pengoksidan sulfit oleh adenosin-5'-posfosulfat … papers/pert vol. 2 (2) dec. 19… ·...
TRANSCRIPT

Pertanika, 2(2), 118-124 (1979)
Pengoksidan Sulfit oleh Adenosin-5'-PosfosulfatReduktase Dalam Thiobacillus denitrificans
(The oxidation of sulphite by Adenosine-5'-phosphosulphate reductase inThiobacillus denitrificans)
M. AMINUDDINJabatan Biokimia dan Mikrobiologi, Fakulti Sains dan Pengajian Alarn Sekitar,
Universiti Pertanian Malaysia.
Key words: Thiobacillus denitrificans \ APS-reduktase
RINGKASAN
Sulfit dioksidkan dengan adanya AMP oleh satu enzini yang terdapat dalam pecahan-larut sel. Denganadanya ferrisianid, sulfit dan AMP dengan mudahnya membentuk APS di dalam ekstrak kasar. Tetapi,jika ferrisianid digantikan dengan nitrat dan FMN di bawah keadaan anaerobik hasilan APS hanya sebanyak10% daripada yang terdapat dengan ferrisianid. APS reduktase telah ditulinkan 35-kali. Enzim yangtertulin dihalang oleh bahan-bahan uji pengikat-thiol Berat molekulnya dianggarkan sebanyak 250,000.
SUMMARY
Sulphite is oxidised in the presence of AMP by an enzyme present in the soluble fraction of the cellIn the presence of ferricyanide, sulphite and AMP readily formed APS in the crude extract. However, whenferricyanide was replaced by nitrate and FMN under anaerobic conditions the yield of APS was only about10% of that obtained with ferricyanide. APS reductase was purified 35-fold. The purified enzyme wasinhibited by thiol-binding reagents. Its molecular weight was estimated to be 250,000.
PENGENALAN
Thiobacillus denitrificans, seperti Iain-lainthiobacilli (Lyric dan Suzuki, 1970; Moriartydan Nicholas, 1970) adalah satu bakteriumkimiolithotrof sebab ianya mengoksidkan se-batian-sebatian sulfan takorganan untuk men-dapatkan tenaga bagi penumbuhan. Ianya jugasatu anaerob fakultatif, iaitu ia menggunakannitrat, bukan oksigen, sebagai penerima elektrondi bawah keadaan anaerobik.
Di dalam T. denitrificans sulfit telah di-tunjukkan sebagai hasil pengoksidan sulfld dansulfan unsur (Aminuddin dan Nicholas, 1973;Aminuddin, 1979A). Pengoksidan sulfit seterus-nya oleh thiobacilli dijalankan mengikut duaedaran. Mekanisma pertama yang dikemukakanoleh Peck (1960, 1962) melibatkan pembentukanadenosin-5'-posfosulfat (APS) daripada AMPdan sulfit yang dimangkinkan oleh APS-reduk-
tase. APS yang terbentuk kemudian ditukarkepada ATP oleh gabungan tindakan dua enzimiaitu ADP-sulfurilase dan adenilat kinase. Kedua-dua tindakbalas ini juga menghasilkan kembaliAMP.
Sulfit juga boleh dioksidkan tanpa AMP(London dan Rittenberg, 1964; Charles danSuzuki, 1966) dan dari itu melibatkan mekanismayang berlainan. Charles dan Suzuki (1966)telah mengasingkan dan menulinkan suatu sulfitoksidase dari Thiobacillus novellus yang memerlu-kan ferrisianid atau sitokrom c sebagai penerimaelektron. Enzim yang serupa juga telah ditulin-kan dari Thiobacillus thioparus (Lyric dan Suzuki,1970A) dan T. denitrificans (Aminuddin danNicholas, 1974).
Kertas ini membentangkan penulinan APS-reduktase serta sifat-sifatnya dari T. denitrificans.
DEAE = diethilaminoethil; EDTA == ethilenediamin tetrasetat; pCMB = para-kloromerkuribenzoat;HOQNO » 2-heptil-4-hidroksikuionoIin-N-oksid; DIECA - diethildithiokarbamat.
118

M. AMINUDDIN
BAHAN DAN CARA
Penumbuhan dan penuaian organismaT. denitrificans, stren "Oslo", ditumbuhkan
secara anaerobik selama 3 hari pada 30°C dalamkarboy-karboy 40-1. Larutan kultura mengan-dungi (g/1): Na2S2O 3. 5H2O, 5.0; KOH, 0.2;NaHCO3, 1.0; KNO3) 2.0: NH4C1, 0.5; MgSO4 .7H2O, 0.5; KH2PO4, 2.0; FeSO4, 0.01. Apabilaasiditi meningkat ianya diubah kepada pH 7.0dengan 20% (berat/isipadu) K2CO3. 101 ino-kulum dituangkan ke dalam 30 1 medium untukmendapatkan isipadu akhir 40 1.
Sel-sel yang tumbuh dikutip pada 2°Cdalam pengempar sejuk Sorvall RC-2 yang di-pasang dengan alat alir-terus. Sel-sel kemudian-nya dibasuh dua kali dengan 0.05M penampanposfat (pH 7.0).
Penyediaan ekstrakSel-sel yang diampai dengan 0.05M penam-
pan posfat (pH 7.0) dipecahkan melalui SelTekanan French pada 20,000 paun/in persegipada 2°C. Prosidur ini diulangi. Homogenatkasar yang diperolehi kemudiannya digemparkanpada 10,000 xg selama 30 min dan supernatanyang didapati digunakan sebagai punca enzim.
Penetapan aktiviti APS-reduktaseKaedah yang digunakan adalah berasaskan
cara Peck (1961) dengan beberapa perubahan.Ramuan tindakbalas mengandungi (jumole): Tris-HC1 (pH 7.5), 150; Na2SO3, 10; K3Fe(CN)6,5.0; AMP, 5.0 dan enzim dalam isipadu akhirsebanyak 3.0 ml. Tindakbalas yang dijalankandalam kuvet quartz (1 sm) dimula dengan me-masukkan sulfit ke dalam kuvet sampel. Kuvetrujukan mengandungi semua komponen kecualisulfit. Perubahan penyerapan pada 420 nm di-ikuti dengan menggunakan spektrofotometerperekod Unicam SP-800 yang telah diekuili-brasikan pada 30°C. Aktiviti enzim disebutsecara /xmole ferrisianid terturun per jam permg protin.
Pengasingan nukleotid-nukleotidNukleotid-nukleotid diasingkan samada
menggunakan kolom DEAE-selulos atau eiektro-foresis voltan tinggi. Kolom DEAE-selulos(8 X 1.7 sm) dibina dengan DE-11 yang telahdiekuilibrasikan dengan 1M penampan format(pH 6.0). Campuran nukleotid kemudian di-letakkan di atas kolom dan dialir-keluar dengancerunan linear 0.2M ammonium bikarbonat danair suling (kedua isipadu sama). Pecahan-pecahan dikutip pada suhu bilik dengan meng-gunakan pemungut pecahan otomatik.
Untuk pengenalan, nukleotid-nukleotid se-terusnya diasingkan di atas kertas kromatografiWhatman 3MM dalam 0.1M penampan nateriumsitrat (pH 5.0) pada 1500 V selama 14 jam.Radas eletrophoresis Tate (1968) digunakan.AMP, ADP, APS dan ATP digunakan sebagainukleotid piawai. Nukleotid-nukleotid ini di-kesan di atas kertas yang sudah kering denganmengikuti penyerapan mereka terhadap cahayalampau-ungu.
Pengiraan radioaktivitiBahagian-bahagian radioaktif elektrophore-
togram dipotong kecil-kecil (1.5 x 2 sm) dansetiap potongan diletakkan ke dalam vail kaca.Larutan kerlipan yang mengandungi 0.5% (berat/isipadu) 2,5-difeniloksazol (PPO) dan 0.3%(berat/isipadu) l,4-bis-(2-(4-methil-5'-feniloksa-zolil) )-benzen (POPOP) dalam toluen kemudian-nya dimasukkan ke dalam vail dan radioaktivitidikira dengan Spektrometer Kerlipan CecairPackard Tri-Carb (Model 3375).
Radioaktiviti dalam larutan pula dikiradengan mengambil satu alikuot sampel dan di-campurkan dengan campuran kerlipan yangterdiri dari 2,0 ml 95% (berat/isipadu) ethanoldan 5 ml toluen dalam vail kaca. PPO danPOPOP kemudian dimasukkan seperti di atasdan radioaktiviti dikira.
Penetapan protinKaedah Folin yang dihuraikan oleh Lowry
et aL (1951) digunakan dengan albumin serumlembu sebagai protin piawai.
Bahan-bahan kimiaLarutan sulfit disediakan dengan melarutkan
Na2SO3 . 9H2O dalam air suling-berganda yangmengandungi 0.2 mM Na-EDTA. Protinpenanda untuk penurasan gel dibeli dari MannResearch Laboratories, New York, U.S.A.Sitokrom c, albumin serum lembu, nukleotid-nukleotid, pCMB dan dithiothreitol didapati dariSigma Chemical Co. Missouri, U.S.A. Nateriumsulfit radioaktif (35S-label) dibeli dari Radio-chemical Centre, Amersham, U.K. WhatmanDEAE-selulos dan kertas kromatografi Whatman3MM dibekalkan oleh Reeve Angel and Co. Ltd.London. Sephadex G-200 dan dekstran birudibeli dari Pharmacia, Uppsala, Sweden. PPOdan POPOP diperolehi dari Packard InstrumentCo. Chicago, U.S.A. Mutu bahan-bahan kimiayang lain adalah yang paling tinggi yang bolehdidapati.
119
A.P.S. REDUKTASE DALAM TH/OBACILLUS DENITRIFICANS
HASIL-HASIL
Pembentukan APSEkstrak kasar (S10) telah diperamkan dengan
35S-sulfit dan AMP serta beberapa penerimaelektron. Hasil tindakbalas ini diasingkan dengankromatografi kolom DEAE-selulos. Nukleotid-nukleotid yang mengalir-keluar seterusnya di-asing dan ditetapkan dengan elektrophoresisvoltan tinggi.
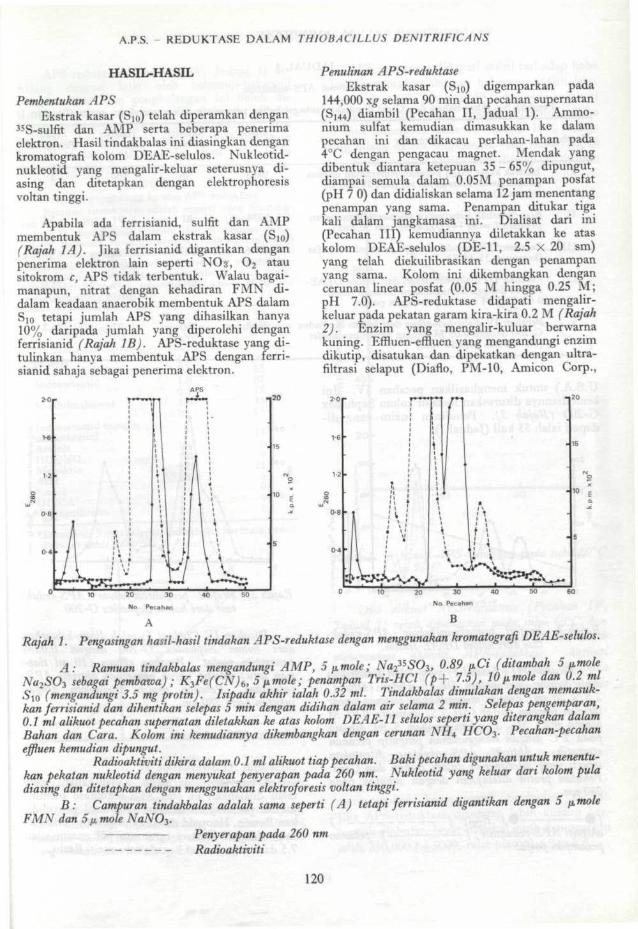
Apabila ada ferrisianid, sulfit dan AMPmembentuk APS dalam ekstrak kasar (SI0)(Rajah 1A). Jika ferrisianid digantikan denganpenerima elektron lain seperti NOj, O2 atausitokrom cy APS tidak terbentuk. Walau bagai-manapun, nitrat dengan kehadiran FMN di-dalam keadaan anaerobik membentuk APS dalamS lo tetapi jumlah APS yang dihasilkan hanya10% daripada jumlah yang diperolehi denganferrisianid (Rajah IB). APS-reduktase yang di-tulinkan hanya membentuk APS dengan ferri-sianid sahaja sebagai penerima elektron.
20
10 20 30
No Pecahan
Penulinan APS-reduktaseEkstrak kasar (S10) digemparkan pada
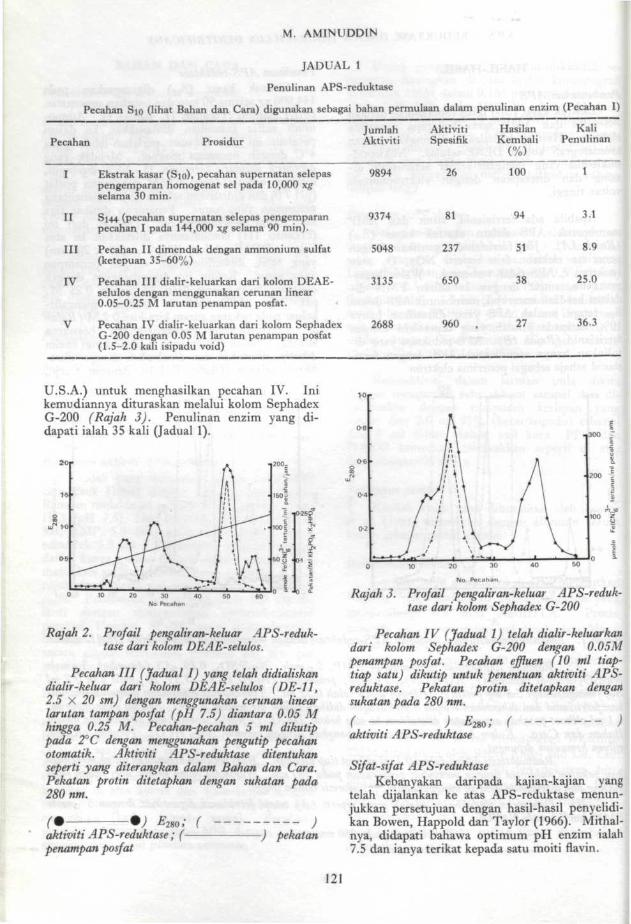
144,000 xg selama 90 min dan pecahan supernatan(S144) diambil (Pecahan II, Jadual 1). Ammo-nium sulfat kemudian dimasukkan ke dalampecahan ini dan dikacau perlahan-lahan pada4°C dengan pengacau magnet. Mendak yangdibentuk diantara ketopuan 35 - 65% dipungut,diampai semula dalam 0.05M penampan posfat(pH 7 0) dan didialiskan selama 12 jam menentangpenampan yang sama. Penampan ditukar tigakali dalam jangkamasa ini. Dialisat dari ini(Pecahan III) kemudiannya diletakkan ke ataskolom DEAE-selulos (DE-11, 2.5 x 20 sm)yang telah diekuilibrasikan dengan penampanyang sama. Kolom ini dikembangkan dengancerunan linear posfat (0.05 M hingga 0.25 M;pH 7.0). APS-reduktase didapati mengalir-keluar pada pekatan garam kira-kira 0.2 M (Rajah2). Enzim yang mengalir-kuluar berwarnakuning. Effluen-effluen yang mengandungi enzimdikutip, disatukan dan dipekatkan dengan ultra-filtrasi selaput (Diaflo, PM-10, Amicon Corp.,
10 *
30
No Pecahan
Rajah L Pengasingan hasil-hasil tindakan APS-reduktase dengan menggunakan kromatografi DEAE-selulos.
A: Ramuan tindakbalas mengandungi AMP, 5 fimole; Na2^SO3t 0.89 pCi (ditambah 5 pNa2SO3 sebagai pembawa) ; K3Fe(CN)6, 5 pmole; penampan Tris-HCl (p+ 7.5), 10pmole dan UJ mlS10 (mengandungi 3,5 mg protin). Isipadu akhir ialah 0,32 ml Tindakbalas dimulakan dengan memasuk-kan ferrisianid dan dihentikan selepas 5 min dengan didihan dalam air selama 2 min. Selepas pengemparan,0.1 ml alikuot pecahan supernatan diletakkan ke atas kolom DEAE-11 selulos seperti yang diterangkan dalamBahan dan Cara. Kolom ini kemudiannya dikembangkan dengan cerunan NHA HCO3. Pecahan-pecahaneffluen kemudian dipungut.
Radioaktivitidikira dalam 0.1 ml alikuot tiap pecahan. Baki pecahan digunakan untukmenentu-kan pekatan nukleotid dengan menyukat penyerapan pada 260 nm. Nukleotid yang keluar dan kolom puladiasing dan ditetapkan dengan menggunakan elektroforesis voltan tinggi.
B: Campuran tindakbalas adalah sama seperti (A) tetapi ferrisianid digantikan dengan 5FMN dan 5(x mole NaNO3.
Penyerapan pada 260 nmRadioaktiviti
120
M. AMINUDDIN
JADUAL 1
Penulinan APS-reduktase
Pecahan Sio (Hhat Bahan dan Cara) digunakan sebagai bahan permulaan dalam penulinan enzim (Pecahan I)
Pecahan Prosidur
I Ekstrak kasar (Sio), pecahan supernatan selepaspengemparan homogenat sel pada 10,000 xgselama 30 min.
II S144 (pecahan supernatan selepas pengemparanpecahan I pada 144,000 xg selama 90 min).
III Pecahan II dimendak dengan ammonium sulfat(ketepuan 35-60%)
IV Pecahan III dialir-keluarkan dari kolom DEAE-selulos dengan menggunakan cerunan linear0.05-0.25 M larutan penampan posfat.
V Pecahan IV dialir-keluarkan dari kolom SephadexG-200 dengan 0.05 M larutan penampan posfat(1.5-2.0 kali isipadu void)
JumlahAktiviti
9894
9374
5048
3135
AktivitiSpesifik
26
81
237
650
HasilanKembali
100
94
51
38
KaliPenulinan
1
3.1
8.9
25.0
2688 960 27 36.3
U.S.A.) untuk menghasilkan pecahan IV. Inikemudiannya dituraskan melalui kolom SephadexG-200 (Rajah 3). Penulinan enzim yang di-dapati ialah 35 kali (Jadual 1).
Rajah 2. Prof ail pengaliran-keluar APS-reduk-tase dari kolom DEAE-selulos.
Pecahan HI (Jadual I) yang telah didialiskandialir-keluar dari kolom DEAE-selulos (DE-11,2.5 X 20 sm) dengan menggunakan cerunan linearlarutan tampan posfat (pH 7.5) diantara 0.05 Mhingga 0.25 M. Pecahan-pecahan 5 ml dikutippada 2°C dengan menggunakan pengutip pecahanotomatik. Aktiviti APS-reduktase ditentukanseperti yang diterangkan dalam Bahan dan Cara.Pekatan protin ditetapkan dengan sukatan pada280 nm.
aktiviti APS-reduktase ; (-penampan posfat
i
-) pekatan
20 30
No Pecahan
Rajah J. Profail pengaliran-keluar APS-reduk-tase dari kolom Sephadex G-200
Pecahan IV (Jadual 1) telah dialir-keluarkandari kolom Sephadex G-200 dengan 0.05Mpenampan posfat. Pecahan effluen (10 ml tiap'tiap satu) dikutip untuk penentuan aktiviti APS-reduktase. Pekatan protin ditetapkan dengansukatan pada 280 nm.( ) £2g0; ( )aktiviti APS-reduktase
Sifat-sifat APS-reduktaseKebanyakan daripada kajian-kajian yang
telah dijalankan ke atas APS-reduktase menun-jukkan persetujuan dengan hasil-hasil penyelidi-kan Bowen, Happold dan Taylor (1966). Mithal-nya, didapati bahawa optimum pH enzim ialah7.5 dan ianya terikat kepada satu moiti flavin.
121
A.P.S. REDUKTASE DALAM I'HIOBACILLUS DENITRIFICANS
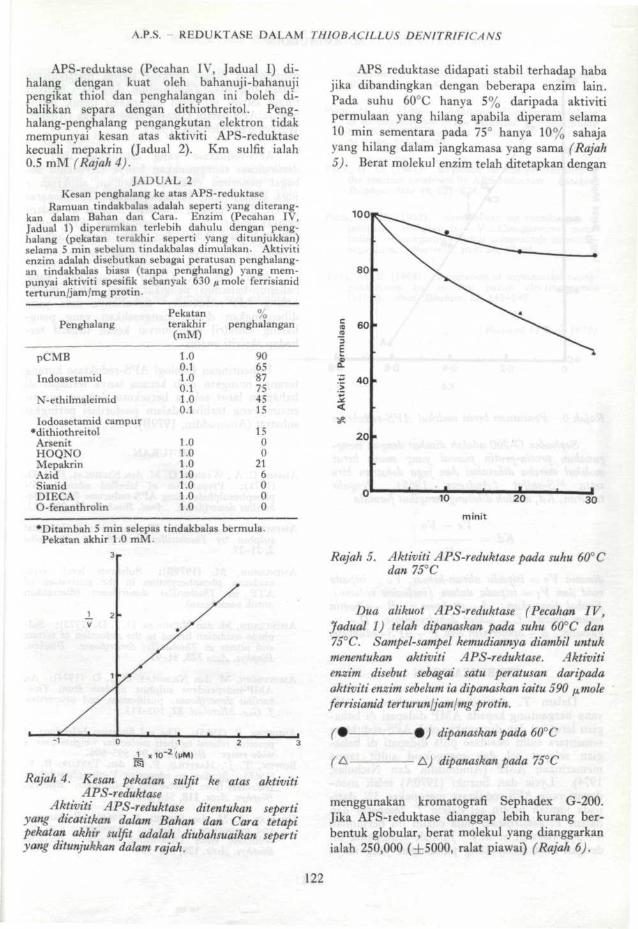
APS-reduktase (Pecahan IV, Jadual I) di-halang dengan kuat oleh bahanuji-bahanujipengikat thiol dan penghalangan ini boleh di-balikkan separa dengan dithiothreitol. Peng-halang-penghalang pengangkutan elektron tidakmempunyai kesan atas aktiviti APS-reduktasekecuali mepakrin (Jadual 2). Km sulfit ialah0.5 mM (Rajah 4).
JADUAL 2Kesan penghalang ke atas APS-reduktase
Ramuan tindakbalas adalah seperti yang diterang-kan dalam Bahan dan Cara. Enzim (Pecahan IV,Jadual 1) diperamkan terlebih dahulu dengan peng-halang (pekatan terakhir seperti yang ditunjukkan)selama 5 min sebelum tindakbalas dimulakan. Aktivitienzim adalah disebutkan sebagai peratusan penghalang-an tindakbalas biasa (tanpa penghalang) yang mem-punyai aktiviti spesifik sebanyak 630 ^ mole ferrisianidterturun/jam/mg protin.
PenghalangPekatanterakhir(mM)
penghalangan
pCMB
Indoasetamid
N-ethilmaleimid
Iodoasetamid campur•dithiothreitolArsenitHOQNOMepakrinAzidSianidDIECAO-fenanthrolin
1.00.11.00.11.00.1
1.01.01.01.01.01.01.0
906587754515
1500
216000
•Ditambah 5 min selepas tindakbalas bermula.Pekatan akhir 1.0 mM.
1 2V
1 x 10"IS1
(MM)
Rajah 4. Kesan pekatan sulfit ke atas aktivitiAPS-reduktase
Aktiviti APS-reduktase ditentukan sepertiyang dicatitkan dalam Bahan dan Cara tetapipekatan akhir sulfit adalah diubahsuaikan sepertiyang ditunjukkan dalam rajah.
APS reduktase didapati stabil terhadap habajika dibandingkan dengan beberapa enzim lain.Pada suhu 60°C hanya 5% daripada aktivitipermulaan yang hilang apabila diperam selama10 min sementara pada 75° hanya 10% sahajayang hilang dalam jangkamasa yang sama (Rajah5). Berat molekul enzim telah ditetapkan dengan
30
Rajah 5. Aktiviti APS-reduktase pada suhu 60° Cdan 75°C
Dua alikuot APS-reduktase (Pecahan IV,Jadual 1) telah dipanaskan pada suhu 60°C dan75°C. Sampel-sampel kemudiannya diambil untukmenentukan aktiviti APS-reduktase. Aktivitienzim disebut sebagai satu peratusan daripadaaktiviti enzim sebelum ia dipanaskan iaitu 590 pjnoleferrisianid terturun\jam\mg protin.
(A-
%) dipanaskan pada 60° C
• A) dipanaskan pada 75°C
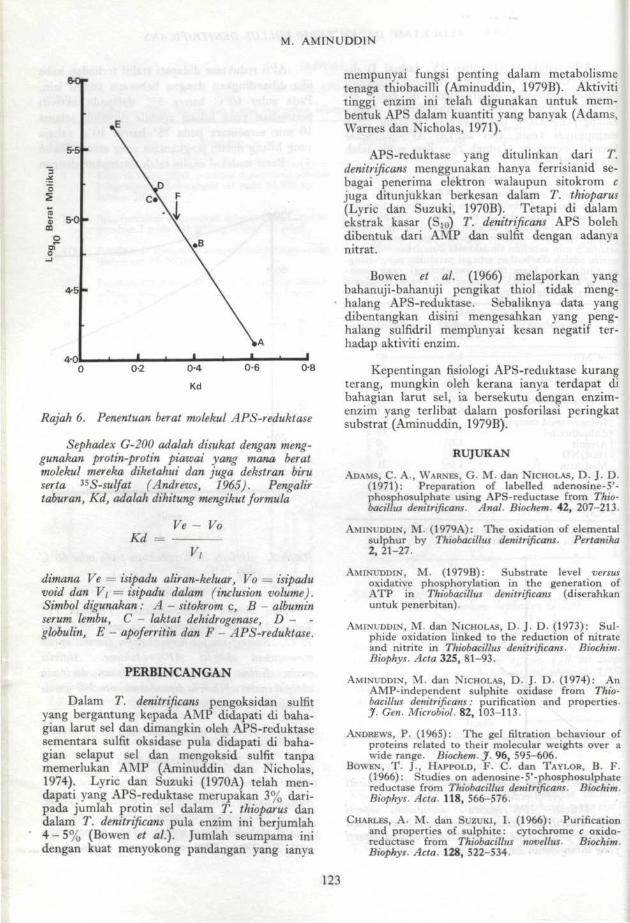
menggunakan kromatografi Sephadex G-200.Jika APS-reduktase dianggap lebih kurang ber-bentuk globular, berat molekul yang dianggarkanialah 250,000 (±5000, ralat piawai) (Rajah 6).
122
M. AMINUDD1N
o2
e-o
5-5 -
5-0 •
4>5
4-00-2 0-4
Kd
0-6 0-8
Rajah 6. Penentuan berat molekul APS-reduktase
Sephadex G-200 adalah disukat dengan meng-gunakan protin-protin piawai yang mana beratmolekul mereka diketahui dan juga dekstran biruserta 3SS-sulfat (Andrews, 1965). Pengalirtaburan, Kd, adalah dihitung mengikut formula
Kd =Ve - Vo
ditnana Ve = isipadu aliran-keluar, Vo = isipaduvoid dan V\ = isipadu dalam (inclusion volume).Simbol digunakan: A - sitokrom c, B - albuminserum lembu, C - laktat dehidrogenase, D - -globulin, E - apoferritin dan F - APS-reduktase.
PERBINCANGAN
Dalam T. denitriftcans pengoksidan sulfityang bergantung kepada AMP didapati di baha-gian larut sel dan dimangkin oleh APS-reduktasesementara sulfit oksidase pula didapati di baha-gian selaput sel dan mengoksid sulfit tanpamemerlukan AMP (Aminuddin dan Nicholas,1974). Lyric dan Suzuki (1970A) telah men-dapati yang APS-reduktase merupakan 3% dari-pada jumlah protin sel dalam T. thioparus dandalam T. denitrificans pula enzim ini berjumlah4 - 5 % (Bowen et al). Jumlah seumpama inidengan kuat menyokong pandangan yang ianya
mempunyai fungsi penting dalam metabolismetenaga thiobacilli (Aminuddin, 1979B). Aktivititinggi enzim ini telah digunakan untuk mem-bentuk APS dalam kuantiti yang banyak (Adams,Warnes dan Nicholas, 1971).
APS-reduktase yang ditulinkan dari T.denitrificans menggunakan hanya ferrisianid se-bagai penerima elektron walaupun sitokrom cjuga ditunjukkan berkesan dalam T. thioparus(Lyric dan Suzuki, 1970B). Tetapi di dalamekstrak kasar (S10) T. denitrificans APS bolehdibentuk dari AMP dan sulfit dengan adanyanit rat.
Bowen et al. (1966) melaporkan yangbahanuji-bahanuji pengikat thiol tidak meng-halang APS-reduktase. Sebaliknya data yangdibentangkan disini mengesahkan yang peng-halang sulfidril mempunyai kesan negatif ter-hadap aktiviti enzim.
Kepentingan fisiologi APS-reduktase kurangterang, mungkin oleh kerana ianya terdapat dibahagian larut sel, ia bersekutu dengan enzim-enzim yang terlibat dalam posforilasi peringkatsubstrat (Aminuddin, 1979B).
RUJUKAN
ADAMS, C. A.t WARNES, G. M. dan NICHOLAS, D. J. D.(1971): Preparation of labelled adenosine-5'-phosphosulphate using APS-reductase from Thio-bacillus denitrificans. Anal. Biochem. 42, 207-213.
AMINUDDIN, M. (1979A): The oxidation of elementalsulphur by Thiobacillus denitrificans, Pertanika2, 21-27.
AMINUDDIN, M. (1979B): Substrate level versusoxidative phosphorylation in the generation ofATP in Thiobacillus denitrificans (diserahkanuntuk penerbitan).
AMINUDDIN, M. dan NICHOLAS, D. J. D. (1973): Sul-phide oxidation linked to the reduction of nitrateand nitrite in Thiobacillus denitrificans. Biochim.Biophys. Acta 325, 81-93.
AMINUDDIN, M- dan NICHOLAS, D. J. D. (1974): AnAMP-independent sulphite oxidase from Thio-bacillus denitrificans: purification and properties.J. Gen. MicrobioL 82, 103-113.
ANDREWS, P. (1965): The gel filtration behaviour ofproteins related to their molecular weights over awide range. Biochem. J. 96, 595-606.
BOWEN, T. J., HAPPOLD, F. C. dan TAYLOR, B. F.(1966): Studies on adenosine-5'-phosphosulphatereductase from Thiobacillus denitrificans. Biochim.Biophys. Acta. 118, 566-576.
CHARLES, A. M. dan SUZUKI, I. (1966): Purificationand properties of sulphite: cytochrome c oxido-reductase from Thiobacillus novellus. Biochim.Biophys. Acta, 128, 522-534.
123

A.P.S. REDUKTASE DALAM THIOBACILLUS DENITRIFICANS
LONDON, J. dan RITTENBERG, S. C. (1964): Path ofsulphur and thiosulphate oxidation by Thiobacilli.Proc. NatL Acad. Sci. U.S. 52, 1183-1190.
LOWRY, O. H., ROSEBROUGH, N. C , FARR, A. L. danRANDALL, R. J. (1951): Protein measurementwith the Folin phenol reagent. J. Biol. Chem.193, 265-275.
LYRIC, R. M. dan SUZUKI, 1. (1970A): Enzymesinvolved in the metabolism of thiosulphate byThiobacilhis thioparus. I. Survey of enzymes andproperties of sulphite: cytochrome c oxidoreduc-tase. Can.J. Biochem. 48, 334-343.
LYRIC, R. M. dan SUZUKI, I. (1970B): Enzymesinvolved in the metabolism of thiosulphate byThiobacillus thioparus. 11. Properties of APS-reductase. Can.J. Biochem. 48, 344-354.
MORIARTY, D. J. W. dan NICHOLAS, D. J. D. (1970):Electron transfer during sulphide and sulphide
oxidation by Thiobacillus concretivorus.Biophys. Ada. 216, 130-138.
Biochmi.
PECK, H. D. (1960): Adenosine- 5' -phosphosulphateas an intermediate in the oxidation of thiosulphateby Thiobacillus thioparus. Proc. NatL Acad. Sci.U.S. 46, 1053-1057.
PECK, H. D. (1961): Evidence for the reversibility ofthe reaction catalysed by APS-reductase. Biochim.Biophys. Ada 49, 621-624.
PECK, H. D. (1962): Symposium on metabolism ofinorganic compounds. V. Comparative meta-bolism of inorganic sulphur compounds in micro-organisms. Bacteriol. Rev. 26, 67-94.
TATE, M. E. (1968): Separation of myoinositol penta-phosphates by moving paper electrophoresis(MPE). Anal. Biochem. 23, 141-149.
(Received 11 June 1972)
124