 in the absence of 9EA-FC-B to156 120583gmLminus1 in the presence of 14 timesMIC of 9EA-FC-B) forMRSA
Ampicillin is a beta-lactam antibiotic that is designedto inhibit PBPs involved in late stage of peptidoglycanbiosynthesis Interference with peptidoglycan biosynthesiscauses deformities in the bacterial cell wall and eventuallyleads to cell death due to high internal osmotic pres-sure Nearly all bacteria can be inhibited by interfering inmechanism of peptidoglycan synthesis [29] Neverthelesstargeting this mechanism is no longer effective due to theproduction PBP2a in MRSA The blocking of normal PBPsby beta-lactams did not exert effects on peptidoglycan orcell wall synthesis because PBP2a replaces their functionand ensures normal formation of cell wall in presence oflethal concentration of beta-lactam drugs [30] However inthis study we experimentally demonstrated restoration ofampicillinrsquos antimicrobial activity by the addition of 9EA-FC-B at subinhibitory level
The synergistic activity observed between 9EA-FC-Band ampicillin against MRSA was shown to be associatedwith inhibition of PBP2a PBP2a is an inducible proteinthat regulates methicillin resistance Its expression is het-erogeneous in nature amidst level of resistance differing tothe beta-lactam being used [31] The mecA gene complexwhich encodes for this protein encompasses the regulatorygenes mecI and mecR Interaction of beta-lactam antibioticswith these regulatory genes eventually allows expression ofmecA in terms of production of PBP2a [32ndash34] As suchthe occurrence of intense PBP2a bands in MRSA culturesgrown in low concentrations of ampicillin compared to thecontrol culture in western blot experiment suggested theinduction of mecA gene transcription In contrast no PBP2abands were detected in MRSA cultures that were exposed to14 times MIC of 9EA-FB-C while PBP2a is only very mildlyexpressed when the concentration of 9EA-FC-B was loweredto 18 timesMIC This suggests that 9EA-FC-B can either inhibitthe production of PBP2a or directly inactivate PBP2a This
suggestion is in agreement with the observation that 9EA-FC-B enhanced the ampicillin susceptibility of MRSA In2004 Shiota et al [24] reported the antibacterial effect ofcorilagin and tellimagrandin I (isolated from Arctostaphylosuva-ursi and Rosa canina resp) against MRSA The twopolyphenolic compounds had exceptionally reduced MICsof beta-lactams for MRSA via inactivation of PBP2a Webelieve that corilagin (a tannin) is also present in the testfraction 9EA-FC-B since corilagin was previously isolatedand identified by our colleagues from the same source of plantmaterials [35] This is further supported by phytochemicaltesting that showed the presence of tannins in 9EA-FC-B(data not shown) Although Shimizu et al [7] previouslyreported that corilagin enhanced antibacterial activity of var-ious beta-lactams by 100- to 2000-fold against MRSA 9EA-FC-B was only shown to enhance the activity of ampicillinby up to 32-fold in the present study The stark differencein antibacterial enhancements can readily be rationalized bythe fact that 9EA-FC-B is a semipure fraction that possiblycontains only a small amount of corilagin in addition tomany other secondarymetabolites that were assumed to havenegligible effect on the antibacterial activity observed
5 Conclusion
From the results reported in this study we found that theantibacterial effect of the fraction 9EA-FC-B obtained fromA wilkesiana and ampicillin is synergistic The synergism isdue the ability of 9EA-FC-B to suppress the production ofPBP2a or directly inactivate it leading to the restoration ofthe susceptibility of MRSA to ampicillin
Conflict of Interests
The authors declared no conflict of interests
Acknowledgment
This study was supported by Science Fund scheme underMinistry of Agriculture and Agro-Based Industry Malaysia(MOA) Malaysia with Grant no M00485401
References
[1] J Snider M Rapids and B Rivard ldquoMethicillin-resistantStaphylococcus aureus a review of current antibiotic therapyrdquoSpectrum Health Grand Rapids 2012
[2] M T Gillespie J W May and R A Skurray ldquoAntibiotic resis-tance in Staphylococcus aureus isolated at an Australian hospitalbetween 1946 and 1981rdquo Journal of Medical Microbiology vol 19no 2 pp 137ndash147 1985
[3] B Berger-Bachi and S Rohrer ldquoFactors influencing methicillinresistance in staphylococcirdquo Archives of Microbiology vol 178no 3 pp 165ndash171 2002
[4] B Berger-Bachi ldquoExpression of resistance to methicillinrdquoTrends in Microbiology vol 2 no 10 pp 389ndash393 1994
[5] J Vouillamoz J M Entenza P Hohl and P MoreillonldquoLB11058 a new cephalosporin with high penicillin-bindingprotein 2a affinity and activity in experimental endocarditis due
BioMed Research International 7
to homogeneously methicillin-resistant Staphylococcus aureusrdquoAntimicrobial Agents and Chemotherapy vol 48 no 11 pp4322ndash4327 2004
[6] S Hemaiswarya A K Kruthiventi and M Doble ldquoSynergismbetween natural products and antibiotics against infectiousdiseasesrdquo Phytomedicine vol 15 no 8 pp 639ndash652 2008
[7] M Shimizu S Shiota T Mizushima et al ldquoMarked poten-tiation of activity of 120573-lactams against methicillin-resistantStaphylococcus aureus by corilaginrdquo Antimicrobial Agents andChemotherapy vol 45 no 11 pp 3198ndash3201 2001
[8] P I Alade andO N Irobi ldquoAntimicrobial activities of crude leafextracts ofAcalypha wilkesianardquo Journal of Ethnopharmacologyvol 39 no 3 pp 171ndash174 1993
[9] B E Akinde andOO Odeyemi ldquoExtraction andmicrobiologi-cal evaluation of the oils from the leaves ofAcalyphawilkesianardquoNigerian Medical Journal vol 17 pp 163ndash165 1987
[10] S W Lim K N Ting T D Bradshaw et al ldquoAcalyphawilkesiana extracts induce apoptosis by causing single strandand double strand DNA breaksrdquo Journal of Ethnopharmacologyvol 138 no 2 pp 616ndash623 2011
[11] S W Lim H S Loh K N Ting T D Bradshaw and N AZeenathul ldquoAcalypha wilkesiana ethyl acetate extract enhancesthe in vitro cytotoxic effects of 120572-tocopherol in human brainand lung cancer cellsrdquo International Journal of BioscienceBiochemistry and Bioinformatics vol 3 no 4 pp 335ndash340 2013
[12] M Othman S Genapathy P S Liew et al ldquoSearch for antibac-terial agents from Malaysian rainforest and tropical plantsrdquoNatural Product Research vol 25 no 19 pp 1857ndash1864 2011
[13] G Spigno and D M de Faveri ldquoAntioxidants from grape stalksandmarc influence of extraction procedure on yield purity andantioxidant power of the extractsrdquo Journal of Food Engineeringvol 78 no 3 pp 793ndash801 2007
[14] MOthmanH S Loh CWiart T J Khoo KH Lim andKNTing ldquoOptimal methods for evaluating antimicrobial activitiesfrom plant extractsrdquo Journal of Microbiological Methods vol 84no 2 pp 161ndash166 2011
[15] Clinical and Laboratory Standards Institute (CLSI) ldquoLabora-tory methods for detection of methicillin-resistant Staphylococ-cus aureus (MRSA)rdquo Supplement M100-S17 2007
[16] JMAndrews ldquoDetermination ofminimum inhibitory concen-trationsrdquo Journal of Antimicrobial Chemotherapy vol 48 no 1pp 5ndash16 2001
[17] D F J Brown ldquoDetection of methicillinoxacillin resistance instaphylococcirdquo Journal of Antimicrobial Chemotherapy vol 48no 1 pp 65ndash70 2001
[18] P Cos A J Vlietinck D V Berghe and LMaes ldquoAnti-infectivepotential of natural products how to develop a stronger in vitroproof-of-conceptrdquo Journal of Ethnopharmacology vol 106 no3 pp 290ndash302 2006
[19] R L White D S Burgess M Manduru and J A BossoldquoComparison of three different in vitro methods of detectingsynergy time-kill checkerboard and E testrdquo AntimicrobialAgents and Chemotherapy vol 40 no 8 pp 1914ndash1918 1996
[20] US Food andDrug Administration (FDA)CDERNewMolecu-lar Entity (NME) Drug and New Biologic Approvals in CalendarYear 2006
[21] F J Schmitz A C Fluit M Luckefahr et al ldquoThe effect ofreserpine an inhibitor of multidrug efflux pumps on the in-vitro activities of ciprofloxacin sparfloxacin and moxifloxacinagainst clinical isolates of Staphylococcus aureusrdquo Journal ofAntimicrobial Chemotherapy vol 42 no 6 pp 807ndash810 1998
[22] F R Stermitz P Lorenz J N Tawara L A Zenewicz and KLewis ldquoSynergy in a medicinal plant Antimicrobial action ofberberine potentiated by 5rsquo-methoxyhydnocarpin a multidrugpump inhibitorrdquoProceedings of theNational Academy of Sciencesof theUnited States of America vol 97 no 4 pp 1433ndash1437 2000
[23] Y Cho N L Schiller and K Oh ldquoAntibacterial effects of greentea polyphenols on clinical isolates of methicillin-resistantStaphylococcus aureusrdquo Current Microbiology vol 57 no 6 pp542ndash546 2008
[24] S Shiota M Shimizu J Sugiyama Y Morita T Mizushimaand T Tsuchiya ldquoMechanisms of action of corilagin andtellimagrandin I that remarkably potentiate the activity of120573-lactams against methicillin-resistant Staphylococcus aureusrdquoMicrobiology and Immunology vol 48 no 1 pp 67ndash73 2004
[25] S Dukan and T Nystrom ldquoBacterial senescence stasis resultsin increased and differential oxidation of cytoplasmic proteinsleading to developmental induction of the heat shock regulonrdquoGenes amp Development vol 12 no 21 pp 3431ndash3441 1998
[26] R A Venezia B E Domaracki A M Evans K E Preston andE M Graffunder ldquoSelection of high-level oxacillin resistancein heteroresistant Staphylococcus aureus by fluoroquinoloneexposurerdquo Journal of Antimicrobial Chemotherapy vol 48 no3 pp 375ndash381 2001
[27] J M Navarro Llorens A Tormo and E Martınez-Garcıa ldquoSta-tionary phase in gram-negative bacteriardquo FEMS MicrobiologyReviews vol 34 no 4 pp 476ndash495 2010
[28] M D Rolfe C J Rice S Lucchini et al ldquoLag phase is a distinctgrowth phase that prepares bacteria for exponential growth andinvolves transient metal accumulationrdquo Journal of Bacteriologyvol 194 no 3 pp 686ndash701 2011
[29] P Lambert ldquoMechanism of action of antibiotics and synthethicanti-infective agentsrdquo in Hugo and Russells PharmaceuticalMicrobiology S PDenyerNAHodges and S PGorman Edspp 202ndash206 Blackwell Publishing Oxford UK 7th edition2004
[30] A Smith ldquoBacterial resistance to antibioticsrdquo inHugo and Rus-sellrsquos Pharmaceutical Microbiology S P Denyer N A Hodgesand S P Gorman Eds pp 220ndash223 Blackwell Oxford UK7th edition 2004
[31] H F Chambers ldquoMethicillin resistance in staphylococcimolec-ular and biochemical basis and clinical implicationsrdquo ClinicalMicrobiology Reviews vol 10 no 4 pp 781ndash791 1997
[32] G L Archer and JM Bosilevac ldquoSignaling antibiotic resistancein staphylococcirdquo Science vol 291 no 5510 pp 1915ndash1916 2001
[33] H Z Zhang C J Hackbarth K M Chansky and H FChambers ldquoA proteolytic transmembrane signaling pathwayand resistance to 120573-lactams in Staphylococcirdquo Science vol 291no 5510 pp 1962ndash1965 2001
[34] S Deresinski ldquoMethicillin-resistant Staphylococcus aureus anevolutionary epidemiologic and therapeutic odysseyrdquo ClinicalInfectious Diseases vol 40 no 4 pp 562ndash573 2005
[35] W M Din J Chu G Clarke et al ldquoAntioxidant and cytopro-tective effects of an ethanol extract of Acalypha wilkesiana varmacafeana from Malaysiardquo Natural Product Communicationsvol 8 no 3 pp 375ndash380 2013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
experimental level For an example combination of corila-gin from the plant Arctostaphylos uva-ursi with oxacillinsuccessfully inhibited production of PBP2a in MRSA TheMIC of oxacillin and other tested beta-lactams was reducedbetween 100-fold and 2000-fold strongly suggesting thatcombination treatment can potentially be an alternativemethod to combat virulence of MRSA [7]
A wilkesiana is a medicinal plant which has widelybeen utilized for treating bacterial and fungal infections[8] In some cases the plant is also used to treat malariagastrointestinal problems [9] and potentially cancers [1011] Previously we found anti-MRSA and other antibacterialactivities in the ethyl acetate and ethanol extracts of Awilkesiana [12] In continuation to our earlier findings wehave now embarked to further investigate the effects of thesemipure extracts ofAwilkesiana in the reversal of ampicillinresistance in MRSA
2 Methods
21 Plant Extraction The plant material was collected fromBroga SelangorMalaysia (September 2010) Voucher sampleis deposited in the herbariumof Faculty of ScienceUniversityof Nottingham Malaysia Campus and assigned as UNMC9 The dried plant material (36 kg) consisting of the wholeplant was subjected to sequential extraction using n-hexanefollowed by ethyl acetate and finally 95 ethanol [13 14]
22 Isolation of Bioactive Fraction 9EA-FC-B The ethylacetate extract of A wilkesiana (9EA) was fractionatedby using vacuum liquid chromatography (silica gel) Thesolvent system used for elution was n-hexane (He) withincreasing amount of chloroform (CHCl
3
) and CHCl3
withincreasing amount of methanol (MeOH) [HeCHCl
3
(1 1)rarr CHCl
3
rarr CHCl3
MeOH (97 3 vv) rarr CHCl3
MeOH(95 5 vv) rarr CHCl
3
MeOH (93 7 vv) rarr CHCl3
MeOH(90 10 vv) rarr CHCl
3
MeOH (85 15 vv)] Fractionsobtainedwere further fractionated via preparative centrifugalthin layer chromatography (silica gel) using a similar solventsystem The semipurified fractions were then tested for anti-MRSA activity
23 Microorganism and Growth Conditions Methicillin sen-sitive S aureusATCC 11632 (MSSA) was grown in tryptic soybroth (TSB) (Hi-Media India) at 37∘C for 24 h with a shakingmode of 220 rpm Aliquot from this suspension was streakedon tryptic soy agar (TSA) (Hi-Media India) and incubatedat 37∘C for another 24 h Two to four single colonies fromthe TSA plate were inoculated in 10mL of Muller Hintonbroth (MHB) (Hi-Media India) and allowed to grow at 37∘Cuntil they reached exponential stage (2 times 108 CFUmLminus1)The suspension was then used for microbroth dilution assayMRSAATCC 43300 was grown similarly except all themediaused was supplemented with 2 sodium chloride (NaCl)(Merck Germany) and incubation temperature was at 35∘CBacterial stocks were kept at minus80∘C in TSB added with 10(vv) glycerol (Sigma USA)
24 Test Samples The crude ethyl acetate extract of Awilkesiana 9EA and a bioactive fraction derived from it 9EA-FC-B (identified from previous experiments) were dissolvedin dimethyl sulfoxide (DMSO) (Sigma USA) at stock con-centration of 100mgmLminus1 Further dilution was carried outusing media and the final concentration of DMSO in themedia did not exceed 1 Our earlier study has reported thelack of solvent (DMSO) effect in the test samples [14] Antibi-otics for susceptibility testing were prepared at 10mgmLminus1in sterile distilled water Tested antibiotics were ampicilin(Amresco USA) oxacillin (Discovery Fine Chemicals UK)and methicillin (Sigma USA)
25 Determination ofMIC MICs of antibiotics crude extract9EA and the active fraction 9EA-FC-B against MRSA andMSSA were determined via microdilution assay with a 96-well plate Test samples were tested in a twofold serialdilution Antibiotics were tested with concentrations rangingfrom 019 to 100 120583gmLminus1 and plant extract samples from 009to 12mgmLminus1
MSSA andMRSAbroth cultures were grown inMHB andMHB + 2NaCl respectively until an exponential stage (2 times108 CFUmLminus1) was reached (see Section 23) These brothcultures were diluted to correspond to final inoculums of5 times 105 CFUmLminus1 upon inoculation into eachwell containingtwofold serial dilutions of test sampleMedia used in the assaywere MHB for MSSA and MHB +2 NaCl for MRSA Inthe final incubation step plates were incubated for 24 h at37∘C forMSSA and at 35∘C forMRSA General guidelines forthis experiment were obtained from Clinical and LaboratoryStandards Institute 2007 [15] with recommendations adaptedfrom several other studies [16ndash18]
26 Synergistic Studies
261 Growth Curves Assay MRSA was grown in a 96-wellplate in the presence of following subinhibitory concentra-tions of 9EA-FC-B 14 times MIC (075mgmLminus1) 18 times MIC(038mgmLminus1) and 116 timesMIC (019mgmLminus1) in combina-tion with subinhibitory concentrations of ampicillin rangingfrom 12 times MIC to 164 times MIC (25 to 078 120583gmLminus1) underaerobic condition Cell growth was monitored by readingoptical density (OD) values at 600nmat indicated timepointsfor 24 h Reading was monitored by using Varioskan FlashMultimode Reader (Thermo Scientific USA)
262 FIC Index Interpretation FIC index for the com-bination treatments in synergy growth curves assay wascalculated The formula used was FIC ampicillin = MIC ofampicillin in combinationMIC ampicillin alone FIC plantextract = MIC of plant extract in combinationMIC of plantextract alone and FIC index = FIC ampicillin + FIC plantextract The combination was defined as synergy if the FICindex was le05 indifference was defined gt05 but le40 andantagonism was defined as when the FIC index was gt4 [19]
BioMed Research International 3
27 Protein Extraction MRSA was grown in MHB + 2NaCl in the presence of subinhibitory concentrations of 9EA-FC-B until late exponential phase The bacterial lysates wereprepared in an extraction buffer containing Tris and EDTAand culture supernatants were harvested via centrifugation4340 g at 4∘C for 10 min The collected pellets were thentreated with 150mgmLminus1 lysozyme DNase and proteininhibitors cocktail before being subjected to 2 h incubationat 37∘C To enhance cell disruption 15-minute sonication inice-bucket was done Following 15min of centrifugation at13 850 g the pellets were obtained as the insoluble proteinextracts that were harvested in elution buffer containingTris urea and sodium dihydrogen phosphate Protein con-centrations were measured using Pierce 660 nm proteinassay
28 SDS-PAGE and Western Blot Assay Extracted protein(3 120583gmLminus1) was stained with 4X lithium dodecyl sulphate(LDS) sample buffer and subjected to sodium dodecyl sul-fate (SDS)mdashpolyacrylamide (12) gel electrophoresis run at120V Upon completion the gel was stained in CoomassieBlue staining solution until a clear background was obtainedfor scanning with GS-800 calibrated densitometer (Bio-RadUSA) In western blot analyses electrophoresed gels weretransferred to BioTrace NT nitrocellulose transfer mem-brane (Pall USA) Membranes were incubated overnightat 4∘C in gelatin from cold water fish skin (blockingagent) (Sigma USA) The production of PBP2a fromMRSA was detected by probing the membranes with mouseanti-PBP2a primary antibody (Denka Seiken Japan) andantiglyceraldehyde 3-phosphate dehydrogenase (GAPDH)(Thermo Scientific USA) with a dilution factor of 1 10000The same membranes were hybridized with anti-mousehorseradish peroxidase-linked secondary antibody (AbcamUK) diluted to 1 10000 to facilitate colorimetric detectionwith 331015840551015840-tetramethylbenzidine (TMB) substrate (NacalaiTesque Japan) Assay response was recorded using GS-800 calibrated densitometer (Bio-Rad USA) Densitometricquantification of western blot images was done using ImageJ 138 programme (Windows version of NIH Image) Resultswere scored in percentage of expression () normalized toGAPDH control
3 Results
31 Anti-MRSA Activity of Antibiotics and A wilkesianaExtract The MIC values of ampicillin crude extract of Awilkesiana 9EA and fraction 9EA-FC-B for MRSA ATCC43300 and MSSA 11632 are presented in Table 1 The MICvalue of ampicillin against MRSA notably confirmed theresistance of the studied strain whileMIC value of ampicillinagainst MSSA revealed the susceptibility of this strain tothe antibiotic Crude extract 9EA demonstrated antibacterialactivity against both the resistant and sensitive strains testedwith a lower MIC value observed for MSSA The anti-MRSAactivity of fraction 9EA-FC-B was fourfold more effectivethan crude extract 9EA
Table 1 MIC values of ampicillin crude extract 9EA and fraction9EA-FC-B against MRSA and MSSA
StrainMIC
Ampicillin(120583gmLminus1)
9EA(mgmLminus1)
9EA-FC-B(mgmLminus1)
MRSA 50 12 3MSSA 625 6 3Values represent triplicates of three independent experiments
32 Synergistic Effects of 9EA-FC-B with Ampicillin on MRSAGrowth Curves The synergistic effects of 9EA-FC-B withampicillin at subinhibitory concentrations on the growth ofMRSA are shown in Figure 1 The growth inhibitory effectsshown by both ampicillin and 9EA-FC-B on MRSA areconcentration dependent while the curve for the untreatedMRSA culture (control) showed an exponential growthSuppression of MRSA growth was detected when the MRSAcultures were treatedwith 9EA-FC-B alone at 14timesMIC 18timesMICand 116timesMIC (not shown in graph)The growth curvessuggested enhanced growth inhibitory effects when MRSAwas treated with ampicillin at subinhibitory concentrationsin the presence of 9EA-FC-B also at subinhibitory concen-trations The inhibitory effect was observed at as low as 132times MIC ampicillin in the presence of 14 times MIC 9EA-FC-BSimilar inhibitory effect was also achieved in the presenceof 18 times MIC of 9EA-FC-B in combination with 116 times MICampicillin
33 Synergistic Effects of 9EA-FC-B with Ampicillin Based onFIC Index The FIC indices for the tested combinations arepresented in Table 2 Synergistic effects were observed when9EA-FC-B was introduced in the treatment at 14 times MIC18 times MIC and 116 times MIC with ampicillin at subinhibitoryconcentrations (14 to 132 times MIC) In the presence of 14times MIC 9EA-FC-B synergistic effects were observed for thewidest range of subinhibitory concentrations of ampicillinand the range gradually decreased as the subinhibitoryconcentration of 9EA-FC-B was lowered
Analysis of the FIC indices revealed the new MIC valuesof ampicillin in the presence of 9EA-FC-B at subinhibitorylevel (Table 3) 9EA-FC-B at subinhibitory concentrations hasenhanced the activity of ampicillin by up to 32-fold againstMRSA for example MIC of ampicillin alone is 50 120583gmLminus1while the MIC of ampicillin in the presence of 9EA-FC-B at14 timesMIC is 156 120583gmLminus1 It is notable that theMIC values ofampicillin (forMRSA) in the presence of 14 timesMIC 9EA-FC-B and 18 timesMIC 9EA-FC-B are lower than the MIC value ofampicillin for MSSA that is 625 120583gmLminus1
34 Expression of PBP2a Expression of PBP2a was detectedat 76 kDaThe percentage of PBP2a expression in tested treat-ments is summarized in Figure 2 GAPDH which served asan internal control was detected in all treatments (results notshown)The presence of PBP2a band was detected for MRSAcultures that were grown at subinhibitory concentrations ofampicillin (116 times MIC and 132 times MIC) in western blot
4 BioMed Research International
0
01
02
03
04
05
06
07
08
0 2 4 6 8 10 12 14 16 18 20 22 24
Opt
ical
den
sity
at600
nmIncubation time (h)
0
01
02
03
04
05
06
07
08
0 2 4 6 8 10 12 14 16 18 20 22 24
Opt
ical
den
sity
at600
nm
Incubation time (h)
Control MRSA132 times MIC AMP
Control MRSA116 times MIC AMP
09
14 times MIC 9EA-FC-B
18 times MIC 9EA-FC-B
132 times MIC AMP + 14 times MIC 9EA-FC-B
116 times MIC AMP + 18 times MIC 9EA-FC-B
Figure 1 Effects of subinhibitory concentration of ampicillin alone 9EA-FC-B alone and combination treatment on the growth of MRSACell growth wasmeasured by usingOD at 600 nm at indicated time pointsThe curves represent triplicates of three independent experimentsError bars show the standard deviation (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 2 FIC indices of some combinations of ampicillin and 9EA-FC-B for MRSA
Ampicillin (120583gmLminus1) 9EA-FC-B (mgmLminus1)14 timesMIC (075) 18 timesMIC (038) 116 timesMIC (019)
12 timesMIC (25) 075 065 05614 timesMIC (125) 043 036 03118 timesMIC (625) 038 025 mdash116 timesMIC (3125) 031 019 mdash132 timesMIC (1563) 028 mdash mdash164 timesMIC (0781) mdash mdash mdashValues represent triplicates of three independent experiments Index interpretation le05 = synergy gt05 but le40 = indifference and gt4 = antagonism (MIC =minimum inhibitory concentrations mdash = no activity)
experiment Both western blot and quantitative densitomet-ric analysis showed that these cultures have higher expressionof PBP2a compared to the untreatedMRSA culture (control)Inhibition of PBP2a expression was observed when MRSAcultures were exposed to subinhibitory concentrations of9EA-FC-B where no PBP2a bandwas seen at 14 timesMIC 9EA-FC-B and only a very low expression (39) was detected at18 times MIC 9EA-FC-B Likewise the MRSA culture did not
show the presence of PBP2a when grown in the presence of132 timesMIC ampicillin + 14 timesMIC 9EA-FC-B
4 Discussion
The scope of the study is to explore the potential of usingactive plant extracts or fractions to combat resistance in
BioMed Research International 5
L 1 2 3 4 5 6 7
76kDa PBP2a80kDa
(a)
MRS
A
MSS
A
116times
MIC
AM
P
132times
MIC
AM
P
14times
MIC
9EA
-FC-
B
18times
MIC
9EA
-FC-
B
0
10
20
30
40
50
60
70
80
90
100
Expr
essio
n of
PBP
2a (
)
14
MIC
9EA
-F-C
-B132times
MIC
AM
P+
(b)
Figure 2 Expression of penicillin-binding protein 2a (PBP2a) of MRSA cultures grown in the presence of subinhibitory concentrations of9EA-FC-B and ampicillin (a)Western blot image lane L molecular mass markers 1 control MRSA 2 control MSSA 3 116 timesMICAMP 4132 timesMICAMP 5 14timesMIC 9EA-FC-B 6 18timesMIC 9EA-FC-B 7 132 timesMICAMP+ 14timesMIC 9EA-FC-B (b) Quantitative densitometricanalysis of PBP2a expression ofMRSA cultures grown in the presence of ampicillin alone 9EA-FC-B alone and in combinations normalizedto GAPDH loading control (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 3 MIC values of ampicillin in combination with subin-hibitory concentrations of 9EA-FC-B for MRSA
Treatment MIC (120583gmLminus1) of ampicillinAmpicillin alone 50With 9EA-FC-B at 14 timesMIC 156With 9EA-FC-B at 18 timesMIC 313With 9EA-FC-B at 116 timesMIC 125Values represent triplicates of three independent experiments (MIC =minimum inhibitory concentrations)
MRSA For the first time in 2006 US Food and DrugAdministration (FDA 2006) has approved a special green teaextract containing a proprietary mixture of phytochemicals(the active ingredient listed as Polyphenon E) [20] as aprescription drug for the topical (external) treatment ofgenital warts caused by the human papilloma virus (HPV)This example reinforces the efforts to study medicinal plantextracts for possible application in clinical practice Thepresent result established the antimicrobial activity of frac-tion 9EA-FC-B obtained from the ethyl acetate crude extractof A wilkesiana It also demonstrated synergism betweenthe fraction 9EA-FC-B and ampicillin in overcoming resis-tance of MRSA by inhibiting production of PBP2a In thepresence of subinhibitory concentrations of 9EA-FC-B theMIC of ampicillin was reduced by as much as 32-fold from50120583gmLminus1 to 156 120583gmLminus1 indicating that MRSA becamemore sensitive to ampicillin when fraction 9EA-FC-B was
introduced in the treatment Based on these findings wepredict that the active constituents from fraction 9EA-FC-B may potentially be used for combating MRSArsquos virulenceSeveral researches have exploited the synergistic effects ofnatural products for drug development [21ndash23] The mostapt example that is closely related to the present study isthe synergistic effects of corilagin and tellimagrandin I incombination with beta-lactam antibiotics on antibacterialactivity against MRSA by inactivation of PBP2a [24]
Results of bacterial growth curve experiment indicatedthat combination of 9EA-FC-B with ampicillin (both agentsat subinhibitory concentrations) distinctly suppressed thegrowth of MRSA in contrast to MRSA cultures that weretreated with either 9EA-FC-B or ampicillin alone GenerallyMIC of ampicillin reduced when subinhibitory concentra-tion of 9EA-FC-B increased in the combination treatmentFrom the kinetic growth curves we were able to deducethat the antimicrobial action of the ampicillin and 9EA-FC-B combination was observed at the beginning of theexponential phase A very minimal bacterial growth wasseen with increase in incubation hours Instead of growingrapidly during the exponential stage the graph portrayedlow growth of bacterial cells (approximately fourfold lower)with extended lag phase compared to the untreated MRSA(control) An extended lag phase was also detected forMRSAtreatedwith 9EA-FC-B alone Lag phase is the particular stagewhen bacteria equilibrate to adapt to the new environment byundergoing macromolecular repair and synthesis of cellulargrowth through DNA replications [25] Hence we deduced
6 BioMed Research International
that a lengthy lag phase observed in our experiment isdue to the inhibition of DNA replications that delays thecellular growth process The prolonged lag phase of MRSAobserved in this experiment is reminiscent of the action offluoroquinolones that caused inhibition of DNA replicationinMRSA leading to a longer lag phase [26] A potent antimi-crobial action was also identified at the exponential phase inwhich bacterial cells were prevented from growing rapidlyin the presence of 9EA-FC-B alone and in combination withampicillin This phenomenon showed probable interferencein cell division which involves multiple rounds of DNAsynthesis that are controlled by a variety of gene regulatoryprocesses [27 28] The plausible mechanism of action of9EA-FC-B observed at the exponential phase is thereforeassociated with the interruption of cell division that causesmembrane derangements and failure inmembrane functions
As for interpretation of the FIC indices lower indicesindicate better synergism [19] Based on the FIC indicesobtained eight out of the 18 combinations tested showedsynergistic effects The presence of 9EA-FC-B significantlyenhanced the potency of ampicillin by up to 32-fold (MICreduced from 50 120583gmLminus1 in the absence of 9EA-FC-B to156 120583gmLminus1 in the presence of 14 timesMIC of 9EA-FC-B) forMRSA
Ampicillin is a beta-lactam antibiotic that is designedto inhibit PBPs involved in late stage of peptidoglycanbiosynthesis Interference with peptidoglycan biosynthesiscauses deformities in the bacterial cell wall and eventuallyleads to cell death due to high internal osmotic pres-sure Nearly all bacteria can be inhibited by interfering inmechanism of peptidoglycan synthesis [29] Neverthelesstargeting this mechanism is no longer effective due to theproduction PBP2a in MRSA The blocking of normal PBPsby beta-lactams did not exert effects on peptidoglycan orcell wall synthesis because PBP2a replaces their functionand ensures normal formation of cell wall in presence oflethal concentration of beta-lactam drugs [30] However inthis study we experimentally demonstrated restoration ofampicillinrsquos antimicrobial activity by the addition of 9EA-FC-B at subinhibitory level
The synergistic activity observed between 9EA-FC-Band ampicillin against MRSA was shown to be associatedwith inhibition of PBP2a PBP2a is an inducible proteinthat regulates methicillin resistance Its expression is het-erogeneous in nature amidst level of resistance differing tothe beta-lactam being used [31] The mecA gene complexwhich encodes for this protein encompasses the regulatorygenes mecI and mecR Interaction of beta-lactam antibioticswith these regulatory genes eventually allows expression ofmecA in terms of production of PBP2a [32ndash34] As suchthe occurrence of intense PBP2a bands in MRSA culturesgrown in low concentrations of ampicillin compared to thecontrol culture in western blot experiment suggested theinduction of mecA gene transcription In contrast no PBP2abands were detected in MRSA cultures that were exposed to14 times MIC of 9EA-FB-C while PBP2a is only very mildlyexpressed when the concentration of 9EA-FC-B was loweredto 18 timesMIC This suggests that 9EA-FC-B can either inhibitthe production of PBP2a or directly inactivate PBP2a This
suggestion is in agreement with the observation that 9EA-FC-B enhanced the ampicillin susceptibility of MRSA In2004 Shiota et al [24] reported the antibacterial effect ofcorilagin and tellimagrandin I (isolated from Arctostaphylosuva-ursi and Rosa canina resp) against MRSA The twopolyphenolic compounds had exceptionally reduced MICsof beta-lactams for MRSA via inactivation of PBP2a Webelieve that corilagin (a tannin) is also present in the testfraction 9EA-FC-B since corilagin was previously isolatedand identified by our colleagues from the same source of plantmaterials [35] This is further supported by phytochemicaltesting that showed the presence of tannins in 9EA-FC-B(data not shown) Although Shimizu et al [7] previouslyreported that corilagin enhanced antibacterial activity of var-ious beta-lactams by 100- to 2000-fold against MRSA 9EA-FC-B was only shown to enhance the activity of ampicillinby up to 32-fold in the present study The stark differencein antibacterial enhancements can readily be rationalized bythe fact that 9EA-FC-B is a semipure fraction that possiblycontains only a small amount of corilagin in addition tomany other secondarymetabolites that were assumed to havenegligible effect on the antibacterial activity observed
5 Conclusion
From the results reported in this study we found that theantibacterial effect of the fraction 9EA-FC-B obtained fromA wilkesiana and ampicillin is synergistic The synergism isdue the ability of 9EA-FC-B to suppress the production ofPBP2a or directly inactivate it leading to the restoration ofthe susceptibility of MRSA to ampicillin
Conflict of Interests
The authors declared no conflict of interests
Acknowledgment
This study was supported by Science Fund scheme underMinistry of Agriculture and Agro-Based Industry Malaysia(MOA) Malaysia with Grant no M00485401
References
[1] J Snider M Rapids and B Rivard ldquoMethicillin-resistantStaphylococcus aureus a review of current antibiotic therapyrdquoSpectrum Health Grand Rapids 2012
[2] M T Gillespie J W May and R A Skurray ldquoAntibiotic resis-tance in Staphylococcus aureus isolated at an Australian hospitalbetween 1946 and 1981rdquo Journal of Medical Microbiology vol 19no 2 pp 137ndash147 1985
[3] B Berger-Bachi and S Rohrer ldquoFactors influencing methicillinresistance in staphylococcirdquo Archives of Microbiology vol 178no 3 pp 165ndash171 2002
[4] B Berger-Bachi ldquoExpression of resistance to methicillinrdquoTrends in Microbiology vol 2 no 10 pp 389ndash393 1994
[5] J Vouillamoz J M Entenza P Hohl and P MoreillonldquoLB11058 a new cephalosporin with high penicillin-bindingprotein 2a affinity and activity in experimental endocarditis due
BioMed Research International 7
to homogeneously methicillin-resistant Staphylococcus aureusrdquoAntimicrobial Agents and Chemotherapy vol 48 no 11 pp4322ndash4327 2004
[6] S Hemaiswarya A K Kruthiventi and M Doble ldquoSynergismbetween natural products and antibiotics against infectiousdiseasesrdquo Phytomedicine vol 15 no 8 pp 639ndash652 2008
[7] M Shimizu S Shiota T Mizushima et al ldquoMarked poten-tiation of activity of 120573-lactams against methicillin-resistantStaphylococcus aureus by corilaginrdquo Antimicrobial Agents andChemotherapy vol 45 no 11 pp 3198ndash3201 2001
[8] P I Alade andO N Irobi ldquoAntimicrobial activities of crude leafextracts ofAcalypha wilkesianardquo Journal of Ethnopharmacologyvol 39 no 3 pp 171ndash174 1993
[9] B E Akinde andOO Odeyemi ldquoExtraction andmicrobiologi-cal evaluation of the oils from the leaves ofAcalyphawilkesianardquoNigerian Medical Journal vol 17 pp 163ndash165 1987
[10] S W Lim K N Ting T D Bradshaw et al ldquoAcalyphawilkesiana extracts induce apoptosis by causing single strandand double strand DNA breaksrdquo Journal of Ethnopharmacologyvol 138 no 2 pp 616ndash623 2011
[11] S W Lim H S Loh K N Ting T D Bradshaw and N AZeenathul ldquoAcalypha wilkesiana ethyl acetate extract enhancesthe in vitro cytotoxic effects of 120572-tocopherol in human brainand lung cancer cellsrdquo International Journal of BioscienceBiochemistry and Bioinformatics vol 3 no 4 pp 335ndash340 2013
[12] M Othman S Genapathy P S Liew et al ldquoSearch for antibac-terial agents from Malaysian rainforest and tropical plantsrdquoNatural Product Research vol 25 no 19 pp 1857ndash1864 2011
[13] G Spigno and D M de Faveri ldquoAntioxidants from grape stalksandmarc influence of extraction procedure on yield purity andantioxidant power of the extractsrdquo Journal of Food Engineeringvol 78 no 3 pp 793ndash801 2007
[14] MOthmanH S Loh CWiart T J Khoo KH Lim andKNTing ldquoOptimal methods for evaluating antimicrobial activitiesfrom plant extractsrdquo Journal of Microbiological Methods vol 84no 2 pp 161ndash166 2011
[15] Clinical and Laboratory Standards Institute (CLSI) ldquoLabora-tory methods for detection of methicillin-resistant Staphylococ-cus aureus (MRSA)rdquo Supplement M100-S17 2007
[16] JMAndrews ldquoDetermination ofminimum inhibitory concen-trationsrdquo Journal of Antimicrobial Chemotherapy vol 48 no 1pp 5ndash16 2001
[17] D F J Brown ldquoDetection of methicillinoxacillin resistance instaphylococcirdquo Journal of Antimicrobial Chemotherapy vol 48no 1 pp 65ndash70 2001
[18] P Cos A J Vlietinck D V Berghe and LMaes ldquoAnti-infectivepotential of natural products how to develop a stronger in vitroproof-of-conceptrdquo Journal of Ethnopharmacology vol 106 no3 pp 290ndash302 2006
[19] R L White D S Burgess M Manduru and J A BossoldquoComparison of three different in vitro methods of detectingsynergy time-kill checkerboard and E testrdquo AntimicrobialAgents and Chemotherapy vol 40 no 8 pp 1914ndash1918 1996
[20] US Food andDrug Administration (FDA)CDERNewMolecu-lar Entity (NME) Drug and New Biologic Approvals in CalendarYear 2006
[21] F J Schmitz A C Fluit M Luckefahr et al ldquoThe effect ofreserpine an inhibitor of multidrug efflux pumps on the in-vitro activities of ciprofloxacin sparfloxacin and moxifloxacinagainst clinical isolates of Staphylococcus aureusrdquo Journal ofAntimicrobial Chemotherapy vol 42 no 6 pp 807ndash810 1998
[22] F R Stermitz P Lorenz J N Tawara L A Zenewicz and KLewis ldquoSynergy in a medicinal plant Antimicrobial action ofberberine potentiated by 5rsquo-methoxyhydnocarpin a multidrugpump inhibitorrdquoProceedings of theNational Academy of Sciencesof theUnited States of America vol 97 no 4 pp 1433ndash1437 2000
[23] Y Cho N L Schiller and K Oh ldquoAntibacterial effects of greentea polyphenols on clinical isolates of methicillin-resistantStaphylococcus aureusrdquo Current Microbiology vol 57 no 6 pp542ndash546 2008
[24] S Shiota M Shimizu J Sugiyama Y Morita T Mizushimaand T Tsuchiya ldquoMechanisms of action of corilagin andtellimagrandin I that remarkably potentiate the activity of120573-lactams against methicillin-resistant Staphylococcus aureusrdquoMicrobiology and Immunology vol 48 no 1 pp 67ndash73 2004
[25] S Dukan and T Nystrom ldquoBacterial senescence stasis resultsin increased and differential oxidation of cytoplasmic proteinsleading to developmental induction of the heat shock regulonrdquoGenes amp Development vol 12 no 21 pp 3431ndash3441 1998
[26] R A Venezia B E Domaracki A M Evans K E Preston andE M Graffunder ldquoSelection of high-level oxacillin resistancein heteroresistant Staphylococcus aureus by fluoroquinoloneexposurerdquo Journal of Antimicrobial Chemotherapy vol 48 no3 pp 375ndash381 2001
[27] J M Navarro Llorens A Tormo and E Martınez-Garcıa ldquoSta-tionary phase in gram-negative bacteriardquo FEMS MicrobiologyReviews vol 34 no 4 pp 476ndash495 2010
[28] M D Rolfe C J Rice S Lucchini et al ldquoLag phase is a distinctgrowth phase that prepares bacteria for exponential growth andinvolves transient metal accumulationrdquo Journal of Bacteriologyvol 194 no 3 pp 686ndash701 2011
[29] P Lambert ldquoMechanism of action of antibiotics and synthethicanti-infective agentsrdquo in Hugo and Russells PharmaceuticalMicrobiology S PDenyerNAHodges and S PGorman Edspp 202ndash206 Blackwell Publishing Oxford UK 7th edition2004
[30] A Smith ldquoBacterial resistance to antibioticsrdquo inHugo and Rus-sellrsquos Pharmaceutical Microbiology S P Denyer N A Hodgesand S P Gorman Eds pp 220ndash223 Blackwell Oxford UK7th edition 2004
[31] H F Chambers ldquoMethicillin resistance in staphylococcimolec-ular and biochemical basis and clinical implicationsrdquo ClinicalMicrobiology Reviews vol 10 no 4 pp 781ndash791 1997
[32] G L Archer and JM Bosilevac ldquoSignaling antibiotic resistancein staphylococcirdquo Science vol 291 no 5510 pp 1915ndash1916 2001
[33] H Z Zhang C J Hackbarth K M Chansky and H FChambers ldquoA proteolytic transmembrane signaling pathwayand resistance to 120573-lactams in Staphylococcirdquo Science vol 291no 5510 pp 1962ndash1965 2001
[34] S Deresinski ldquoMethicillin-resistant Staphylococcus aureus anevolutionary epidemiologic and therapeutic odysseyrdquo ClinicalInfectious Diseases vol 40 no 4 pp 562ndash573 2005
[35] W M Din J Chu G Clarke et al ldquoAntioxidant and cytopro-tective effects of an ethanol extract of Acalypha wilkesiana varmacafeana from Malaysiardquo Natural Product Communicationsvol 8 no 3 pp 375ndash380 2013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
27 Protein Extraction MRSA was grown in MHB + 2NaCl in the presence of subinhibitory concentrations of 9EA-FC-B until late exponential phase The bacterial lysates wereprepared in an extraction buffer containing Tris and EDTAand culture supernatants were harvested via centrifugation4340 g at 4∘C for 10 min The collected pellets were thentreated with 150mgmLminus1 lysozyme DNase and proteininhibitors cocktail before being subjected to 2 h incubationat 37∘C To enhance cell disruption 15-minute sonication inice-bucket was done Following 15min of centrifugation at13 850 g the pellets were obtained as the insoluble proteinextracts that were harvested in elution buffer containingTris urea and sodium dihydrogen phosphate Protein con-centrations were measured using Pierce 660 nm proteinassay
28 SDS-PAGE and Western Blot Assay Extracted protein(3 120583gmLminus1) was stained with 4X lithium dodecyl sulphate(LDS) sample buffer and subjected to sodium dodecyl sul-fate (SDS)mdashpolyacrylamide (12) gel electrophoresis run at120V Upon completion the gel was stained in CoomassieBlue staining solution until a clear background was obtainedfor scanning with GS-800 calibrated densitometer (Bio-RadUSA) In western blot analyses electrophoresed gels weretransferred to BioTrace NT nitrocellulose transfer mem-brane (Pall USA) Membranes were incubated overnightat 4∘C in gelatin from cold water fish skin (blockingagent) (Sigma USA) The production of PBP2a fromMRSA was detected by probing the membranes with mouseanti-PBP2a primary antibody (Denka Seiken Japan) andantiglyceraldehyde 3-phosphate dehydrogenase (GAPDH)(Thermo Scientific USA) with a dilution factor of 1 10000The same membranes were hybridized with anti-mousehorseradish peroxidase-linked secondary antibody (AbcamUK) diluted to 1 10000 to facilitate colorimetric detectionwith 331015840551015840-tetramethylbenzidine (TMB) substrate (NacalaiTesque Japan) Assay response was recorded using GS-800 calibrated densitometer (Bio-Rad USA) Densitometricquantification of western blot images was done using ImageJ 138 programme (Windows version of NIH Image) Resultswere scored in percentage of expression () normalized toGAPDH control
3 Results
31 Anti-MRSA Activity of Antibiotics and A wilkesianaExtract The MIC values of ampicillin crude extract of Awilkesiana 9EA and fraction 9EA-FC-B for MRSA ATCC43300 and MSSA 11632 are presented in Table 1 The MICvalue of ampicillin against MRSA notably confirmed theresistance of the studied strain whileMIC value of ampicillinagainst MSSA revealed the susceptibility of this strain tothe antibiotic Crude extract 9EA demonstrated antibacterialactivity against both the resistant and sensitive strains testedwith a lower MIC value observed for MSSA The anti-MRSAactivity of fraction 9EA-FC-B was fourfold more effectivethan crude extract 9EA
Table 1 MIC values of ampicillin crude extract 9EA and fraction9EA-FC-B against MRSA and MSSA
StrainMIC
Ampicillin(120583gmLminus1)
9EA(mgmLminus1)
9EA-FC-B(mgmLminus1)
MRSA 50 12 3MSSA 625 6 3Values represent triplicates of three independent experiments
32 Synergistic Effects of 9EA-FC-B with Ampicillin on MRSAGrowth Curves The synergistic effects of 9EA-FC-B withampicillin at subinhibitory concentrations on the growth ofMRSA are shown in Figure 1 The growth inhibitory effectsshown by both ampicillin and 9EA-FC-B on MRSA areconcentration dependent while the curve for the untreatedMRSA culture (control) showed an exponential growthSuppression of MRSA growth was detected when the MRSAcultures were treatedwith 9EA-FC-B alone at 14timesMIC 18timesMICand 116timesMIC (not shown in graph)The growth curvessuggested enhanced growth inhibitory effects when MRSAwas treated with ampicillin at subinhibitory concentrationsin the presence of 9EA-FC-B also at subinhibitory concen-trations The inhibitory effect was observed at as low as 132times MIC ampicillin in the presence of 14 times MIC 9EA-FC-BSimilar inhibitory effect was also achieved in the presenceof 18 times MIC of 9EA-FC-B in combination with 116 times MICampicillin
33 Synergistic Effects of 9EA-FC-B with Ampicillin Based onFIC Index The FIC indices for the tested combinations arepresented in Table 2 Synergistic effects were observed when9EA-FC-B was introduced in the treatment at 14 times MIC18 times MIC and 116 times MIC with ampicillin at subinhibitoryconcentrations (14 to 132 times MIC) In the presence of 14times MIC 9EA-FC-B synergistic effects were observed for thewidest range of subinhibitory concentrations of ampicillinand the range gradually decreased as the subinhibitoryconcentration of 9EA-FC-B was lowered
Analysis of the FIC indices revealed the new MIC valuesof ampicillin in the presence of 9EA-FC-B at subinhibitorylevel (Table 3) 9EA-FC-B at subinhibitory concentrations hasenhanced the activity of ampicillin by up to 32-fold againstMRSA for example MIC of ampicillin alone is 50 120583gmLminus1while the MIC of ampicillin in the presence of 9EA-FC-B at14 timesMIC is 156 120583gmLminus1 It is notable that theMIC values ofampicillin (forMRSA) in the presence of 14 timesMIC 9EA-FC-B and 18 timesMIC 9EA-FC-B are lower than the MIC value ofampicillin for MSSA that is 625 120583gmLminus1
34 Expression of PBP2a Expression of PBP2a was detectedat 76 kDaThe percentage of PBP2a expression in tested treat-ments is summarized in Figure 2 GAPDH which served asan internal control was detected in all treatments (results notshown)The presence of PBP2a band was detected for MRSAcultures that were grown at subinhibitory concentrations ofampicillin (116 times MIC and 132 times MIC) in western blot
4 BioMed Research International
0
01
02
03
04
05
06
07
08
0 2 4 6 8 10 12 14 16 18 20 22 24
Opt
ical
den
sity
at600
nmIncubation time (h)
0
01
02
03
04
05
06
07
08
0 2 4 6 8 10 12 14 16 18 20 22 24
Opt
ical
den
sity
at600
nm
Incubation time (h)
Control MRSA132 times MIC AMP
Control MRSA116 times MIC AMP
09
14 times MIC 9EA-FC-B
18 times MIC 9EA-FC-B
132 times MIC AMP + 14 times MIC 9EA-FC-B
116 times MIC AMP + 18 times MIC 9EA-FC-B
Figure 1 Effects of subinhibitory concentration of ampicillin alone 9EA-FC-B alone and combination treatment on the growth of MRSACell growth wasmeasured by usingOD at 600 nm at indicated time pointsThe curves represent triplicates of three independent experimentsError bars show the standard deviation (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 2 FIC indices of some combinations of ampicillin and 9EA-FC-B for MRSA
Ampicillin (120583gmLminus1) 9EA-FC-B (mgmLminus1)14 timesMIC (075) 18 timesMIC (038) 116 timesMIC (019)
12 timesMIC (25) 075 065 05614 timesMIC (125) 043 036 03118 timesMIC (625) 038 025 mdash116 timesMIC (3125) 031 019 mdash132 timesMIC (1563) 028 mdash mdash164 timesMIC (0781) mdash mdash mdashValues represent triplicates of three independent experiments Index interpretation le05 = synergy gt05 but le40 = indifference and gt4 = antagonism (MIC =minimum inhibitory concentrations mdash = no activity)
experiment Both western blot and quantitative densitomet-ric analysis showed that these cultures have higher expressionof PBP2a compared to the untreatedMRSA culture (control)Inhibition of PBP2a expression was observed when MRSAcultures were exposed to subinhibitory concentrations of9EA-FC-B where no PBP2a bandwas seen at 14 timesMIC 9EA-FC-B and only a very low expression (39) was detected at18 times MIC 9EA-FC-B Likewise the MRSA culture did not
show the presence of PBP2a when grown in the presence of132 timesMIC ampicillin + 14 timesMIC 9EA-FC-B
4 Discussion
The scope of the study is to explore the potential of usingactive plant extracts or fractions to combat resistance in
BioMed Research International 5
L 1 2 3 4 5 6 7
76kDa PBP2a80kDa
(a)
MRS
A
MSS
A
116times
MIC
AM
P
132times
MIC
AM
P
14times
MIC
9EA
-FC-
B
18times
MIC
9EA
-FC-
B
0
10
20
30
40
50
60
70
80
90
100
Expr
essio
n of
PBP
2a (
)
14
MIC
9EA
-F-C
-B132times
MIC
AM
P+
(b)
Figure 2 Expression of penicillin-binding protein 2a (PBP2a) of MRSA cultures grown in the presence of subinhibitory concentrations of9EA-FC-B and ampicillin (a)Western blot image lane L molecular mass markers 1 control MRSA 2 control MSSA 3 116 timesMICAMP 4132 timesMICAMP 5 14timesMIC 9EA-FC-B 6 18timesMIC 9EA-FC-B 7 132 timesMICAMP+ 14timesMIC 9EA-FC-B (b) Quantitative densitometricanalysis of PBP2a expression ofMRSA cultures grown in the presence of ampicillin alone 9EA-FC-B alone and in combinations normalizedto GAPDH loading control (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 3 MIC values of ampicillin in combination with subin-hibitory concentrations of 9EA-FC-B for MRSA
Treatment MIC (120583gmLminus1) of ampicillinAmpicillin alone 50With 9EA-FC-B at 14 timesMIC 156With 9EA-FC-B at 18 timesMIC 313With 9EA-FC-B at 116 timesMIC 125Values represent triplicates of three independent experiments (MIC =minimum inhibitory concentrations)
MRSA For the first time in 2006 US Food and DrugAdministration (FDA 2006) has approved a special green teaextract containing a proprietary mixture of phytochemicals(the active ingredient listed as Polyphenon E) [20] as aprescription drug for the topical (external) treatment ofgenital warts caused by the human papilloma virus (HPV)This example reinforces the efforts to study medicinal plantextracts for possible application in clinical practice Thepresent result established the antimicrobial activity of frac-tion 9EA-FC-B obtained from the ethyl acetate crude extractof A wilkesiana It also demonstrated synergism betweenthe fraction 9EA-FC-B and ampicillin in overcoming resis-tance of MRSA by inhibiting production of PBP2a In thepresence of subinhibitory concentrations of 9EA-FC-B theMIC of ampicillin was reduced by as much as 32-fold from50120583gmLminus1 to 156 120583gmLminus1 indicating that MRSA becamemore sensitive to ampicillin when fraction 9EA-FC-B was
introduced in the treatment Based on these findings wepredict that the active constituents from fraction 9EA-FC-B may potentially be used for combating MRSArsquos virulenceSeveral researches have exploited the synergistic effects ofnatural products for drug development [21ndash23] The mostapt example that is closely related to the present study isthe synergistic effects of corilagin and tellimagrandin I incombination with beta-lactam antibiotics on antibacterialactivity against MRSA by inactivation of PBP2a [24]
Results of bacterial growth curve experiment indicatedthat combination of 9EA-FC-B with ampicillin (both agentsat subinhibitory concentrations) distinctly suppressed thegrowth of MRSA in contrast to MRSA cultures that weretreated with either 9EA-FC-B or ampicillin alone GenerallyMIC of ampicillin reduced when subinhibitory concentra-tion of 9EA-FC-B increased in the combination treatmentFrom the kinetic growth curves we were able to deducethat the antimicrobial action of the ampicillin and 9EA-FC-B combination was observed at the beginning of theexponential phase A very minimal bacterial growth wasseen with increase in incubation hours Instead of growingrapidly during the exponential stage the graph portrayedlow growth of bacterial cells (approximately fourfold lower)with extended lag phase compared to the untreated MRSA(control) An extended lag phase was also detected forMRSAtreatedwith 9EA-FC-B alone Lag phase is the particular stagewhen bacteria equilibrate to adapt to the new environment byundergoing macromolecular repair and synthesis of cellulargrowth through DNA replications [25] Hence we deduced
6 BioMed Research International
that a lengthy lag phase observed in our experiment isdue to the inhibition of DNA replications that delays thecellular growth process The prolonged lag phase of MRSAobserved in this experiment is reminiscent of the action offluoroquinolones that caused inhibition of DNA replicationinMRSA leading to a longer lag phase [26] A potent antimi-crobial action was also identified at the exponential phase inwhich bacterial cells were prevented from growing rapidlyin the presence of 9EA-FC-B alone and in combination withampicillin This phenomenon showed probable interferencein cell division which involves multiple rounds of DNAsynthesis that are controlled by a variety of gene regulatoryprocesses [27 28] The plausible mechanism of action of9EA-FC-B observed at the exponential phase is thereforeassociated with the interruption of cell division that causesmembrane derangements and failure inmembrane functions
As for interpretation of the FIC indices lower indicesindicate better synergism [19] Based on the FIC indicesobtained eight out of the 18 combinations tested showedsynergistic effects The presence of 9EA-FC-B significantlyenhanced the potency of ampicillin by up to 32-fold (MICreduced from 50 120583gmLminus1 in the absence of 9EA-FC-B to156 120583gmLminus1 in the presence of 14 timesMIC of 9EA-FC-B) forMRSA
Ampicillin is a beta-lactam antibiotic that is designedto inhibit PBPs involved in late stage of peptidoglycanbiosynthesis Interference with peptidoglycan biosynthesiscauses deformities in the bacterial cell wall and eventuallyleads to cell death due to high internal osmotic pres-sure Nearly all bacteria can be inhibited by interfering inmechanism of peptidoglycan synthesis [29] Neverthelesstargeting this mechanism is no longer effective due to theproduction PBP2a in MRSA The blocking of normal PBPsby beta-lactams did not exert effects on peptidoglycan orcell wall synthesis because PBP2a replaces their functionand ensures normal formation of cell wall in presence oflethal concentration of beta-lactam drugs [30] However inthis study we experimentally demonstrated restoration ofampicillinrsquos antimicrobial activity by the addition of 9EA-FC-B at subinhibitory level
The synergistic activity observed between 9EA-FC-Band ampicillin against MRSA was shown to be associatedwith inhibition of PBP2a PBP2a is an inducible proteinthat regulates methicillin resistance Its expression is het-erogeneous in nature amidst level of resistance differing tothe beta-lactam being used [31] The mecA gene complexwhich encodes for this protein encompasses the regulatorygenes mecI and mecR Interaction of beta-lactam antibioticswith these regulatory genes eventually allows expression ofmecA in terms of production of PBP2a [32ndash34] As suchthe occurrence of intense PBP2a bands in MRSA culturesgrown in low concentrations of ampicillin compared to thecontrol culture in western blot experiment suggested theinduction of mecA gene transcription In contrast no PBP2abands were detected in MRSA cultures that were exposed to14 times MIC of 9EA-FB-C while PBP2a is only very mildlyexpressed when the concentration of 9EA-FC-B was loweredto 18 timesMIC This suggests that 9EA-FC-B can either inhibitthe production of PBP2a or directly inactivate PBP2a This
suggestion is in agreement with the observation that 9EA-FC-B enhanced the ampicillin susceptibility of MRSA In2004 Shiota et al [24] reported the antibacterial effect ofcorilagin and tellimagrandin I (isolated from Arctostaphylosuva-ursi and Rosa canina resp) against MRSA The twopolyphenolic compounds had exceptionally reduced MICsof beta-lactams for MRSA via inactivation of PBP2a Webelieve that corilagin (a tannin) is also present in the testfraction 9EA-FC-B since corilagin was previously isolatedand identified by our colleagues from the same source of plantmaterials [35] This is further supported by phytochemicaltesting that showed the presence of tannins in 9EA-FC-B(data not shown) Although Shimizu et al [7] previouslyreported that corilagin enhanced antibacterial activity of var-ious beta-lactams by 100- to 2000-fold against MRSA 9EA-FC-B was only shown to enhance the activity of ampicillinby up to 32-fold in the present study The stark differencein antibacterial enhancements can readily be rationalized bythe fact that 9EA-FC-B is a semipure fraction that possiblycontains only a small amount of corilagin in addition tomany other secondarymetabolites that were assumed to havenegligible effect on the antibacterial activity observed
5 Conclusion
From the results reported in this study we found that theantibacterial effect of the fraction 9EA-FC-B obtained fromA wilkesiana and ampicillin is synergistic The synergism isdue the ability of 9EA-FC-B to suppress the production ofPBP2a or directly inactivate it leading to the restoration ofthe susceptibility of MRSA to ampicillin
Conflict of Interests
The authors declared no conflict of interests
Acknowledgment
This study was supported by Science Fund scheme underMinistry of Agriculture and Agro-Based Industry Malaysia(MOA) Malaysia with Grant no M00485401
References
[1] J Snider M Rapids and B Rivard ldquoMethicillin-resistantStaphylococcus aureus a review of current antibiotic therapyrdquoSpectrum Health Grand Rapids 2012
[2] M T Gillespie J W May and R A Skurray ldquoAntibiotic resis-tance in Staphylococcus aureus isolated at an Australian hospitalbetween 1946 and 1981rdquo Journal of Medical Microbiology vol 19no 2 pp 137ndash147 1985
[3] B Berger-Bachi and S Rohrer ldquoFactors influencing methicillinresistance in staphylococcirdquo Archives of Microbiology vol 178no 3 pp 165ndash171 2002
[4] B Berger-Bachi ldquoExpression of resistance to methicillinrdquoTrends in Microbiology vol 2 no 10 pp 389ndash393 1994
[5] J Vouillamoz J M Entenza P Hohl and P MoreillonldquoLB11058 a new cephalosporin with high penicillin-bindingprotein 2a affinity and activity in experimental endocarditis due
BioMed Research International 7
to homogeneously methicillin-resistant Staphylococcus aureusrdquoAntimicrobial Agents and Chemotherapy vol 48 no 11 pp4322ndash4327 2004
[6] S Hemaiswarya A K Kruthiventi and M Doble ldquoSynergismbetween natural products and antibiotics against infectiousdiseasesrdquo Phytomedicine vol 15 no 8 pp 639ndash652 2008
[7] M Shimizu S Shiota T Mizushima et al ldquoMarked poten-tiation of activity of 120573-lactams against methicillin-resistantStaphylococcus aureus by corilaginrdquo Antimicrobial Agents andChemotherapy vol 45 no 11 pp 3198ndash3201 2001
[8] P I Alade andO N Irobi ldquoAntimicrobial activities of crude leafextracts ofAcalypha wilkesianardquo Journal of Ethnopharmacologyvol 39 no 3 pp 171ndash174 1993
[9] B E Akinde andOO Odeyemi ldquoExtraction andmicrobiologi-cal evaluation of the oils from the leaves ofAcalyphawilkesianardquoNigerian Medical Journal vol 17 pp 163ndash165 1987
[10] S W Lim K N Ting T D Bradshaw et al ldquoAcalyphawilkesiana extracts induce apoptosis by causing single strandand double strand DNA breaksrdquo Journal of Ethnopharmacologyvol 138 no 2 pp 616ndash623 2011
[11] S W Lim H S Loh K N Ting T D Bradshaw and N AZeenathul ldquoAcalypha wilkesiana ethyl acetate extract enhancesthe in vitro cytotoxic effects of 120572-tocopherol in human brainand lung cancer cellsrdquo International Journal of BioscienceBiochemistry and Bioinformatics vol 3 no 4 pp 335ndash340 2013
[12] M Othman S Genapathy P S Liew et al ldquoSearch for antibac-terial agents from Malaysian rainforest and tropical plantsrdquoNatural Product Research vol 25 no 19 pp 1857ndash1864 2011
[13] G Spigno and D M de Faveri ldquoAntioxidants from grape stalksandmarc influence of extraction procedure on yield purity andantioxidant power of the extractsrdquo Journal of Food Engineeringvol 78 no 3 pp 793ndash801 2007
[14] MOthmanH S Loh CWiart T J Khoo KH Lim andKNTing ldquoOptimal methods for evaluating antimicrobial activitiesfrom plant extractsrdquo Journal of Microbiological Methods vol 84no 2 pp 161ndash166 2011
[15] Clinical and Laboratory Standards Institute (CLSI) ldquoLabora-tory methods for detection of methicillin-resistant Staphylococ-cus aureus (MRSA)rdquo Supplement M100-S17 2007
[16] JMAndrews ldquoDetermination ofminimum inhibitory concen-trationsrdquo Journal of Antimicrobial Chemotherapy vol 48 no 1pp 5ndash16 2001
[17] D F J Brown ldquoDetection of methicillinoxacillin resistance instaphylococcirdquo Journal of Antimicrobial Chemotherapy vol 48no 1 pp 65ndash70 2001
[18] P Cos A J Vlietinck D V Berghe and LMaes ldquoAnti-infectivepotential of natural products how to develop a stronger in vitroproof-of-conceptrdquo Journal of Ethnopharmacology vol 106 no3 pp 290ndash302 2006
[19] R L White D S Burgess M Manduru and J A BossoldquoComparison of three different in vitro methods of detectingsynergy time-kill checkerboard and E testrdquo AntimicrobialAgents and Chemotherapy vol 40 no 8 pp 1914ndash1918 1996
[20] US Food andDrug Administration (FDA)CDERNewMolecu-lar Entity (NME) Drug and New Biologic Approvals in CalendarYear 2006
[21] F J Schmitz A C Fluit M Luckefahr et al ldquoThe effect ofreserpine an inhibitor of multidrug efflux pumps on the in-vitro activities of ciprofloxacin sparfloxacin and moxifloxacinagainst clinical isolates of Staphylococcus aureusrdquo Journal ofAntimicrobial Chemotherapy vol 42 no 6 pp 807ndash810 1998
[22] F R Stermitz P Lorenz J N Tawara L A Zenewicz and KLewis ldquoSynergy in a medicinal plant Antimicrobial action ofberberine potentiated by 5rsquo-methoxyhydnocarpin a multidrugpump inhibitorrdquoProceedings of theNational Academy of Sciencesof theUnited States of America vol 97 no 4 pp 1433ndash1437 2000
[23] Y Cho N L Schiller and K Oh ldquoAntibacterial effects of greentea polyphenols on clinical isolates of methicillin-resistantStaphylococcus aureusrdquo Current Microbiology vol 57 no 6 pp542ndash546 2008
[24] S Shiota M Shimizu J Sugiyama Y Morita T Mizushimaand T Tsuchiya ldquoMechanisms of action of corilagin andtellimagrandin I that remarkably potentiate the activity of120573-lactams against methicillin-resistant Staphylococcus aureusrdquoMicrobiology and Immunology vol 48 no 1 pp 67ndash73 2004
[25] S Dukan and T Nystrom ldquoBacterial senescence stasis resultsin increased and differential oxidation of cytoplasmic proteinsleading to developmental induction of the heat shock regulonrdquoGenes amp Development vol 12 no 21 pp 3431ndash3441 1998
[26] R A Venezia B E Domaracki A M Evans K E Preston andE M Graffunder ldquoSelection of high-level oxacillin resistancein heteroresistant Staphylococcus aureus by fluoroquinoloneexposurerdquo Journal of Antimicrobial Chemotherapy vol 48 no3 pp 375ndash381 2001
[27] J M Navarro Llorens A Tormo and E Martınez-Garcıa ldquoSta-tionary phase in gram-negative bacteriardquo FEMS MicrobiologyReviews vol 34 no 4 pp 476ndash495 2010
[28] M D Rolfe C J Rice S Lucchini et al ldquoLag phase is a distinctgrowth phase that prepares bacteria for exponential growth andinvolves transient metal accumulationrdquo Journal of Bacteriologyvol 194 no 3 pp 686ndash701 2011
[29] P Lambert ldquoMechanism of action of antibiotics and synthethicanti-infective agentsrdquo in Hugo and Russells PharmaceuticalMicrobiology S PDenyerNAHodges and S PGorman Edspp 202ndash206 Blackwell Publishing Oxford UK 7th edition2004
[30] A Smith ldquoBacterial resistance to antibioticsrdquo inHugo and Rus-sellrsquos Pharmaceutical Microbiology S P Denyer N A Hodgesand S P Gorman Eds pp 220ndash223 Blackwell Oxford UK7th edition 2004
[31] H F Chambers ldquoMethicillin resistance in staphylococcimolec-ular and biochemical basis and clinical implicationsrdquo ClinicalMicrobiology Reviews vol 10 no 4 pp 781ndash791 1997
[32] G L Archer and JM Bosilevac ldquoSignaling antibiotic resistancein staphylococcirdquo Science vol 291 no 5510 pp 1915ndash1916 2001
[33] H Z Zhang C J Hackbarth K M Chansky and H FChambers ldquoA proteolytic transmembrane signaling pathwayand resistance to 120573-lactams in Staphylococcirdquo Science vol 291no 5510 pp 1962ndash1965 2001
[34] S Deresinski ldquoMethicillin-resistant Staphylococcus aureus anevolutionary epidemiologic and therapeutic odysseyrdquo ClinicalInfectious Diseases vol 40 no 4 pp 562ndash573 2005
[35] W M Din J Chu G Clarke et al ldquoAntioxidant and cytopro-tective effects of an ethanol extract of Acalypha wilkesiana varmacafeana from Malaysiardquo Natural Product Communicationsvol 8 no 3 pp 375ndash380 2013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 BioMed Research International
0
01
02
03
04
05
06
07
08
0 2 4 6 8 10 12 14 16 18 20 22 24
Opt
ical
den
sity
at600
nmIncubation time (h)
0
01
02
03
04
05
06
07
08
0 2 4 6 8 10 12 14 16 18 20 22 24
Opt
ical
den
sity
at600
nm
Incubation time (h)
Control MRSA132 times MIC AMP
Control MRSA116 times MIC AMP
09
14 times MIC 9EA-FC-B
18 times MIC 9EA-FC-B
132 times MIC AMP + 14 times MIC 9EA-FC-B
116 times MIC AMP + 18 times MIC 9EA-FC-B
Figure 1 Effects of subinhibitory concentration of ampicillin alone 9EA-FC-B alone and combination treatment on the growth of MRSACell growth wasmeasured by usingOD at 600 nm at indicated time pointsThe curves represent triplicates of three independent experimentsError bars show the standard deviation (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 2 FIC indices of some combinations of ampicillin and 9EA-FC-B for MRSA
Ampicillin (120583gmLminus1) 9EA-FC-B (mgmLminus1)14 timesMIC (075) 18 timesMIC (038) 116 timesMIC (019)
12 timesMIC (25) 075 065 05614 timesMIC (125) 043 036 03118 timesMIC (625) 038 025 mdash116 timesMIC (3125) 031 019 mdash132 timesMIC (1563) 028 mdash mdash164 timesMIC (0781) mdash mdash mdashValues represent triplicates of three independent experiments Index interpretation le05 = synergy gt05 but le40 = indifference and gt4 = antagonism (MIC =minimum inhibitory concentrations mdash = no activity)
experiment Both western blot and quantitative densitomet-ric analysis showed that these cultures have higher expressionof PBP2a compared to the untreatedMRSA culture (control)Inhibition of PBP2a expression was observed when MRSAcultures were exposed to subinhibitory concentrations of9EA-FC-B where no PBP2a bandwas seen at 14 timesMIC 9EA-FC-B and only a very low expression (39) was detected at18 times MIC 9EA-FC-B Likewise the MRSA culture did not
show the presence of PBP2a when grown in the presence of132 timesMIC ampicillin + 14 timesMIC 9EA-FC-B
4 Discussion
The scope of the study is to explore the potential of usingactive plant extracts or fractions to combat resistance in
BioMed Research International 5
L 1 2 3 4 5 6 7
76kDa PBP2a80kDa
(a)
MRS
A
MSS
A
116times
MIC
AM
P
132times
MIC
AM
P
14times
MIC
9EA
-FC-
B
18times
MIC
9EA
-FC-
B
0
10
20
30
40
50
60
70
80
90
100
Expr
essio
n of
PBP
2a (
)
14
MIC
9EA
-F-C
-B132times
MIC
AM
P+
(b)
Figure 2 Expression of penicillin-binding protein 2a (PBP2a) of MRSA cultures grown in the presence of subinhibitory concentrations of9EA-FC-B and ampicillin (a)Western blot image lane L molecular mass markers 1 control MRSA 2 control MSSA 3 116 timesMICAMP 4132 timesMICAMP 5 14timesMIC 9EA-FC-B 6 18timesMIC 9EA-FC-B 7 132 timesMICAMP+ 14timesMIC 9EA-FC-B (b) Quantitative densitometricanalysis of PBP2a expression ofMRSA cultures grown in the presence of ampicillin alone 9EA-FC-B alone and in combinations normalizedto GAPDH loading control (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 3 MIC values of ampicillin in combination with subin-hibitory concentrations of 9EA-FC-B for MRSA
Treatment MIC (120583gmLminus1) of ampicillinAmpicillin alone 50With 9EA-FC-B at 14 timesMIC 156With 9EA-FC-B at 18 timesMIC 313With 9EA-FC-B at 116 timesMIC 125Values represent triplicates of three independent experiments (MIC =minimum inhibitory concentrations)
MRSA For the first time in 2006 US Food and DrugAdministration (FDA 2006) has approved a special green teaextract containing a proprietary mixture of phytochemicals(the active ingredient listed as Polyphenon E) [20] as aprescription drug for the topical (external) treatment ofgenital warts caused by the human papilloma virus (HPV)This example reinforces the efforts to study medicinal plantextracts for possible application in clinical practice Thepresent result established the antimicrobial activity of frac-tion 9EA-FC-B obtained from the ethyl acetate crude extractof A wilkesiana It also demonstrated synergism betweenthe fraction 9EA-FC-B and ampicillin in overcoming resis-tance of MRSA by inhibiting production of PBP2a In thepresence of subinhibitory concentrations of 9EA-FC-B theMIC of ampicillin was reduced by as much as 32-fold from50120583gmLminus1 to 156 120583gmLminus1 indicating that MRSA becamemore sensitive to ampicillin when fraction 9EA-FC-B was
introduced in the treatment Based on these findings wepredict that the active constituents from fraction 9EA-FC-B may potentially be used for combating MRSArsquos virulenceSeveral researches have exploited the synergistic effects ofnatural products for drug development [21ndash23] The mostapt example that is closely related to the present study isthe synergistic effects of corilagin and tellimagrandin I incombination with beta-lactam antibiotics on antibacterialactivity against MRSA by inactivation of PBP2a [24]
Results of bacterial growth curve experiment indicatedthat combination of 9EA-FC-B with ampicillin (both agentsat subinhibitory concentrations) distinctly suppressed thegrowth of MRSA in contrast to MRSA cultures that weretreated with either 9EA-FC-B or ampicillin alone GenerallyMIC of ampicillin reduced when subinhibitory concentra-tion of 9EA-FC-B increased in the combination treatmentFrom the kinetic growth curves we were able to deducethat the antimicrobial action of the ampicillin and 9EA-FC-B combination was observed at the beginning of theexponential phase A very minimal bacterial growth wasseen with increase in incubation hours Instead of growingrapidly during the exponential stage the graph portrayedlow growth of bacterial cells (approximately fourfold lower)with extended lag phase compared to the untreated MRSA(control) An extended lag phase was also detected forMRSAtreatedwith 9EA-FC-B alone Lag phase is the particular stagewhen bacteria equilibrate to adapt to the new environment byundergoing macromolecular repair and synthesis of cellulargrowth through DNA replications [25] Hence we deduced
6 BioMed Research International
that a lengthy lag phase observed in our experiment isdue to the inhibition of DNA replications that delays thecellular growth process The prolonged lag phase of MRSAobserved in this experiment is reminiscent of the action offluoroquinolones that caused inhibition of DNA replicationinMRSA leading to a longer lag phase [26] A potent antimi-crobial action was also identified at the exponential phase inwhich bacterial cells were prevented from growing rapidlyin the presence of 9EA-FC-B alone and in combination withampicillin This phenomenon showed probable interferencein cell division which involves multiple rounds of DNAsynthesis that are controlled by a variety of gene regulatoryprocesses [27 28] The plausible mechanism of action of9EA-FC-B observed at the exponential phase is thereforeassociated with the interruption of cell division that causesmembrane derangements and failure inmembrane functions
As for interpretation of the FIC indices lower indicesindicate better synergism [19] Based on the FIC indicesobtained eight out of the 18 combinations tested showedsynergistic effects The presence of 9EA-FC-B significantlyenhanced the potency of ampicillin by up to 32-fold (MICreduced from 50 120583gmLminus1 in the absence of 9EA-FC-B to156 120583gmLminus1 in the presence of 14 timesMIC of 9EA-FC-B) forMRSA
Ampicillin is a beta-lactam antibiotic that is designedto inhibit PBPs involved in late stage of peptidoglycanbiosynthesis Interference with peptidoglycan biosynthesiscauses deformities in the bacterial cell wall and eventuallyleads to cell death due to high internal osmotic pres-sure Nearly all bacteria can be inhibited by interfering inmechanism of peptidoglycan synthesis [29] Neverthelesstargeting this mechanism is no longer effective due to theproduction PBP2a in MRSA The blocking of normal PBPsby beta-lactams did not exert effects on peptidoglycan orcell wall synthesis because PBP2a replaces their functionand ensures normal formation of cell wall in presence oflethal concentration of beta-lactam drugs [30] However inthis study we experimentally demonstrated restoration ofampicillinrsquos antimicrobial activity by the addition of 9EA-FC-B at subinhibitory level
The synergistic activity observed between 9EA-FC-Band ampicillin against MRSA was shown to be associatedwith inhibition of PBP2a PBP2a is an inducible proteinthat regulates methicillin resistance Its expression is het-erogeneous in nature amidst level of resistance differing tothe beta-lactam being used [31] The mecA gene complexwhich encodes for this protein encompasses the regulatorygenes mecI and mecR Interaction of beta-lactam antibioticswith these regulatory genes eventually allows expression ofmecA in terms of production of PBP2a [32ndash34] As suchthe occurrence of intense PBP2a bands in MRSA culturesgrown in low concentrations of ampicillin compared to thecontrol culture in western blot experiment suggested theinduction of mecA gene transcription In contrast no PBP2abands were detected in MRSA cultures that were exposed to14 times MIC of 9EA-FB-C while PBP2a is only very mildlyexpressed when the concentration of 9EA-FC-B was loweredto 18 timesMIC This suggests that 9EA-FC-B can either inhibitthe production of PBP2a or directly inactivate PBP2a This
suggestion is in agreement with the observation that 9EA-FC-B enhanced the ampicillin susceptibility of MRSA In2004 Shiota et al [24] reported the antibacterial effect ofcorilagin and tellimagrandin I (isolated from Arctostaphylosuva-ursi and Rosa canina resp) against MRSA The twopolyphenolic compounds had exceptionally reduced MICsof beta-lactams for MRSA via inactivation of PBP2a Webelieve that corilagin (a tannin) is also present in the testfraction 9EA-FC-B since corilagin was previously isolatedand identified by our colleagues from the same source of plantmaterials [35] This is further supported by phytochemicaltesting that showed the presence of tannins in 9EA-FC-B(data not shown) Although Shimizu et al [7] previouslyreported that corilagin enhanced antibacterial activity of var-ious beta-lactams by 100- to 2000-fold against MRSA 9EA-FC-B was only shown to enhance the activity of ampicillinby up to 32-fold in the present study The stark differencein antibacterial enhancements can readily be rationalized bythe fact that 9EA-FC-B is a semipure fraction that possiblycontains only a small amount of corilagin in addition tomany other secondarymetabolites that were assumed to havenegligible effect on the antibacterial activity observed
5 Conclusion
From the results reported in this study we found that theantibacterial effect of the fraction 9EA-FC-B obtained fromA wilkesiana and ampicillin is synergistic The synergism isdue the ability of 9EA-FC-B to suppress the production ofPBP2a or directly inactivate it leading to the restoration ofthe susceptibility of MRSA to ampicillin
Conflict of Interests
The authors declared no conflict of interests
Acknowledgment
This study was supported by Science Fund scheme underMinistry of Agriculture and Agro-Based Industry Malaysia(MOA) Malaysia with Grant no M00485401
References
[1] J Snider M Rapids and B Rivard ldquoMethicillin-resistantStaphylococcus aureus a review of current antibiotic therapyrdquoSpectrum Health Grand Rapids 2012
[2] M T Gillespie J W May and R A Skurray ldquoAntibiotic resis-tance in Staphylococcus aureus isolated at an Australian hospitalbetween 1946 and 1981rdquo Journal of Medical Microbiology vol 19no 2 pp 137ndash147 1985
[3] B Berger-Bachi and S Rohrer ldquoFactors influencing methicillinresistance in staphylococcirdquo Archives of Microbiology vol 178no 3 pp 165ndash171 2002
[4] B Berger-Bachi ldquoExpression of resistance to methicillinrdquoTrends in Microbiology vol 2 no 10 pp 389ndash393 1994
[5] J Vouillamoz J M Entenza P Hohl and P MoreillonldquoLB11058 a new cephalosporin with high penicillin-bindingprotein 2a affinity and activity in experimental endocarditis due
BioMed Research International 7
to homogeneously methicillin-resistant Staphylococcus aureusrdquoAntimicrobial Agents and Chemotherapy vol 48 no 11 pp4322ndash4327 2004
[6] S Hemaiswarya A K Kruthiventi and M Doble ldquoSynergismbetween natural products and antibiotics against infectiousdiseasesrdquo Phytomedicine vol 15 no 8 pp 639ndash652 2008
[7] M Shimizu S Shiota T Mizushima et al ldquoMarked poten-tiation of activity of 120573-lactams against methicillin-resistantStaphylococcus aureus by corilaginrdquo Antimicrobial Agents andChemotherapy vol 45 no 11 pp 3198ndash3201 2001
[8] P I Alade andO N Irobi ldquoAntimicrobial activities of crude leafextracts ofAcalypha wilkesianardquo Journal of Ethnopharmacologyvol 39 no 3 pp 171ndash174 1993
[9] B E Akinde andOO Odeyemi ldquoExtraction andmicrobiologi-cal evaluation of the oils from the leaves ofAcalyphawilkesianardquoNigerian Medical Journal vol 17 pp 163ndash165 1987
[10] S W Lim K N Ting T D Bradshaw et al ldquoAcalyphawilkesiana extracts induce apoptosis by causing single strandand double strand DNA breaksrdquo Journal of Ethnopharmacologyvol 138 no 2 pp 616ndash623 2011
[11] S W Lim H S Loh K N Ting T D Bradshaw and N AZeenathul ldquoAcalypha wilkesiana ethyl acetate extract enhancesthe in vitro cytotoxic effects of 120572-tocopherol in human brainand lung cancer cellsrdquo International Journal of BioscienceBiochemistry and Bioinformatics vol 3 no 4 pp 335ndash340 2013
[12] M Othman S Genapathy P S Liew et al ldquoSearch for antibac-terial agents from Malaysian rainforest and tropical plantsrdquoNatural Product Research vol 25 no 19 pp 1857ndash1864 2011
[13] G Spigno and D M de Faveri ldquoAntioxidants from grape stalksandmarc influence of extraction procedure on yield purity andantioxidant power of the extractsrdquo Journal of Food Engineeringvol 78 no 3 pp 793ndash801 2007
[14] MOthmanH S Loh CWiart T J Khoo KH Lim andKNTing ldquoOptimal methods for evaluating antimicrobial activitiesfrom plant extractsrdquo Journal of Microbiological Methods vol 84no 2 pp 161ndash166 2011
[15] Clinical and Laboratory Standards Institute (CLSI) ldquoLabora-tory methods for detection of methicillin-resistant Staphylococ-cus aureus (MRSA)rdquo Supplement M100-S17 2007
[16] JMAndrews ldquoDetermination ofminimum inhibitory concen-trationsrdquo Journal of Antimicrobial Chemotherapy vol 48 no 1pp 5ndash16 2001
[17] D F J Brown ldquoDetection of methicillinoxacillin resistance instaphylococcirdquo Journal of Antimicrobial Chemotherapy vol 48no 1 pp 65ndash70 2001
[18] P Cos A J Vlietinck D V Berghe and LMaes ldquoAnti-infectivepotential of natural products how to develop a stronger in vitroproof-of-conceptrdquo Journal of Ethnopharmacology vol 106 no3 pp 290ndash302 2006
[19] R L White D S Burgess M Manduru and J A BossoldquoComparison of three different in vitro methods of detectingsynergy time-kill checkerboard and E testrdquo AntimicrobialAgents and Chemotherapy vol 40 no 8 pp 1914ndash1918 1996
[20] US Food andDrug Administration (FDA)CDERNewMolecu-lar Entity (NME) Drug and New Biologic Approvals in CalendarYear 2006
[21] F J Schmitz A C Fluit M Luckefahr et al ldquoThe effect ofreserpine an inhibitor of multidrug efflux pumps on the in-vitro activities of ciprofloxacin sparfloxacin and moxifloxacinagainst clinical isolates of Staphylococcus aureusrdquo Journal ofAntimicrobial Chemotherapy vol 42 no 6 pp 807ndash810 1998
[22] F R Stermitz P Lorenz J N Tawara L A Zenewicz and KLewis ldquoSynergy in a medicinal plant Antimicrobial action ofberberine potentiated by 5rsquo-methoxyhydnocarpin a multidrugpump inhibitorrdquoProceedings of theNational Academy of Sciencesof theUnited States of America vol 97 no 4 pp 1433ndash1437 2000
[23] Y Cho N L Schiller and K Oh ldquoAntibacterial effects of greentea polyphenols on clinical isolates of methicillin-resistantStaphylococcus aureusrdquo Current Microbiology vol 57 no 6 pp542ndash546 2008
[24] S Shiota M Shimizu J Sugiyama Y Morita T Mizushimaand T Tsuchiya ldquoMechanisms of action of corilagin andtellimagrandin I that remarkably potentiate the activity of120573-lactams against methicillin-resistant Staphylococcus aureusrdquoMicrobiology and Immunology vol 48 no 1 pp 67ndash73 2004
[25] S Dukan and T Nystrom ldquoBacterial senescence stasis resultsin increased and differential oxidation of cytoplasmic proteinsleading to developmental induction of the heat shock regulonrdquoGenes amp Development vol 12 no 21 pp 3431ndash3441 1998
[26] R A Venezia B E Domaracki A M Evans K E Preston andE M Graffunder ldquoSelection of high-level oxacillin resistancein heteroresistant Staphylococcus aureus by fluoroquinoloneexposurerdquo Journal of Antimicrobial Chemotherapy vol 48 no3 pp 375ndash381 2001
[27] J M Navarro Llorens A Tormo and E Martınez-Garcıa ldquoSta-tionary phase in gram-negative bacteriardquo FEMS MicrobiologyReviews vol 34 no 4 pp 476ndash495 2010
[28] M D Rolfe C J Rice S Lucchini et al ldquoLag phase is a distinctgrowth phase that prepares bacteria for exponential growth andinvolves transient metal accumulationrdquo Journal of Bacteriologyvol 194 no 3 pp 686ndash701 2011
[29] P Lambert ldquoMechanism of action of antibiotics and synthethicanti-infective agentsrdquo in Hugo and Russells PharmaceuticalMicrobiology S PDenyerNAHodges and S PGorman Edspp 202ndash206 Blackwell Publishing Oxford UK 7th edition2004
[30] A Smith ldquoBacterial resistance to antibioticsrdquo inHugo and Rus-sellrsquos Pharmaceutical Microbiology S P Denyer N A Hodgesand S P Gorman Eds pp 220ndash223 Blackwell Oxford UK7th edition 2004
[31] H F Chambers ldquoMethicillin resistance in staphylococcimolec-ular and biochemical basis and clinical implicationsrdquo ClinicalMicrobiology Reviews vol 10 no 4 pp 781ndash791 1997
[32] G L Archer and JM Bosilevac ldquoSignaling antibiotic resistancein staphylococcirdquo Science vol 291 no 5510 pp 1915ndash1916 2001
[33] H Z Zhang C J Hackbarth K M Chansky and H FChambers ldquoA proteolytic transmembrane signaling pathwayand resistance to 120573-lactams in Staphylococcirdquo Science vol 291no 5510 pp 1962ndash1965 2001
[34] S Deresinski ldquoMethicillin-resistant Staphylococcus aureus anevolutionary epidemiologic and therapeutic odysseyrdquo ClinicalInfectious Diseases vol 40 no 4 pp 562ndash573 2005
[35] W M Din J Chu G Clarke et al ldquoAntioxidant and cytopro-tective effects of an ethanol extract of Acalypha wilkesiana varmacafeana from Malaysiardquo Natural Product Communicationsvol 8 no 3 pp 375ndash380 2013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
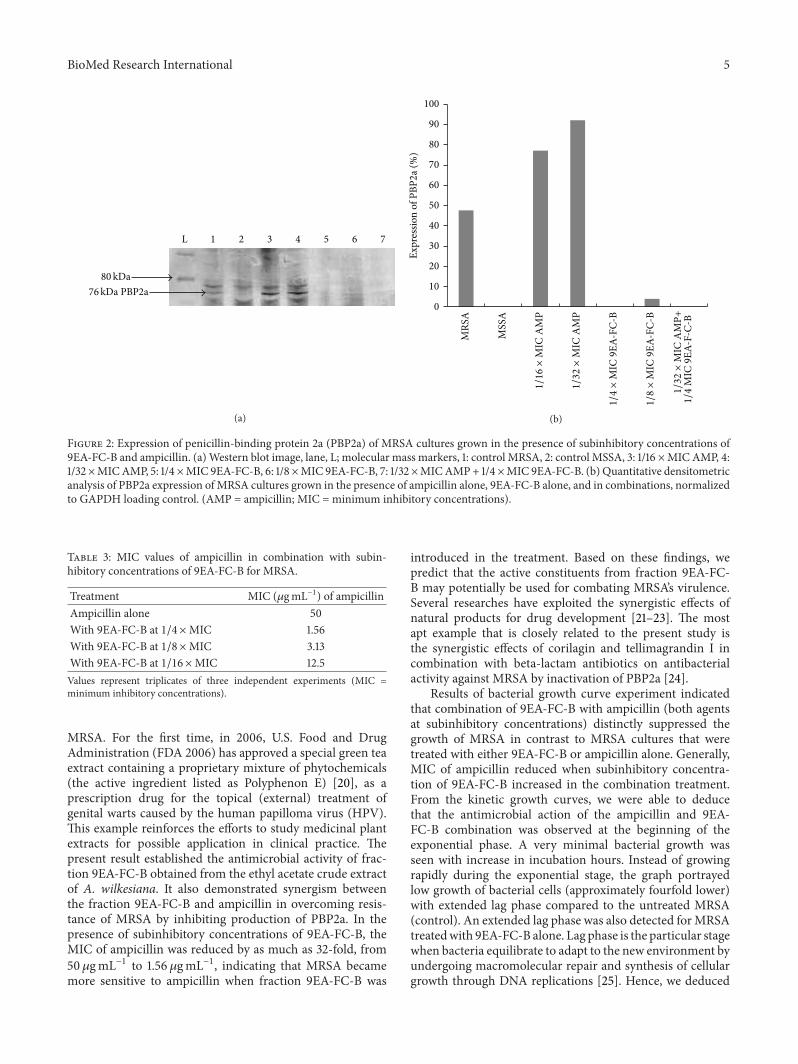
BioMed Research International 5
L 1 2 3 4 5 6 7
76kDa PBP2a80kDa
(a)
MRS
A
MSS
A
116times
MIC
AM
P
132times
MIC
AM
P
14times
MIC
9EA
-FC-
B
18times
MIC
9EA
-FC-
B
0
10
20
30
40
50
60
70
80
90
100
Expr
essio
n of
PBP
2a (
)
14
MIC
9EA
-F-C
-B132times
MIC
AM
P+
(b)
Figure 2 Expression of penicillin-binding protein 2a (PBP2a) of MRSA cultures grown in the presence of subinhibitory concentrations of9EA-FC-B and ampicillin (a)Western blot image lane L molecular mass markers 1 control MRSA 2 control MSSA 3 116 timesMICAMP 4132 timesMICAMP 5 14timesMIC 9EA-FC-B 6 18timesMIC 9EA-FC-B 7 132 timesMICAMP+ 14timesMIC 9EA-FC-B (b) Quantitative densitometricanalysis of PBP2a expression ofMRSA cultures grown in the presence of ampicillin alone 9EA-FC-B alone and in combinations normalizedto GAPDH loading control (AMP = ampicillin MIC = minimum inhibitory concentrations)
Table 3 MIC values of ampicillin in combination with subin-hibitory concentrations of 9EA-FC-B for MRSA
Treatment MIC (120583gmLminus1) of ampicillinAmpicillin alone 50With 9EA-FC-B at 14 timesMIC 156With 9EA-FC-B at 18 timesMIC 313With 9EA-FC-B at 116 timesMIC 125Values represent triplicates of three independent experiments (MIC =minimum inhibitory concentrations)
MRSA For the first time in 2006 US Food and DrugAdministration (FDA 2006) has approved a special green teaextract containing a proprietary mixture of phytochemicals(the active ingredient listed as Polyphenon E) [20] as aprescription drug for the topical (external) treatment ofgenital warts caused by the human papilloma virus (HPV)This example reinforces the efforts to study medicinal plantextracts for possible application in clinical practice Thepresent result established the antimicrobial activity of frac-tion 9EA-FC-B obtained from the ethyl acetate crude extractof A wilkesiana It also demonstrated synergism betweenthe fraction 9EA-FC-B and ampicillin in overcoming resis-tance of MRSA by inhibiting production of PBP2a In thepresence of subinhibitory concentrations of 9EA-FC-B theMIC of ampicillin was reduced by as much as 32-fold from50120583gmLminus1 to 156 120583gmLminus1 indicating that MRSA becamemore sensitive to ampicillin when fraction 9EA-FC-B was
introduced in the treatment Based on these findings wepredict that the active constituents from fraction 9EA-FC-B may potentially be used for combating MRSArsquos virulenceSeveral researches have exploited the synergistic effects ofnatural products for drug development [21ndash23] The mostapt example that is closely related to the present study isthe synergistic effects of corilagin and tellimagrandin I incombination with beta-lactam antibiotics on antibacterialactivity against MRSA by inactivation of PBP2a [24]
Results of bacterial growth curve experiment indicatedthat combination of 9EA-FC-B with ampicillin (both agentsat subinhibitory concentrations) distinctly suppressed thegrowth of MRSA in contrast to MRSA cultures that weretreated with either 9EA-FC-B or ampicillin alone GenerallyMIC of ampicillin reduced when subinhibitory concentra-tion of 9EA-FC-B increased in the combination treatmentFrom the kinetic growth curves we were able to deducethat the antimicrobial action of the ampicillin and 9EA-FC-B combination was observed at the beginning of theexponential phase A very minimal bacterial growth wasseen with increase in incubation hours Instead of growingrapidly during the exponential stage the graph portrayedlow growth of bacterial cells (approximately fourfold lower)with extended lag phase compared to the untreated MRSA(control) An extended lag phase was also detected forMRSAtreatedwith 9EA-FC-B alone Lag phase is the particular stagewhen bacteria equilibrate to adapt to the new environment byundergoing macromolecular repair and synthesis of cellulargrowth through DNA replications [25] Hence we deduced
6 BioMed Research International
that a lengthy lag phase observed in our experiment isdue to the inhibition of DNA replications that delays thecellular growth process The prolonged lag phase of MRSAobserved in this experiment is reminiscent of the action offluoroquinolones that caused inhibition of DNA replicationinMRSA leading to a longer lag phase [26] A potent antimi-crobial action was also identified at the exponential phase inwhich bacterial cells were prevented from growing rapidlyin the presence of 9EA-FC-B alone and in combination withampicillin This phenomenon showed probable interferencein cell division which involves multiple rounds of DNAsynthesis that are controlled by a variety of gene regulatoryprocesses [27 28] The plausible mechanism of action of9EA-FC-B observed at the exponential phase is thereforeassociated with the interruption of cell division that causesmembrane derangements and failure inmembrane functions
As for interpretation of the FIC indices lower indicesindicate better synergism [19] Based on the FIC indicesobtained eight out of the 18 combinations tested showedsynergistic effects The presence of 9EA-FC-B significantlyenhanced the potency of ampicillin by up to 32-fold (MICreduced from 50 120583gmLminus1 in the absence of 9EA-FC-B to156 120583gmLminus1 in the presence of 14 timesMIC of 9EA-FC-B) forMRSA
Ampicillin is a beta-lactam antibiotic that is designedto inhibit PBPs involved in late stage of peptidoglycanbiosynthesis Interference with peptidoglycan biosynthesiscauses deformities in the bacterial cell wall and eventuallyleads to cell death due to high internal osmotic pres-sure Nearly all bacteria can be inhibited by interfering inmechanism of peptidoglycan synthesis [29] Neverthelesstargeting this mechanism is no longer effective due to theproduction PBP2a in MRSA The blocking of normal PBPsby beta-lactams did not exert effects on peptidoglycan orcell wall synthesis because PBP2a replaces their functionand ensures normal formation of cell wall in presence oflethal concentration of beta-lactam drugs [30] However inthis study we experimentally demonstrated restoration ofampicillinrsquos antimicrobial activity by the addition of 9EA-FC-B at subinhibitory level
The synergistic activity observed between 9EA-FC-Band ampicillin against MRSA was shown to be associatedwith inhibition of PBP2a PBP2a is an inducible proteinthat regulates methicillin resistance Its expression is het-erogeneous in nature amidst level of resistance differing tothe beta-lactam being used [31] The mecA gene complexwhich encodes for this protein encompasses the regulatorygenes mecI and mecR Interaction of beta-lactam antibioticswith these regulatory genes eventually allows expression ofmecA in terms of production of PBP2a [32ndash34] As suchthe occurrence of intense PBP2a bands in MRSA culturesgrown in low concentrations of ampicillin compared to thecontrol culture in western blot experiment suggested theinduction of mecA gene transcription In contrast no PBP2abands were detected in MRSA cultures that were exposed to14 times MIC of 9EA-FB-C while PBP2a is only very mildlyexpressed when the concentration of 9EA-FC-B was loweredto 18 timesMIC This suggests that 9EA-FC-B can either inhibitthe production of PBP2a or directly inactivate PBP2a This
suggestion is in agreement with the observation that 9EA-FC-B enhanced the ampicillin susceptibility of MRSA In2004 Shiota et al [24] reported the antibacterial effect ofcorilagin and tellimagrandin I (isolated from Arctostaphylosuva-ursi and Rosa canina resp) against MRSA The twopolyphenolic compounds had exceptionally reduced MICsof beta-lactams for MRSA via inactivation of PBP2a Webelieve that corilagin (a tannin) is also present in the testfraction 9EA-FC-B since corilagin was previously isolatedand identified by our colleagues from the same source of plantmaterials [35] This is further supported by phytochemicaltesting that showed the presence of tannins in 9EA-FC-B(data not shown) Although Shimizu et al [7] previouslyreported that corilagin enhanced antibacterial activity of var-ious beta-lactams by 100- to 2000-fold against MRSA 9EA-FC-B was only shown to enhance the activity of ampicillinby up to 32-fold in the present study The stark differencein antibacterial enhancements can readily be rationalized bythe fact that 9EA-FC-B is a semipure fraction that possiblycontains only a small amount of corilagin in addition tomany other secondarymetabolites that were assumed to havenegligible effect on the antibacterial activity observed
5 Conclusion
From the results reported in this study we found that theantibacterial effect of the fraction 9EA-FC-B obtained fromA wilkesiana and ampicillin is synergistic The synergism isdue the ability of 9EA-FC-B to suppress the production ofPBP2a or directly inactivate it leading to the restoration ofthe susceptibility of MRSA to ampicillin
Conflict of Interests
The authors declared no conflict of interests
Acknowledgment
This study was supported by Science Fund scheme underMinistry of Agriculture and Agro-Based Industry Malaysia(MOA) Malaysia with Grant no M00485401
References
[1] J Snider M Rapids and B Rivard ldquoMethicillin-resistantStaphylococcus aureus a review of current antibiotic therapyrdquoSpectrum Health Grand Rapids 2012
[2] M T Gillespie J W May and R A Skurray ldquoAntibiotic resis-tance in Staphylococcus aureus isolated at an Australian hospitalbetween 1946 and 1981rdquo Journal of Medical Microbiology vol 19no 2 pp 137ndash147 1985
[3] B Berger-Bachi and S Rohrer ldquoFactors influencing methicillinresistance in staphylococcirdquo Archives of Microbiology vol 178no 3 pp 165ndash171 2002
[4] B Berger-Bachi ldquoExpression of resistance to methicillinrdquoTrends in Microbiology vol 2 no 10 pp 389ndash393 1994
[5] J Vouillamoz J M Entenza P Hohl and P MoreillonldquoLB11058 a new cephalosporin with high penicillin-bindingprotein 2a affinity and activity in experimental endocarditis due
BioMed Research International 7
to homogeneously methicillin-resistant Staphylococcus aureusrdquoAntimicrobial Agents and Chemotherapy vol 48 no 11 pp4322ndash4327 2004
[6] S Hemaiswarya A K Kruthiventi and M Doble ldquoSynergismbetween natural products and antibiotics against infectiousdiseasesrdquo Phytomedicine vol 15 no 8 pp 639ndash652 2008
[7] M Shimizu S Shiota T Mizushima et al ldquoMarked poten-tiation of activity of 120573-lactams against methicillin-resistantStaphylococcus aureus by corilaginrdquo Antimicrobial Agents andChemotherapy vol 45 no 11 pp 3198ndash3201 2001
[8] P I Alade andO N Irobi ldquoAntimicrobial activities of crude leafextracts ofAcalypha wilkesianardquo Journal of Ethnopharmacologyvol 39 no 3 pp 171ndash174 1993
[9] B E Akinde andOO Odeyemi ldquoExtraction andmicrobiologi-cal evaluation of the oils from the leaves ofAcalyphawilkesianardquoNigerian Medical Journal vol 17 pp 163ndash165 1987
[10] S W Lim K N Ting T D Bradshaw et al ldquoAcalyphawilkesiana extracts induce apoptosis by causing single strandand double strand DNA breaksrdquo Journal of Ethnopharmacologyvol 138 no 2 pp 616ndash623 2011
[11] S W Lim H S Loh K N Ting T D Bradshaw and N AZeenathul ldquoAcalypha wilkesiana ethyl acetate extract enhancesthe in vitro cytotoxic effects of 120572-tocopherol in human brainand lung cancer cellsrdquo International Journal of BioscienceBiochemistry and Bioinformatics vol 3 no 4 pp 335ndash340 2013
[12] M Othman S Genapathy P S Liew et al ldquoSearch for antibac-terial agents from Malaysian rainforest and tropical plantsrdquoNatural Product Research vol 25 no 19 pp 1857ndash1864 2011
[13] G Spigno and D M de Faveri ldquoAntioxidants from grape stalksandmarc influence of extraction procedure on yield purity andantioxidant power of the extractsrdquo Journal of Food Engineeringvol 78 no 3 pp 793ndash801 2007
[14] MOthmanH S Loh CWiart T J Khoo KH Lim andKNTing ldquoOptimal methods for evaluating antimicrobial activitiesfrom plant extractsrdquo Journal of Microbiological Methods vol 84no 2 pp 161ndash166 2011
[15] Clinical and Laboratory Standards Institute (CLSI) ldquoLabora-tory methods for detection of methicillin-resistant Staphylococ-cus aureus (MRSA)rdquo Supplement M100-S17 2007
[16] JMAndrews ldquoDetermination ofminimum inhibitory concen-trationsrdquo Journal of Antimicrobial Chemotherapy vol 48 no 1pp 5ndash16 2001
[17] D F J Brown ldquoDetection of methicillinoxacillin resistance instaphylococcirdquo Journal of Antimicrobial Chemotherapy vol 48no 1 pp 65ndash70 2001
[18] P Cos A J Vlietinck D V Berghe and LMaes ldquoAnti-infectivepotential of natural products how to develop a stronger in vitroproof-of-conceptrdquo Journal of Ethnopharmacology vol 106 no3 pp 290ndash302 2006
[19] R L White D S Burgess M Manduru and J A BossoldquoComparison of three different in vitro methods of detectingsynergy time-kill checkerboard and E testrdquo AntimicrobialAgents and Chemotherapy vol 40 no 8 pp 1914ndash1918 1996
[20] US Food andDrug Administration (FDA)CDERNewMolecu-lar Entity (NME) Drug and New Biologic Approvals in CalendarYear 2006
[21] F J Schmitz A C Fluit M Luckefahr et al ldquoThe effect ofreserpine an inhibitor of multidrug efflux pumps on the in-vitro activities of ciprofloxacin sparfloxacin and moxifloxacinagainst clinical isolates of Staphylococcus aureusrdquo Journal ofAntimicrobial Chemotherapy vol 42 no 6 pp 807ndash810 1998
[22] F R Stermitz P Lorenz J N Tawara L A Zenewicz and KLewis ldquoSynergy in a medicinal plant Antimicrobial action ofberberine potentiated by 5rsquo-methoxyhydnocarpin a multidrugpump inhibitorrdquoProceedings of theNational Academy of Sciencesof theUnited States of America vol 97 no 4 pp 1433ndash1437 2000
[23] Y Cho N L Schiller and K Oh ldquoAntibacterial effects of greentea polyphenols on clinical isolates of methicillin-resistantStaphylococcus aureusrdquo Current Microbiology vol 57 no 6 pp542ndash546 2008
[24] S Shiota M Shimizu J Sugiyama Y Morita T Mizushimaand T Tsuchiya ldquoMechanisms of action of corilagin andtellimagrandin I that remarkably potentiate the activity of120573-lactams against methicillin-resistant Staphylococcus aureusrdquoMicrobiology and Immunology vol 48 no 1 pp 67ndash73 2004
[25] S Dukan and T Nystrom ldquoBacterial senescence stasis resultsin increased and differential oxidation of cytoplasmic proteinsleading to developmental induction of the heat shock regulonrdquoGenes amp Development vol 12 no 21 pp 3431ndash3441 1998
[26] R A Venezia B E Domaracki A M Evans K E Preston andE M Graffunder ldquoSelection of high-level oxacillin resistancein heteroresistant Staphylococcus aureus by fluoroquinoloneexposurerdquo Journal of Antimicrobial Chemotherapy vol 48 no3 pp 375ndash381 2001
[27] J M Navarro Llorens A Tormo and E Martınez-Garcıa ldquoSta-tionary phase in gram-negative bacteriardquo FEMS MicrobiologyReviews vol 34 no 4 pp 476ndash495 2010
[28] M D Rolfe C J Rice S Lucchini et al ldquoLag phase is a distinctgrowth phase that prepares bacteria for exponential growth andinvolves transient metal accumulationrdquo Journal of Bacteriologyvol 194 no 3 pp 686ndash701 2011
[29] P Lambert ldquoMechanism of action of antibiotics and synthethicanti-infective agentsrdquo in Hugo and Russells PharmaceuticalMicrobiology S PDenyerNAHodges and S PGorman Edspp 202ndash206 Blackwell Publishing Oxford UK 7th edition2004
[30] A Smith ldquoBacterial resistance to antibioticsrdquo inHugo and Rus-sellrsquos Pharmaceutical Microbiology S P Denyer N A Hodgesand S P Gorman Eds pp 220ndash223 Blackwell Oxford UK7th edition 2004
[31] H F Chambers ldquoMethicillin resistance in staphylococcimolec-ular and biochemical basis and clinical implicationsrdquo ClinicalMicrobiology Reviews vol 10 no 4 pp 781ndash791 1997
[32] G L Archer and JM Bosilevac ldquoSignaling antibiotic resistancein staphylococcirdquo Science vol 291 no 5510 pp 1915ndash1916 2001
[33] H Z Zhang C J Hackbarth K M Chansky and H FChambers ldquoA proteolytic transmembrane signaling pathwayand resistance to 120573-lactams in Staphylococcirdquo Science vol 291no 5510 pp 1962ndash1965 2001
[34] S Deresinski ldquoMethicillin-resistant Staphylococcus aureus anevolutionary epidemiologic and therapeutic odysseyrdquo ClinicalInfectious Diseases vol 40 no 4 pp 562ndash573 2005
[35] W M Din J Chu G Clarke et al ldquoAntioxidant and cytopro-tective effects of an ethanol extract of Acalypha wilkesiana varmacafeana from Malaysiardquo Natural Product Communicationsvol 8 no 3 pp 375ndash380 2013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 BioMed Research International
that a lengthy lag phase observed in our experiment isdue to the inhibition of DNA replications that delays thecellular growth process The prolonged lag phase of MRSAobserved in this experiment is reminiscent of the action offluoroquinolones that caused inhibition of DNA replicationinMRSA leading to a longer lag phase [26] A potent antimi-crobial action was also identified at the exponential phase inwhich bacterial cells were prevented from growing rapidlyin the presence of 9EA-FC-B alone and in combination withampicillin This phenomenon showed probable interferencein cell division which involves multiple rounds of DNAsynthesis that are controlled by a variety of gene regulatoryprocesses [27 28] The plausible mechanism of action of9EA-FC-B observed at the exponential phase is thereforeassociated with the interruption of cell division that causesmembrane derangements and failure inmembrane functions
As for interpretation of the FIC indices lower indicesindicate better synergism [19] Based on the FIC indicesobtained eight out of the 18 combinations tested showedsynergistic effects The presence of 9EA-FC-B significantlyenhanced the potency of ampicillin by up to 32-fold (MICreduced from 50 120583gmLminus1 in the absence of 9EA-FC-B to156 120583gmLminus1 in the presence of 14 timesMIC of 9EA-FC-B) forMRSA
Ampicillin is a beta-lactam antibiotic that is designedto inhibit PBPs involved in late stage of peptidoglycanbiosynthesis Interference with peptidoglycan biosynthesiscauses deformities in the bacterial cell wall and eventuallyleads to cell death due to high internal osmotic pres-sure Nearly all bacteria can be inhibited by interfering inmechanism of peptidoglycan synthesis [29] Neverthelesstargeting this mechanism is no longer effective due to theproduction PBP2a in MRSA The blocking of normal PBPsby beta-lactams did not exert effects on peptidoglycan orcell wall synthesis because PBP2a replaces their functionand ensures normal formation of cell wall in presence oflethal concentration of beta-lactam drugs [30] However inthis study we experimentally demonstrated restoration ofampicillinrsquos antimicrobial activity by the addition of 9EA-FC-B at subinhibitory level
The synergistic activity observed between 9EA-FC-Band ampicillin against MRSA was shown to be associatedwith inhibition of PBP2a PBP2a is an inducible proteinthat regulates methicillin resistance Its expression is het-erogeneous in nature amidst level of resistance differing tothe beta-lactam being used [31] The mecA gene complexwhich encodes for this protein encompasses the regulatorygenes mecI and mecR Interaction of beta-lactam antibioticswith these regulatory genes eventually allows expression ofmecA in terms of production of PBP2a [32ndash34] As suchthe occurrence of intense PBP2a bands in MRSA culturesgrown in low concentrations of ampicillin compared to thecontrol culture in western blot experiment suggested theinduction of mecA gene transcription In contrast no PBP2abands were detected in MRSA cultures that were exposed to14 times MIC of 9EA-FB-C while PBP2a is only very mildlyexpressed when the concentration of 9EA-FC-B was loweredto 18 timesMIC This suggests that 9EA-FC-B can either inhibitthe production of PBP2a or directly inactivate PBP2a This
suggestion is in agreement with the observation that 9EA-FC-B enhanced the ampicillin susceptibility of MRSA In2004 Shiota et al [24] reported the antibacterial effect ofcorilagin and tellimagrandin I (isolated from Arctostaphylosuva-ursi and Rosa canina resp) against MRSA The twopolyphenolic compounds had exceptionally reduced MICsof beta-lactams for MRSA via inactivation of PBP2a Webelieve that corilagin (a tannin) is also present in the testfraction 9EA-FC-B since corilagin was previously isolatedand identified by our colleagues from the same source of plantmaterials [35] This is further supported by phytochemicaltesting that showed the presence of tannins in 9EA-FC-B(data not shown) Although Shimizu et al [7] previouslyreported that corilagin enhanced antibacterial activity of var-ious beta-lactams by 100- to 2000-fold against MRSA 9EA-FC-B was only shown to enhance the activity of ampicillinby up to 32-fold in the present study The stark differencein antibacterial enhancements can readily be rationalized bythe fact that 9EA-FC-B is a semipure fraction that possiblycontains only a small amount of corilagin in addition tomany other secondarymetabolites that were assumed to havenegligible effect on the antibacterial activity observed
5 Conclusion
From the results reported in this study we found that theantibacterial effect of the fraction 9EA-FC-B obtained fromA wilkesiana and ampicillin is synergistic The synergism isdue the ability of 9EA-FC-B to suppress the production ofPBP2a or directly inactivate it leading to the restoration ofthe susceptibility of MRSA to ampicillin
Conflict of Interests
The authors declared no conflict of interests
Acknowledgment
This study was supported by Science Fund scheme underMinistry of Agriculture and Agro-Based Industry Malaysia(MOA) Malaysia with Grant no M00485401
References
[1] J Snider M Rapids and B Rivard ldquoMethicillin-resistantStaphylococcus aureus a review of current antibiotic therapyrdquoSpectrum Health Grand Rapids 2012
[2] M T Gillespie J W May and R A Skurray ldquoAntibiotic resis-tance in Staphylococcus aureus isolated at an Australian hospitalbetween 1946 and 1981rdquo Journal of Medical Microbiology vol 19no 2 pp 137ndash147 1985
[3] B Berger-Bachi and S Rohrer ldquoFactors influencing methicillinresistance in staphylococcirdquo Archives of Microbiology vol 178no 3 pp 165ndash171 2002
[4] B Berger-Bachi ldquoExpression of resistance to methicillinrdquoTrends in Microbiology vol 2 no 10 pp 389ndash393 1994
[5] J Vouillamoz J M Entenza P Hohl and P MoreillonldquoLB11058 a new cephalosporin with high penicillin-bindingprotein 2a affinity and activity in experimental endocarditis due
BioMed Research International 7
to homogeneously methicillin-resistant Staphylococcus aureusrdquoAntimicrobial Agents and Chemotherapy vol 48 no 11 pp4322ndash4327 2004
[6] S Hemaiswarya A K Kruthiventi and M Doble ldquoSynergismbetween natural products and antibiotics against infectiousdiseasesrdquo Phytomedicine vol 15 no 8 pp 639ndash652 2008
[7] M Shimizu S Shiota T Mizushima et al ldquoMarked poten-tiation of activity of 120573-lactams against methicillin-resistantStaphylococcus aureus by corilaginrdquo Antimicrobial Agents andChemotherapy vol 45 no 11 pp 3198ndash3201 2001
[8] P I Alade andO N Irobi ldquoAntimicrobial activities of crude leafextracts ofAcalypha wilkesianardquo Journal of Ethnopharmacologyvol 39 no 3 pp 171ndash174 1993
[9] B E Akinde andOO Odeyemi ldquoExtraction andmicrobiologi-cal evaluation of the oils from the leaves ofAcalyphawilkesianardquoNigerian Medical Journal vol 17 pp 163ndash165 1987
[10] S W Lim K N Ting T D Bradshaw et al ldquoAcalyphawilkesiana extracts induce apoptosis by causing single strandand double strand DNA breaksrdquo Journal of Ethnopharmacologyvol 138 no 2 pp 616ndash623 2011
[11] S W Lim H S Loh K N Ting T D Bradshaw and N AZeenathul ldquoAcalypha wilkesiana ethyl acetate extract enhancesthe in vitro cytotoxic effects of 120572-tocopherol in human brainand lung cancer cellsrdquo International Journal of BioscienceBiochemistry and Bioinformatics vol 3 no 4 pp 335ndash340 2013
[12] M Othman S Genapathy P S Liew et al ldquoSearch for antibac-terial agents from Malaysian rainforest and tropical plantsrdquoNatural Product Research vol 25 no 19 pp 1857ndash1864 2011
[13] G Spigno and D M de Faveri ldquoAntioxidants from grape stalksandmarc influence of extraction procedure on yield purity andantioxidant power of the extractsrdquo Journal of Food Engineeringvol 78 no 3 pp 793ndash801 2007
[14] MOthmanH S Loh CWiart T J Khoo KH Lim andKNTing ldquoOptimal methods for evaluating antimicrobial activitiesfrom plant extractsrdquo Journal of Microbiological Methods vol 84no 2 pp 161ndash166 2011
[15] Clinical and Laboratory Standards Institute (CLSI) ldquoLabora-tory methods for detection of methicillin-resistant Staphylococ-cus aureus (MRSA)rdquo Supplement M100-S17 2007
[16] JMAndrews ldquoDetermination ofminimum inhibitory concen-trationsrdquo Journal of Antimicrobial Chemotherapy vol 48 no 1pp 5ndash16 2001
[17] D F J Brown ldquoDetection of methicillinoxacillin resistance instaphylococcirdquo Journal of Antimicrobial Chemotherapy vol 48no 1 pp 65ndash70 2001
[18] P Cos A J Vlietinck D V Berghe and LMaes ldquoAnti-infectivepotential of natural products how to develop a stronger in vitroproof-of-conceptrdquo Journal of Ethnopharmacology vol 106 no3 pp 290ndash302 2006
[19] R L White D S Burgess M Manduru and J A BossoldquoComparison of three different in vitro methods of detectingsynergy time-kill checkerboard and E testrdquo AntimicrobialAgents and Chemotherapy vol 40 no 8 pp 1914ndash1918 1996
[20] US Food andDrug Administration (FDA)CDERNewMolecu-lar Entity (NME) Drug and New Biologic Approvals in CalendarYear 2006
[21] F J Schmitz A C Fluit M Luckefahr et al ldquoThe effect ofreserpine an inhibitor of multidrug efflux pumps on the in-vitro activities of ciprofloxacin sparfloxacin and moxifloxacinagainst clinical isolates of Staphylococcus aureusrdquo Journal ofAntimicrobial Chemotherapy vol 42 no 6 pp 807ndash810 1998
[22] F R Stermitz P Lorenz J N Tawara L A Zenewicz and KLewis ldquoSynergy in a medicinal plant Antimicrobial action ofberberine potentiated by 5rsquo-methoxyhydnocarpin a multidrugpump inhibitorrdquoProceedings of theNational Academy of Sciencesof theUnited States of America vol 97 no 4 pp 1433ndash1437 2000
[23] Y Cho N L Schiller and K Oh ldquoAntibacterial effects of greentea polyphenols on clinical isolates of methicillin-resistantStaphylococcus aureusrdquo Current Microbiology vol 57 no 6 pp542ndash546 2008
[24] S Shiota M Shimizu J Sugiyama Y Morita T Mizushimaand T Tsuchiya ldquoMechanisms of action of corilagin andtellimagrandin I that remarkably potentiate the activity of120573-lactams against methicillin-resistant Staphylococcus aureusrdquoMicrobiology and Immunology vol 48 no 1 pp 67ndash73 2004
[25] S Dukan and T Nystrom ldquoBacterial senescence stasis resultsin increased and differential oxidation of cytoplasmic proteinsleading to developmental induction of the heat shock regulonrdquoGenes amp Development vol 12 no 21 pp 3431ndash3441 1998
[26] R A Venezia B E Domaracki A M Evans K E Preston andE M Graffunder ldquoSelection of high-level oxacillin resistancein heteroresistant Staphylococcus aureus by fluoroquinoloneexposurerdquo Journal of Antimicrobial Chemotherapy vol 48 no3 pp 375ndash381 2001
[27] J M Navarro Llorens A Tormo and E Martınez-Garcıa ldquoSta-tionary phase in gram-negative bacteriardquo FEMS MicrobiologyReviews vol 34 no 4 pp 476ndash495 2010
[28] M D Rolfe C J Rice S Lucchini et al ldquoLag phase is a distinctgrowth phase that prepares bacteria for exponential growth andinvolves transient metal accumulationrdquo Journal of Bacteriologyvol 194 no 3 pp 686ndash701 2011
[29] P Lambert ldquoMechanism of action of antibiotics and synthethicanti-infective agentsrdquo in Hugo and Russells PharmaceuticalMicrobiology S PDenyerNAHodges and S PGorman Edspp 202ndash206 Blackwell Publishing Oxford UK 7th edition2004
[30] A Smith ldquoBacterial resistance to antibioticsrdquo inHugo and Rus-sellrsquos Pharmaceutical Microbiology S P Denyer N A Hodgesand S P Gorman Eds pp 220ndash223 Blackwell Oxford UK7th edition 2004
[31] H F Chambers ldquoMethicillin resistance in staphylococcimolec-ular and biochemical basis and clinical implicationsrdquo ClinicalMicrobiology Reviews vol 10 no 4 pp 781ndash791 1997
[32] G L Archer and JM Bosilevac ldquoSignaling antibiotic resistancein staphylococcirdquo Science vol 291 no 5510 pp 1915ndash1916 2001
[33] H Z Zhang C J Hackbarth K M Chansky and H FChambers ldquoA proteolytic transmembrane signaling pathwayand resistance to 120573-lactams in Staphylococcirdquo Science vol 291no 5510 pp 1962ndash1965 2001
[34] S Deresinski ldquoMethicillin-resistant Staphylococcus aureus anevolutionary epidemiologic and therapeutic odysseyrdquo ClinicalInfectious Diseases vol 40 no 4 pp 562ndash573 2005
[35] W M Din J Chu G Clarke et al ldquoAntioxidant and cytopro-tective effects of an ethanol extract of Acalypha wilkesiana varmacafeana from Malaysiardquo Natural Product Communicationsvol 8 no 3 pp 375ndash380 2013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 7
to homogeneously methicillin-resistant Staphylococcus aureusrdquoAntimicrobial Agents and Chemotherapy vol 48 no 11 pp4322ndash4327 2004
[6] S Hemaiswarya A K Kruthiventi and M Doble ldquoSynergismbetween natural products and antibiotics against infectiousdiseasesrdquo Phytomedicine vol 15 no 8 pp 639ndash652 2008
[7] M Shimizu S Shiota T Mizushima et al ldquoMarked poten-tiation of activity of 120573-lactams against methicillin-resistantStaphylococcus aureus by corilaginrdquo Antimicrobial Agents andChemotherapy vol 45 no 11 pp 3198ndash3201 2001
[8] P I Alade andO N Irobi ldquoAntimicrobial activities of crude leafextracts ofAcalypha wilkesianardquo Journal of Ethnopharmacologyvol 39 no 3 pp 171ndash174 1993
[9] B E Akinde andOO Odeyemi ldquoExtraction andmicrobiologi-cal evaluation of the oils from the leaves ofAcalyphawilkesianardquoNigerian Medical Journal vol 17 pp 163ndash165 1987
[10] S W Lim K N Ting T D Bradshaw et al ldquoAcalyphawilkesiana extracts induce apoptosis by causing single strandand double strand DNA breaksrdquo Journal of Ethnopharmacologyvol 138 no 2 pp 616ndash623 2011
[11] S W Lim H S Loh K N Ting T D Bradshaw and N AZeenathul ldquoAcalypha wilkesiana ethyl acetate extract enhancesthe in vitro cytotoxic effects of 120572-tocopherol in human brainand lung cancer cellsrdquo International Journal of BioscienceBiochemistry and Bioinformatics vol 3 no 4 pp 335ndash340 2013
[12] M Othman S Genapathy P S Liew et al ldquoSearch for antibac-terial agents from Malaysian rainforest and tropical plantsrdquoNatural Product Research vol 25 no 19 pp 1857ndash1864 2011
[13] G Spigno and D M de Faveri ldquoAntioxidants from grape stalksandmarc influence of extraction procedure on yield purity andantioxidant power of the extractsrdquo Journal of Food Engineeringvol 78 no 3 pp 793ndash801 2007
[14] MOthmanH S Loh CWiart T J Khoo KH Lim andKNTing ldquoOptimal methods for evaluating antimicrobial activitiesfrom plant extractsrdquo Journal of Microbiological Methods vol 84no 2 pp 161ndash166 2011
[15] Clinical and Laboratory Standards Institute (CLSI) ldquoLabora-tory methods for detection of methicillin-resistant Staphylococ-cus aureus (MRSA)rdquo Supplement M100-S17 2007
[16] JMAndrews ldquoDetermination ofminimum inhibitory concen-trationsrdquo Journal of Antimicrobial Chemotherapy vol 48 no 1pp 5ndash16 2001
[17] D F J Brown ldquoDetection of methicillinoxacillin resistance instaphylococcirdquo Journal of Antimicrobial Chemotherapy vol 48no 1 pp 65ndash70 2001
[18] P Cos A J Vlietinck D V Berghe and LMaes ldquoAnti-infectivepotential of natural products how to develop a stronger in vitroproof-of-conceptrdquo Journal of Ethnopharmacology vol 106 no3 pp 290ndash302 2006
[19] R L White D S Burgess M Manduru and J A BossoldquoComparison of three different in vitro methods of detectingsynergy time-kill checkerboard and E testrdquo AntimicrobialAgents and Chemotherapy vol 40 no 8 pp 1914ndash1918 1996
[20] US Food andDrug Administration (FDA)CDERNewMolecu-lar Entity (NME) Drug and New Biologic Approvals in CalendarYear 2006
[21] F J Schmitz A C Fluit M Luckefahr et al ldquoThe effect ofreserpine an inhibitor of multidrug efflux pumps on the in-vitro activities of ciprofloxacin sparfloxacin and moxifloxacinagainst clinical isolates of Staphylococcus aureusrdquo Journal ofAntimicrobial Chemotherapy vol 42 no 6 pp 807ndash810 1998
[22] F R Stermitz P Lorenz J N Tawara L A Zenewicz and KLewis ldquoSynergy in a medicinal plant Antimicrobial action ofberberine potentiated by 5rsquo-methoxyhydnocarpin a multidrugpump inhibitorrdquoProceedings of theNational Academy of Sciencesof theUnited States of America vol 97 no 4 pp 1433ndash1437 2000
[23] Y Cho N L Schiller and K Oh ldquoAntibacterial effects of greentea polyphenols on clinical isolates of methicillin-resistantStaphylococcus aureusrdquo Current Microbiology vol 57 no 6 pp542ndash546 2008
[24] S Shiota M Shimizu J Sugiyama Y Morita T Mizushimaand T Tsuchiya ldquoMechanisms of action of corilagin andtellimagrandin I that remarkably potentiate the activity of120573-lactams against methicillin-resistant Staphylococcus aureusrdquoMicrobiology and Immunology vol 48 no 1 pp 67ndash73 2004
[25] S Dukan and T Nystrom ldquoBacterial senescence stasis resultsin increased and differential oxidation of cytoplasmic proteinsleading to developmental induction of the heat shock regulonrdquoGenes amp Development vol 12 no 21 pp 3431ndash3441 1998
[26] R A Venezia B E Domaracki A M Evans K E Preston andE M Graffunder ldquoSelection of high-level oxacillin resistancein heteroresistant Staphylococcus aureus by fluoroquinoloneexposurerdquo Journal of Antimicrobial Chemotherapy vol 48 no3 pp 375ndash381 2001
[27] J M Navarro Llorens A Tormo and E Martınez-Garcıa ldquoSta-tionary phase in gram-negative bacteriardquo FEMS MicrobiologyReviews vol 34 no 4 pp 476ndash495 2010
[28] M D Rolfe C J Rice S Lucchini et al ldquoLag phase is a distinctgrowth phase that prepares bacteria for exponential growth andinvolves transient metal accumulationrdquo Journal of Bacteriologyvol 194 no 3 pp 686ndash701 2011
[29] P Lambert ldquoMechanism of action of antibiotics and synthethicanti-infective agentsrdquo in Hugo and Russells PharmaceuticalMicrobiology S PDenyerNAHodges and S PGorman Edspp 202ndash206 Blackwell Publishing Oxford UK 7th edition2004
[30] A Smith ldquoBacterial resistance to antibioticsrdquo inHugo and Rus-sellrsquos Pharmaceutical Microbiology S P Denyer N A Hodgesand S P Gorman Eds pp 220ndash223 Blackwell Oxford UK7th edition 2004
[31] H F Chambers ldquoMethicillin resistance in staphylococcimolec-ular and biochemical basis and clinical implicationsrdquo ClinicalMicrobiology Reviews vol 10 no 4 pp 781ndash791 1997
[32] G L Archer and JM Bosilevac ldquoSignaling antibiotic resistancein staphylococcirdquo Science vol 291 no 5510 pp 1915ndash1916 2001
[33] H Z Zhang C J Hackbarth K M Chansky and H FChambers ldquoA proteolytic transmembrane signaling pathwayand resistance to 120573-lactams in Staphylococcirdquo Science vol 291no 5510 pp 1962ndash1965 2001
[34] S Deresinski ldquoMethicillin-resistant Staphylococcus aureus anevolutionary epidemiologic and therapeutic odysseyrdquo ClinicalInfectious Diseases vol 40 no 4 pp 562ndash573 2005
[35] W M Din J Chu G Clarke et al ldquoAntioxidant and cytopro-tective effects of an ethanol extract of Acalypha wilkesiana varmacafeana from Malaysiardquo Natural Product Communicationsvol 8 no 3 pp 375ndash380 2013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology