keragaman morfologi dan genetik serta derajat … · dan tekanan lingkungan tumbuh juga...
TRANSCRIPT

KERAGAMAN MORFOLOGI DAN GENETIK
SERTA DERAJAT KOMPETISI BEBERAPA AKSESI GULMA Echinochloa crus-galli (L.) Beauv. TERHADAP TANAMAN PADI SAWAH
DWI GUNTORO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2012

PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa segala pernyataan dalam disertasi
yang berjudul:
KERAGAMAN MORFOLOGI DAN GENETIK SERTA DERAJAT KOMPETISI BEBERAPA AKSESI GULMA
Echinochloa crus-galli (L.) Beauv. TERHADAP TANAMAN PADI SAWAH
adalah karya saya sendiri dengan arahan komisi pembimbing, bukan hasil jiplakan
atau tiruan serta belum pernah diajukan dalam bentuk apapun untuk memperoleh
gelar program sejenis di perguruan tinggi mana pun. Sumber informasi yang
berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari
penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di
bagian akhir disertasi ini.
Bogor, Januari 2012
Dwi Guntoro
NIM A361050091

ABSTRACT
DWI GUNTORO. Morphological and Genetic Diversity and Degree of Competition of Several Echinochloa crus-galli (L.) Beauv. Accessions to Rice. (Supervised by M. AHMAD CHOZIN, EDI SANTOSA, SOEKISMAN TJITROSEMITO, and ABDUL HARRIS BURHAN).
Weed disturbance is an important issue in rice production in Indonesia.
Echinochloa crus-galli (L.) Beauv. is the most dominant weed in rice leading to loss of production. The research aims to analyze the morphology and genetic diversity of weeds accession E. crus-galli from West Java and degree of competition of E. crus-galli to rice.
Research was carried out by using several steps, i.e : (1) Preliminary experiments on growth and production of rice with E. crus-galli weed population levels originating from three locations, (2) Morphological and genetic evaluation, (3) Identification of potential weed allelophaty E. crus-galli accession, (4) Study effect of E. crus-galli accession on the growth and rice production (greenhouse experiments), (5) Effects of accession and population rate of E. crus-galli on the growth and rice production (field trial), (6) Estimation of degree competition of E. crus-galli by the replacement series method, (7) Study of physiology competition between rice with weeds.
Results showed that E. crus-galli accession from West Java exhibited morphological and genetic diversity. Accession from different geographical locations showed differences in morphology. The diversity of morphology is presumably due to the phenotypic plasticity and adaptation ability of E. crus-galli accessions. Cluster analysis based on SSR molecular markers produced four sub-groups with similarity coefficient of 0.86, with most of the group clustered in geographic zones of western West Java and northen. This genetic diversity might be caused by the movement of genetic material through harvest or through irrigation, isolation distance, and the possibility of mutation.
E. crus-galli accessions had the potential allelophaty based on inhibit of radicle and plumula growth of rice. Based on the potential allelophaty, the accessions clustered to six groups at 0.72 similarity coefficient. Each accession of E. crus-galli showed a potential difference in reducing the growth and yield of rice plants. This ability is probablt related to morphological characteristics and allelophatyic potential. Accession K6 from Karawang showed the highest ability to suppress the growth and production of rice plants.
E. crus-galli weed had a higher degree of competition to rice plants when the weed population was higher than the population of rice plants. Based on the value of aggressiveness, E. crus-galli competed stronger than rice when the population of rice plants and weeds is equal or weed population was higher than that of rice population. Each rice variety had a different response to population levels of E. crus-galli. The competition between rice plants with a E. crus-galli inhibited the physiological process of rice plant.
Key words : Echinochloa crus-galli, morphology, genetic, diversity, allelophaty, competition, replacement series, physiology, rice variety.

RINGKASAN
DWI GUNTORO. Keragaman Morfologi dan Genetik serta Derajat Kompetisi Beberapa Aksesi Gulma Echinochloa crus-galli (L.) Beauv. terhadap Tanaman Padi Sawah. (Dibimbing oleh M. AHMAD CHOZIN, EDI SANTOSA, SOEKISMAN TJITROSEMITO, dan ABDUL HARRIS BURHAN).
Salah satu masalah penting dalam upaya peningkatan produksi padi di Indonesia adalah menurunkan gangguan gulma selama pertumbuhan. Echinochloa crus-galli (L.) Beauv. merupakan gulma dominan pada tanaman padi sawah yang menyebabkan kehilangan hasil produksi. Penurunan produksi tersebut disebabkan oleh adanya kompetisi antara gulma dan tanaman padi terhadap sumberdaya yang tersedia.
Penelitian bertujuan untuk : (1) menganalisis keragaman morfologi dan genetik aksesi gulma E. crus-galli asal Jawa Barat, (2) Mengidentifikasi potensi alelopati aksesi gulma E. crus-galli asal Jawa Barat, (3) Mempelajari pengaruh aksesi dan tingkat populasi E. crus-galli terhadap pertumbuhan dan produksi padi sawah, (4) Mengkaji derajat kompetisi gulma E. crus-galli dengan metode replacement series, dan (5) Mempelajari fisiologi kompetisi antara padi dengan gulma E. crus-galli. Penelitian dilakukan dengan beberapa tahapan antara lain : (1) Percobaan pendahuluan tentang Pertumbuhan dan Produksi Padi pada berbagai Tingkat Populasi Gulma E. crus-galli yang Berasal dari Tiga Lokasi, (2) Keragaman Morfologi dan Genetik Aksesi E. crus-galli asal Jawa Barat, (3) Identifikasi Potensi Alelopati Aksesi Gulma E. crus-galli Asal Jawa Barat, (4) Kajian Pengaruh Aksesi Gulma E. crus-galli terhadap Pertumbuhan dan Produksi Padi (Percobaan Rumah Kaca), (5) Kajian Pengaruh Aksesi dan Tingkat Populasi Gulma E. crus-galli terhadap Pertumbuhan dan Produksi Padi (Percobaan Lapangan), (6) Pendugaan Derajat Kompetisi Gulma E. crus-galli dengan Metode Replacement Series, (7) Kajian Fisiologi Kompetisi antara Padi dengan Gulma.
Hasil penelitian menunjukkan adanya keragaman morfologi dan genetik aksesi gulma E. crus-galli asal Jawa Barat. Aksesi dari lokasi geografis yang berbeda menunjukkan perbedaan morfologi. Keragaman morfologi aksesi selain disebabkan oleh genetik juga disebabkan oleh perbedaan lingkungan tumbuh. Keragaman morfologi pada lingkungan tumbuh yang berbeda disebabkan adanya perbedaan plastisitas fenotipik dan kemampuan adaptasi aksesi gulma E. crus-galli. Praktek budidaya tanaman padi yang intensif seperti di wilayah Karawang dan tekanan lingkungan tumbuh juga mempengaruhi keragaman morfologi aksesi gulma E. crus-galli.
Analisis cluster berdasarkan karakter morfologi dari habitat asal menghasilkan 5 grup pada koefisien kemiripan sebesar 0.78, sedangkan berdasarkan karakter morfologi di rumah kaca (250 m dpl) membentuk 5 grup pada koefisien kemiripan 0.64 dengan anggota grup aksesi yang berbeda.
Analisis cluster berdasarkan penanda molekuler SSR menghasilkan 4 sub grup dengan koefisien kemiripan 0.86, dengan sebagian besar grup mengelompok pada zona geografi Jawa Barat bagian barat dan bagian utara (pantai utara Jawa) dengan pusat keragaman adalah Subang, Karawang dan Pangalengan. Keragaman

genetik ini dapat disebabkan oleh perpindahan material genetik melalui hasil panen atau melalui irigasi, isolasi jarak, dan kemungkinan adanya mutasi.
Aksesi gulma E. crus-galli asal Jawa Barat memiliki potensi alelopati berdasarkan penghambatan plumula dan radikula kecambah padi. Senyawa alelopati potensial yang teridentifikasi di dalam ekstrak akar aksesi E. crus-galli diantaranya adalah golongan senyawa phenolic, pthalic acid, decanoid acid, propanoid, quinon, dan sterol.
Berdasarkan potensi alelopatinya, analisis cluster menghasilkan enam kelompok aksesi pada koefisien kemiripan 0.72 yaitu aksesi dengan inhibition rate plumula rendah dan inhibition rate radikula rendah, inhibition rate plumula tinggi dan inhibition rate radikula sedang, inhibition rate plumula rendah dan inhibition rate radikula sedang, inhibition rate plumula sedang dan inhibition rate radikula rendah, inhibition rate plumula tinggi dan inhibition rate radikula rendah, dan inhibition rate plumula tinggi dan inhibition rate radikula tinggi.
Setiap aksesi gulma E. crus-galli menunjukkan perbedaan potensi dalam menurunkan pertumbuhan dan hasil tanaman padi. Kemampuan tersebut terkait dengan sifat morfologi dan potensi alelopati. Aksesi K6 asal Karawang menunjukkan kemampuan tertinggi dalam menurunkan pertumbuhan dan produksi tanaman padi. Kemampuan ini terkait dengan karakter morfologi aksesi K6 yaitu panjang daun panjang, lebar daun sempit, sudut daun kecil, dan umur berbunga lambat, serta memiliki IR plumula tinggi dan IR radikula sedang.
Gulma E. crus-galli memiliki derajat kompetisi yang lebih besar dibandingkan dengan tanaman padi ketika populasi gulma lebih tinggi daripada populasi tanaman padi yang ditunjukkan dengan penguasaan sarana tumbuh lebih besar. Berdasarkan nilai agresivitas, gulma E. crus-galli lebih kuat berkompetisi dibandingkan tanaman padi ketika populasi padi dan gulma seimbang ataupun populasi gulma lebih tinggi daripada populasi tanaman padi. Derajat kompetisi gulma ditentukan oleh tingkat populasi gulma E. crus-galli di lapangan. Semakin tinggi populasi gulma E. crus-galli, maka pertumbuhan dan produksi tanaman padi semakin menurun. Produksi padi per hektar menurun sebesar 15.33% pada populasi 4 gulma E. crus-galli/m2 hingga 61.50% pada populasi 12 gulma E. crus-galli/m2
Setiap varietas padi memiliki respon yang berbeda terhadap tingkat populasi gulma E. crus-galli. Varietas Fatmawati menunjukkan varietas yang lebih kompetitif dibandingkan dengan varietas Ciherang. Kompetisi antara tanaman padi dengan gulma E. crus-galli menghambat proses fisiologi tanaman padi yang ditunjukkan dengan penurunan peubah proses fisiologi seperti Indeks Luas Daun (ILD), Net Assimilation Ratio (NAR), Relative Growth Rate (RGR), Crop Growth Rate (CGR), dan peningkatan Leaf Area Ratio (LAR).
.
Kata kunci : Echinochloa crus-galli, morfologi, genetik, keragaman, alelopati, kompetisi,
replacement series, fisiologi, varietas padi.


©Hak Cipta Milik Institut Pertanian Bogor, Tahun 2012 Hak Cipta Dilindungi Undang-Undang
1. Dilarang mengutip, sebagian atau seluruhnya dari karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan
karya ilmiah, penyususnan laporan, penulisan kritik atau tinjauan suatu
masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
tulis dalam bentuk apapun tanpa izin IPB


KERAGAMAN MORFOLOGI DAN GENETIK
SERTA DERAJAT KOMPETISI BEBERAPA AKSESI
GULMA Echinochloa crus-galli (L.) Beauv.
TERHADAP TANAMAN PADI SAWAH
DWI GUNTORO
Disertasi Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2012

Judul Disertasi : Keragaman Morfologi dan Genetik serta Derajat Kompetisi
Beberapa Aksesi Gulma Echinochloa crus-galli (L.) Beauv.
terhadap Tanaman Padi Sawah
Nama : Dwi Guntoro
NIM : A361050091
Disetujui
Komisi Pembimbing
Ketua Prof. Dr. Ir M. Ahmad Chozin, MAgr.
Dr. Edi Santosa, SP., MSi Anggota Anggota
Dr. Soekisman Tjitrosemito
Anggota Dr. Ir. Abdul Harris Burhan, MSc
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana Prof. Dr. Ir. Munif Ghulamahdi, MS Dr. Ir. Dahrul Syah, MSc.Agr Tanggal Ujian: Tanggal Lulus:

PRAKATA
Puji dan Syukur penulis panjatkan kepada Allah SWT atas segala karunia
dan petunjuk-Nya sehingga disertasi yang berjudul “Keragaman Morfologi dan
Genetik serta Derajat Kompetisi Beberapa Aksesi Gulma Echinochloa crus-galli
terhadap Tanaman Padi Sawah” dapat diselesaikan.
Penghargaan dan ungkapan terima kasih yang sebesar-besarnya penulis
sampaikan kepada ketua komisi pembimbing Prof. Dr. Ir. M. Ahmad Chozin,
MAgr yang telah memberikan bimbingan, arahan dan masukan selama penelitian
dan penulisan disertasi. Ungkapan penghargaan dan terima kasih juga penulis
sampaikan kepada anggota komisi pembimbing : Dr. Soekisman Tjitrosemito,
Dr. Ir. Abdul Harris Burhan, MSc, dan Dr. Edi Santosa, SP, MSi yang telah
memberikan bimbingan dan masukan dalam penyelesaian disertasi.
Penghargaan dan terima kasih juga penulis sampaikan kepada berbagai
pihak yang telah banyak membantu sehingga disertasi dapat diselesaikan :
1. Kementerian Pendidikan Nasional cq. Direktorat Pendidkan Tinggi atas
beasiswa BPPS yang penulis terima pada tahun 2005-2008.
2. Rektor Institut Pertanian Bogor, Dekan Fakultas Pertanian dan Ketua
Departemen Agronomi dan Hortikultura atas pemberian izin untuk
melanjutkan studi program Doktor.
3. Prof. Dr. Ir. Munif Ghulamahdi, MS, Dr. Ir. Trikoesoeaningtyas, MSc, dan
Dr. Ir. Maya Melati selaku Pimpinan Program Studi Agronomi, Sekolah
Pascasarjana IPB atas masukan dan saran yang diberikan.
4. Dr. Ir. Sri Sudarmiyati Tjitrosoedirdjo dan Dr. Ir. Ahmad Junaedi, Msi
selaku penguji pada ujian prakualifikasi doktor atas masukan dan saran yang
diberikan.
5. Dr. Ir. Sugiyanta, MSi dan Dr. Ir. Ahmad Junaedi, MSi selaku dosen
penguji luar komisi pada ujian tertutup atas masukan-masukan yang
diberikan.
6. Prof. Dr. Ir. A. Karim Makarim, MSc dan Dr. Ir. Sudradjat, MS selaku
penguji luar komisi pada ujian terbuka atas saran dan masukan yang
diberikan.

7. Seluruh Staf Pengajar Departemen Agronomi dan Hortikultura atas
kerjasama, bantuan, dukungan selama penyelesaian studi.
8. Rosalia Frauke, SP., Ikhsan Suhud, SP., Lidya Sofianty, SP., Dwi Arie
Novianti, SP., Verdha Sahrilla Sandhi, SP., Sudianto Samosir, SP, dan
Rusmato, SP atas semua bantuan dan kerjasamanya.
9. Tim Laboratorium Ekotoksikologi, Limbah & Agen Hayati, Departemen
Agronomi dan Hortikultura IPB (Dita Nurul Latifah, Anif Lailatusifa,
Ekasari, Yeni Fitria, dkk.) atas dukungan dan kerjasamanya selamanya
penyelesaian disertasi.
10. Teknisi dan laboran (Mbak Lasih dkk.) atas bantuan yang diberikan selama
penelitian berlangsung.
11. Keluarga tercinta atas semua dukungan, kasih sayang yang tulus, kesabaran,
pengertian dan doa yang diberikan.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2012
Dwi Guntoro

RIWAYAT HIDUP
Penulis dilahirkan di Pekalongan pada tanggal 29 Agustus 1970 sebagai
anak keempat dari pasangan Kartono (alm) dan Casmini (alm). Pendidikan
sarjana ditempuh di Jurusan Budi Daya Pertanian, Fakultas Pertanian IPB, lulus
pada tahun 1995. Pada tahun 2003 penulis menyelesaikan program Magister
Sains di Program Studi Agronomi, Program Pascasarjana IPB. Pada tahun 2005
penulis berkesempatan melanjutkan studi ke program Doktor pada program studi
dan perguruan tinggi yang sama dengan beasiswa BPPS dari Departemen
Pendidikan Nasional pada tahun (2005-2008).
Penulis bekerja sebagai Staf Pengajar di Departemen Agronomi dan
Hortikultura, Fakultas Pertanian IPB sejak tahun 1997 sampai sekarang. Sesuai
dengan pengembangan staf dan bidang ilmu di Departemen, penulis menjadi
anggota Bagian Ekofisiologi Tanaman.

DAFTAR ISI
Halaman
DAFTAR TABEL ........................................................................................ iv
DAFTAR GAMBAR ................................................................................... vii
PENDAHULUAN ....................................................................................... 1 Latar Belakang ................................................................................. 1 Tujuan Penelitian .............................................................................. 3 Hipotesis .......................................................................................... 3
TINJAUAN PUSTAKA ............................................................................... 5 Botani dan Morfologi Echinochloa cruss-galli .................................. 5 Ekologi Gulma E. crus-galli ............................................................. 8 Fisiologi E. crus-galli ....................................................................... 10 Marka Molekuler Simple Sequence Repeat (SSR) ............................. 16
PERCOBAAN PENDAHULUAN TENTANG PERTUMBUHAN DAN PRODUKSI PADI PADA BERBAGAI TINGKAT POPULASI GULMA Echinochloa cruss-galli YANG BERASAL DARI TIGA LOKASI .............. 19
Pendahuluan ..................................................................................... 19 Bahan dan Metode ............................................................................ 20 Hasil dan Pembahasan ...................................................................... 21
Pertumbuhan Tanaman Padi .................................................... 21 Komponen Produksi Padi ........................................................ 26 Produksi Gabah ....................................................................... 27
Kesimpulan ...................................................................................... 28
KERAGAMAN MORFOLOGI DAN GENETIK AKSESI GULMA Echinochloa cruss-galli ASAL JAWA BARAT ............................................ 29
Pendahuluan ..................................................................................... 29 Metode Penelitian ............................................................................. 30 Hasil dan Pembahasan ...................................................................... 34
Karakteristik Lingkungan Habitat Asal Aksesi E. crus-galli ..... 34 Keragaman Morfologi Aksesi Gulma E. crus-galli di Habitat Asal .......................................................................................... 35 Keragaman Morfologi Aksesi Gulma E. crus-galli di Rumah Kaca ......................................................................................... 38 Keragaman Genetik Aksesi Gulma E. crus-galli ....................... 42 Pembahasan.............................................................................. 43
Kesimpulan ...................................................................................... 46 IDENTIFIKASI POTENSI ALELOPATI BEBERAPA AKSESI GULMA Echinochloa cruss-galli ASAL JAWA BARAT…. ....................................... 47
Pendahuluan ..................................................................................... 47 Bahan dan Metode ............................................................................ 48

ii
Hasil dan Pembahasan ...................................................................... 49 Potensi Alelopati Aksesi Gulma E. crus-galli .......................... 49 Hubungan antara Potensi Alelopati dengan Keragaman Genetik Aksesi Gulma E. crus-galli …………………………. 50 Kandungan Senyawa Alelopati Aksesi E. crus-galli ………... 52 Pembahasan ……….................................................................... 53
Kesimpulan ...................................................................................... 54 PENGARUH AKSESI GULMA Echinochloa cruss-galli TERHADAP PERTUMBUHAN DAN PRODUKSI PADI (PERCOBAAN RUMAH KACA) ......................................................................................................... 55
Pendahuluan ..................................................................................... 55 Bahan dan Metode ............................................................................. 56 Hasil dan Pembahasan ...................................................................... 57
Pertumbuhan Vegetatif Tanaman Padi ..................................... 57 Pertumbuhan Generatif Tanaman Padi ..................................... 61 Pembahasan ............................................................................. 62
Kesimpulan ...................................................................................... 64
PENGARUH AKSESI DAN TINGKAT POPULASI Echinochloa cruss-galli TERHADAP PERTUMBUHAN DAN PRODUKSI PADI (PERCOBAAN LAPANGAN) ..................................................................... 65
Pendahuluan ..................................................................................... 65 Bahan dan Metode ............................................................................. 66 Hasil dan Pembahasan ...................................................................... 67
Pertumbuhan Vegetatif Tanamaman Padi ................................ 67 Komponen Hasil Tanaman Padi ............................................... 75 Hasil Produks i Tanaman Padi .................................................. 78 Pembahasan.............................................................................. 79
Kesimpulan dan Saran ...................................................................... 81 PENDUGAAN DERAJAT KOMPETISI GULMA E. crus-galli MELALUI METODE REPLACEMENT SERIES ............................................................ 83 Pendahuluan ..................................................................................... 83 Bahan dan Metode ............................................................................ 84 Hasil dan Pembahasan ...................................................................... 85
Kompetisi di Atas Permukaan Tanah ....................................... 85 Kompetisi di Bawah Permukaan Tanah .................................... 88 Pengaruh Kompetisi terhadap Produksi Biomass Total ............. 92 Pengaruh Kompetisi terhadap Komponen Hasil dan Hasil Produksi ................................................................................... 93 Penguasaan Sarana Tumbuh ..................................................... 94 Koefisien Pendesakan ............................................................... 98 Nilai Agresivitas (A) ................................................................ 98 Pembahasan ............................................................................. 99
Kesimpulan dan Saran ...................................................................... 101

iii
KAJIAN FISIOLOGI KOMPETISI ANTARA PADI DENGAN GULMA Echinochloa crus-galli ................................................................................. 103 Pendahuluan ..................................................................................... 103 Bahan dan Metode ............................................................................ 104 Hasil dan Pembahasan ...................................................................... 106 Pertumbuhan Vegetatif Tanaman Padi ..................................... 106
Fisiologi Kompetisi antara Tanaman Padi dengan Gulma E. crus-galli .................................................................................. 112
Pertumbuhan Generatif Tanaman Padi ..................................... 115 Komponen Produksi Tanaman Padi ......................................... 115
Hasil Produksi Tanaman Padi .................................................. 117 Mutu Hasil Panen ..................................................................... 119 Indeks Panen ............................................................................ 119 Pembahasan.............................................................................. 120
Kesimpulan ...................................................................................... 123 PEMBAHASAN UMUM ............................................................................ 125 Keragaman Morfologi dan Genetik serta Implikasinya dalam
Manajemen Gulma ............................................................................ 125 Potensi Alelopati Gulma E. crus-galli ................................................ 128 Keragaman Aksesi Gulma dalam Penekanan terhadap Pertumbuhan
dan Produksi Padi ............................................................................. 129 Respon Tanaman Padi terhadap Tingkat Populasi Gulma E. crus-
galli ................................................................................................... 131 Fisiologi Kompetisi Padi-Gulma ........................................................ 132 KESIMPULAN DAN SARAN .................................................................... 135 Kesimpulan ....................................................................................... 135 Saran ................................................................................................. 137
DAFTAR PUSTAKA .................................................................................. 138 LAMPIRAN ................................................................................................ 147

iv
DAFTAR TABEL
No. Halaman
1. Pengaruh aksesi gulma terhadap tinggi tanaman padi ........................ 21
Teks
2. Pengaruh aksesi dan populasi E. crus-galli terhadap jumlah anakan tanaman padi ..................................................................................... 22
3. Jumlah daun tanaman padi pada perlakuan populasi E. crus-galli ..... 23
4. Bobot kering tajuk dan akar padi pada perlakuan populasi E. crus-galli .................................................................................................. 24
5. Panjang, lebar, luas daun bendera padi pada perlakuan populasi E. crus-galli .......................................................................................... 25
6. Jumlah anakan produktif, panjang malai dan kepadatan malai padi pada perlakuan aksesi dan populasi E. crus-galli ............................... 26
7. Pengaruh aksesi dan populasi E. crus-galli terhadap bobot gabah dan persentase kehampaan ...................................................................... 28
8. Deskripsi asal-usul 16 aksesi gulma E. crus-galli di Jawa Barat ......... 31 9. Primer Simple Sequence Repeat (SSR) dan sekuen nukleotidanya
yang digunakan dalam analisis genetic aksesi E. crus-galli .............. 32 10. Karakteristik lingkungan dari habitat asal aksesi gulma E. crus-galli
dan lokasi penanaman gulma E. crus-galli ........................................ 34 11. Karakter morfologi aksesi gulma E. crus-galli di habitat asal dari
berbagai lokasi di Jawa Barat ............................................................ 35 12. Nilai eigenvalue berdasarkan karakter morfologi di habitat asal ......... 36
13. Nilai komponen utama berdasarkan karakter morfologi pada habitat asal aksesi gulma E. crus-galli .......................................................... 37
14. Karakter vegetatif aksesi gulma E. crus-galli asal Jawa Barat di rumah kaca (Bogor, 250 m dpl) ......................................................... 38
15. Karakter generatif aksesi gulma E. crus-galli asal Jawa Barat di rumah kaca (Bogor, 250 m dpl) ......................................................... 39
16. Nilai eigenvalue berdasarkan karakter morfologi di rumah kaca ....... 41 17. Nilai komponen utama berdasarkan karakter morfologi di rumah
kaca .................................................................................................. 41 18. Pengaruh aksesi gulma E. crus-galli terhadap penghambatan
plumula dan radikula kecambah padi saat 14 hari setelah semai ......... 49 19. Penggolongan potensi alelopati aksesi gulma E. crus-galli ............... 52

v
20. Kandungan senyawa dalam ekstrak akar beberapa aksesi gulma E. crus-galli asal Jawa Barat .................................................................. 52
21. Pengaruh aksesi gulma E. crus-galli terhadap pertumbuhan vegetatif tanaman padi ..................................................................................... 58
22. Pertumbuhan vegetatif gulma E. crus-galli ....................................... 58
23. Pengaruh aksesi gulma E. crus-galli terhadap jumlah anakan produktif, panjang malai, dan kepadatan malai padi ........................... 61
24. Pengaruh aksesi gulma E. crus-galli terhadap panjang dan lebar daun bendera tanaman padi ....................................................................... 62
25. Pengaruh aksesi gulma E. crus-galli terhadap bobot gabah total, bobot gabah isi, bobot gabah hampa, dan bobot 100 butir ................. 62
26. Jumlah anakan tanaman padi pada perlakuan aksesi dan populasi gulma E. crus-galli ............................................................................ 69
27. Jumlah daun tanaman padi pada perlakuan aksesi dan populasi gulma E. crus-galli ............................................................................ 69
28. Bobot kering akar tanaman padi pada interaksi aksesi dan populasi gulma E. crus-galli ............................................................................ 70
29. Bobot kering akar E. crus-galli pada pertanaman padi hibrida ........... 71 30. Bobot kering tajuk tanaman padi pada interaksi aksesi dan populasi
gulma E. crus-galli ........................................................................... 71 31. Bobot kering tajuk E. crus-galli pada tanaman padi hibrida ............... 72
32. Panjang akar tanaman padi pada perlakuan aksesi dan populasi gulma E. crus-galli ........................................................................... 72
33. Indeks luas daun tanaman padi dan gulma E. crus-galli saat 8 MST dari berbagai perlakuan ..................................................................... 73
34. Laju tumbuh relatif tanaman padi dan gulma E. crus-galli dari umur 4 MST sampai dengan 8 MST .......................................................... 74
35. Kandungan hara tajuk tanaman padi .................................................. 75 36. Kandungan hara tajuk gulma E. crus-galli ......................................... 75
37. Jumlah anakan produktif dan panjang malai tanaman padi pada perlakuan aksesi dan populasi E. crus-galli ....................................... 76
38. Pengaruh interaksi antara aksesi dengan populasi gulma E. crus-galli terhadap kepadatan malai tanaman padi ............................................. 76
39. Pengaruh aksesi dan populasi E. crus-galli terhadap jumlah gabah per malai dan persentase kehampaan ................................................. 77
40. Gabah kering panen dan gabah kering giling tanaman padi pada perlakuan populasi E. crus-galli......................................................... 78
41. Pengaruh interaksi antara aksesi dan populasi gulma E. crus-galli terhadap bobot gabah 1 000 butir tanaman padi ................................. 79

vi
42. Pengaruh populasi tanaman terhadap tinggi tanaman dan jumlah anakan padi dan gulma E. crus-galli pada 9 MST ............................ 86
43. Pengaruh populasi tanaman per pot terhadap jumlah daun, panjang daun, dan lebar daun padi dan gulma E. crus-galli pada 9 MST ........ 87
44. Pengaruh populasi tanaman per pot terhadap komponen hasil tanaman padi .................................................................................... 93
45. Pengaruh populasi tanaman per pot terhadap komponen hasil gulma E. crus-galli ...................................................................................... 94
46. Pengaruh kepadatan populasi terhadap bobot gabah total, bobot gabah isi, bobot gabah hampa, dan persentase gabah hampa ............. 94
47. Hasil dugaan produksi biji tanaman padi pada pertanaman monokultur ........................................................................................ 95
48. Hasil dugaan produksi biji gulma E. crus-galli pada pertanaman monokultur ........................................................................................ 96
49. Penguasaan sarana tumbuh tanaman padi pada pertanaman tumpangsari dengan gulma E. crus-galli ............................................ 96
50. Nilai koefisien pendesakan (KP) pada pertanaman tumpangsari padi dengan gulma E. crus-galli ................................................................ 98
51. Nilai agresivitas tanaman padi dan gulma E. crus-galli pada pertanaman tumpangsari .................................................................... 99
52. Pengaruh varietas dan populasi gulma
53. Kandungan hara daun beberapa varietas tanaman padi pada populasi gulma E.crus-galli yang berbeda
E.crus-galli/m² terhadap ILD tanaman padi pada 8 MST ................................................................. 108
54. Efisiensi serapan hara pada beberapa varietas tanaman padi dan populasi gulma E. crus-galli
....................................................... 112
55. LAR, NAR, RGR, dan CGR beberapa varietas padi pada beberapa populasi gulma E.crus-galli
............................................................. 113
56. Waktu heading tanaman padi pada perlakuan varietas padi dan populasi gulma E.crus-galli ............................................................... 115
............................................................. 114
57. Pengaruh interaksi varietas padi dan populasi gulma E.crus-galli terhadap panjang malai, jumlah biji per malai .................................... 116
58. Bobot 1 000 butir gabah pada beberapa varietas tanaman padi dan pada berbagai populasi gulma E.crus-galli
59. Pengaruh tingkat populasi gulma E. crus-galli terhadap indeks panen pada beberapa varietas tanaman padi ................................................. 120
........................................ 117

vii
Lampiran
1. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Karawang (K, 37 m dpl) berdasarkan analisis GCMS ......... 148
2. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Subang (S, 29 m dpl) berdasarkan analisis GCMS .............. 149
3. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Cikampek (C, 40 m dpl) berdasarkan analisis GCMS ......... 150
4. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Indramayu (I, 16 m dpl) berdasarkan analisis GCMS .......... 151
5. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Cianjur (Ta, 250 m dpl) berdasarkan analisis GCMS .......... 152
6. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Sukabumi (Tc, 750 m dpl) berdasarkan analisis GCMS ...... 153
7. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Cianjur (Td, 1000 m dpl) berdasarkan analisis GCMS ........ 154
8. Kandungan senyawa kimia dalam ekstrak akar gulma E. crus-galli aksesi gulma asal Pangalengan (Te, 1250 m dpl) berdasarkan analisis GCMS ................................................................................................. 155
9. Kandungan senyawa kimia dalam ekstrak akar gulma E. crus-galli aksesi gulma asal Pangalengan (Tf, 1500 m dpl) berdasarkan analisis GCMS ................................................................................................. 156

viii
DAFTAR GAMBAR
No. Halaman
1. Bagan alir tahapan penelitian ............................................................ 4
Teks
2. Echinochloa cruss-galli (L.) Beauv .................................................. 6 3. Hubungan kekerabatan aksesi gulma E. crus-galli dari berbagai
lokasi di Jawa Barat berdasarkan karakter morfologi di habitat asal .. 36 4. Pengelompokan aksesi gulma E. crus-galli berdasarkan analisis
komponen utama .............................................................................. 37 5. Dendogram kekerabatan aksesi gulma E. crus-galli berdasarkan
karakter morfologi di rumah kaca ..................................................... 40 6. Diagram pencar (komponen I-II) aksesi gulma E. crus-galli
berdasarkan karakter vegetative di rumah kaca ................................. 42 7. Dendogram kekerabatan genetik aksesi gulma E. crus-galli
berdasarkan marka molekuler Simple Sequence Repeat (SSR) ........... 43 8. Dendogram kekerabatan aksesi gulma E. crus-galli dari berbagai
habitat padi sawah di Jawa Barat berdasarkan potensi alelopati.......... 51 9. Pengaruh aksesi gulma E. crus-galli terhadap tinggi tanaman padi .... 57
10. Pengaruh aksesi gulma E. crus-galli terhadap jumlah anakan tanaman padi .................................................................................... 59
11. Pengaruh aksesi gulma E. crus-galli terhadap panjang akar tanaman padi .................................................................................................. 60
12. Bobot tajuk tanaman padi pada perlakuan aksesi gulma E. crus-galli .......................................................................................................... 60
13. Bobot kering akar tanaman padi pada perlakuan aksesi gulma E. crus-galli .......................................................................................... 61
14. Perlakuan populasi gulma E. crus-galli per m² pada pertanaman padi 66 15. Tinggi tanaman padi pada 6 MST pada perlakuan aksesi dan tingkat
populasi gulma E. crus-galli .............................................................. 68 16. Hubungan antara populasi gulma E. crus-galli dengan produksi
gabah ................................................................................................. 79 17. Skema penanaman padi dan gulma E. crus-galli di pot pada
percobaan replacement series ........................................................... 84 18. Bobot kering biomasa tajuk tanaman padi dan gulma E. crus-galli
pada pertanaman monokultur dan pertanaman campuran ................... 88

ix
19. Panjang akar tanaman padi dan gulma E. crus-galli pada pertanaman monokultur dan pertanaman campuran ............................................. 88
20. Bobot kering biomass akar padi dan gulma E. crus-galli pada pertanaman monokultur dan pertanaman campuran ........................... 89
21. Serapan hara N tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran ................................................................. 90
22. Serapan hara P tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran ................................................................. 90
23. Serapan hara K tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran ................................................................. 91
24. Serapan hara Ca tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran ................................................................. 91
25. Serapan hara Mg tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran ................................................................. 92
26. Bobot kering biomass total tanaman padi dan gulma E. crus-galli pada pertanaman monokultur dan pertanaman campuran ................... 93
27. Penguasaan sarana tumbuh tanaman padi dan E. crus-galli ................ 97
28. Hubungan antara produksi tanaman padi dan gulma E. crus-galli pada pertanaman monokultur dan tumpangsari .................................. 97
29. Perlakuan populasi gulma E.crus-galli per m² .................................... 105 30. Perkembangan tinggi tanaman beberapa varietas padi pada perlakuan
populasi gulma E.crus-galli ............................................................... 106 31. Pengaruh interaksi varietas padi dan populasi gulma E.crus-galli
terhadap jumlah anakan ..................................................................... 107 32. Pengaruh interaksi varietas padi dan populasi gulma E.crus-galli
terhadap jumlah daun tanaman padi ................................................... 108 33. Bobot kering tajuk beberapa varietas tanaman padi ............................ 109
34. Pengaruh populasi gulma E.crus-galli terhadap bobot kering tajuk tanaman padi ..................................................................................... 109
35. Perkembangan bobot kering akar beberapa varietas tanaman padi ...... 110 36. Perkembangan bobot kering akar tanaman padi pada beberapa
populasi gulma E.crus-galli ............................................................... 110 37. Bobot kering total (tajuk dan akar) pada perlakuan varietas dan
populasi gulma E.crus-galli ............................................................... 111 38. Panjang akar beberapa varietas tanaman padi pada perlakuan
populasi gulma E.crus-galli ............................................................... 111 39. Pengaruh populasi gulma E.crus-galli terhadap jumlah anakan
produktif beberapa varietas tanaman padi .......................................... 116

x
40. Hubungan tingkat populasi gulma dan produksi gabah kering giling pada beberapa varietas padi ............................................................... 118
41. Dugaan hasil produksi gabah/hektar dari berbagai populasi gulma E.crus-galli/m² .................................................................................. 118
42. Persentase kehampaan pada beberapa varietas padi pada berbagai populasi gulma E.crus-galli ............................................................... 119

PENDAHULUAN
Latar Belakang
Tanaman padi (Oryza sativa) merupakan komoditas yang strategis di
Indonesia karena beras merupakan sumber makanan pokok bagi hampir seluruh
rakyat Indonesia. Kebutuhan beras di Indonesia semakin meningkat sejalan
dengan pertambahan jumlah penduduk. Jumlah penduduk Indonesia tahun 2011
mencapai 241 juta jiwa dan kebutuhan beras mencapai 34 juta ton per tahun (BPS,
2011). Produksi padi pada tahun 2011 berdasarkan Angka Ramalan II (ARAM II)
diperkirakan mencapai 68.06 juta ton gabah kering giling (GKG) atau 38.2 juta
ton beras. Kebutuhan beras pada tahun 2025 diperkirakan mencapai 48.5 juta ton
atau setara dengan 70 juta ton GKG.
Untuk memenuhi kebutuhan beras tersebut diperlukan usaha peningkatan
produksi dan produktivitas padi di Indonesia. Peningkatan produksi beras tahun
2011 dibandingkan tahun 2010 disebabkan oleh peningkatan luas areal (0.11%)
dan peningkatan produktivitas (1.24%) (BPS 2011). Pada tahun-tahun
mendatang, upaya peningkatan produksi beras akan menghadapi banyak kendala
diantaranya perubahan fungsi lahan sawah menjadi lahan non pertanian yang
mencapai 187 720 hektar/tahun (Ditjen Pengelolaan Lahan dan Air 2005),
degradasi kesuburan lahan yang saat ini mencapai 4 juta hektar dari degradasi
berat (50%) hingga degradasi rendah (0.8%) (Badan Litbang Pertanian 2011),
serta adanya serangan organisme pengganggu tanaman.
Salah satu organisme pengganggu tanaman yang dapat menurunkan
produksi dan produktivitas tanaman padi adalah gulma. Kehilangan hasil akibat
gulma di seluruh dunia diperkirakan mencapai 10-15% (Smith 1968; Smith 1983;
Zoschke 1990; Baltazar dan De Datta 1992), bahkan kehilangan hasil dapat
mencapai 86% jika tanpa dilakukan pengendalian gulma (Kropff 1993).
Salah satu gulma penting dan dominan pada lahan padi sawah di Indonesia
adalah gulma jajagoan (Echinochloa crus-galli) (Ali dan Sankaran 1984; Ali
1985). Gulma E. crus-galli menjadi masalah utama pada budidaya tanaman padi
sawah dan merupakan penyebab kehilangan hasil produksi yang utama pada
produksi padi sawah (Gealy et al. 2003; Haefele et al. 2004). Penurunan

2
produksi padi akibat gulma E. crus-galli dapat mencapai 46-59% (Sultana 2000;
Chin 2001), 57-95% (Ahn dan Chung 2000), bahkan hingga 97% (Islam dan
Karim 2003).
Penurunan produksi tanaman padi oleh gulma E. crus-galli dapat terjadi
karena kompetisi, alelopati, dan menjadi inang hama penyakit tanaman padi
(alelomediasi). Kompetisi gulma E. crus-galli menyebabkan penurunan hasil
produksi akibat penurunan jumlah anakan, jumlah malai, dan jumlah gabah per
malai (Tindall et al. 2005). Gulma E. crus-galli berpotensi mengeluarkan
senyawa alelopati yang dapat menghambat pertumbuhan tanaman (Putnam 1986).
Eksudat akar E. crus-galli yang berupa senyawa p-Hidroxymandelic menunjukkan
efek alelopati dan menekan pertumbuhan tajuk dan pemanjangan akar tanaman
padi (Yamamoto et al. 1999; Xuan et al. 2006). Gulma E. crus-galli dapat
menjadi inang perantara bagi Leptocorisa oratorius, Acrocylindricum oryzae,
Corticium sasakii, dan Rhynchosporium oryzae (Tjitrosemito 1994). Selain
menurunkan kuantitas hasil, keberadaan gulma E. crus-galli juga menyebabkan
penurunan kualitas dalam produksi benih akibat tercampurnya benih padi dengan
biji-biji E. crus-galli.
Gulma E. crus-galli sulit dikendalikan karena kemiripan morfologi dengan
tanaman padi. Gulma ini menjadi lebih bermasalah pada budidaya tanaman padi
ketika cara tanam padi berubah dari cara tanam pindah (transplanting) menjadi
cara tebar benih langung (direct planting). Keberadaan gulma E. crus-galli pada
pertanaman padi sawah dapat menurunkan pendapatan petani padi akibat
peningkatan biaya pengendalian gulma.
Usaha peningkatan produksi yang dilakukan oleh pemerintah harus
diimbangi dengan upaya penyelamatan kehilangan hasil akibat organisme
pengganggu tanaman. E. crus-galli memiliki daya adaptasi yang luas pada
kondisi lingkungan yang bervariasi (Galinato et al. 1999). E. crus-galli yang
berasal dari habitat yang berbeda diduga memiliki daya kompetisi yang berbeda
pula. Studi tentang potensi aksesi gulma E. crus-galli dalam menurunkan
produksi padi masih jarang dilakukan di Indonesia. Studi keragaman morfologi
dan genetik serta tingkat kompetisi aksesi gulma Echinochloa crus-galli dari
beberapa habitat sawah di Jawa Barat sangat penting dilakukan. Pengetahuan

3
tentang karakter dan perilaku gulma tersebut dapat menjadi dasar bagi
pengembangan teknik pengendalian di lapangan sehingga dapat mendukung usaha
peningkatan produksi padi nasional.
Tujuan Penelitian
Secara umum penelitian ini bertujuan untuk mengkaji keragaman
morfologi dan genetik aksesi gulma E. crus-galli serta menganalisis tingkat
kompetisinya terhadap tanaman padi sawah. Adapun tujuan khusus penelitian ini
adalah :
1. Menganalisis keragaman morfologi dan genetik aksesi E. crus-galli asal
Jawa Barat
2. Mengidentifikasi potensi alelopati aksesi E. crus-galli asal Jawa Barat
3. Mempelajari pengaruh aksesi dan tingkat populasi E. crus-galli terhadap
pertumbuhan dan produksi padi sawah
4. Menduga derajat kompetisi gulma E. crus-galli melalui pendekatan
replacement series
5. Mempelajari fisiologi kompetisi antara padi dengan gulma E. crus-galli.
Hipotesis
1. Aksesi gulma E. crus-galli asal Jawa Barat memiliki keragaman karakter
morfologi dan genetik. Keragaman morfologi yang terjadi di lapangan
disebabkan oleh keragaman genetik dan oleh kemampuan plastisitas fenotipik.
2. Tiap aksesi gulma E. crus-galli asal Jawa Barat memiliki potensi alelopati
yang berbeda dan memiliki perbedaan kemampuan dalam menurunkan
pertumbuhan dan produksi tanaman padi sawah.
3. Gulma E. crus-galli memiliki daya kompetisi yang lebih kuat dibandingkan
dengan tanaman padi sawah dalam memperebutkan sumberdaya.
4. Perbedaan populasi dan asal aksesi gulma E. crus-galli menyebabkan
perbedaan respon pertumbuhan dan produksi tanaman padi sawah.
5. Kompetisi gulma E. crus-galli terhadap tanaman padi sawah menghambat
proses fisiologi tanaman padi.
4
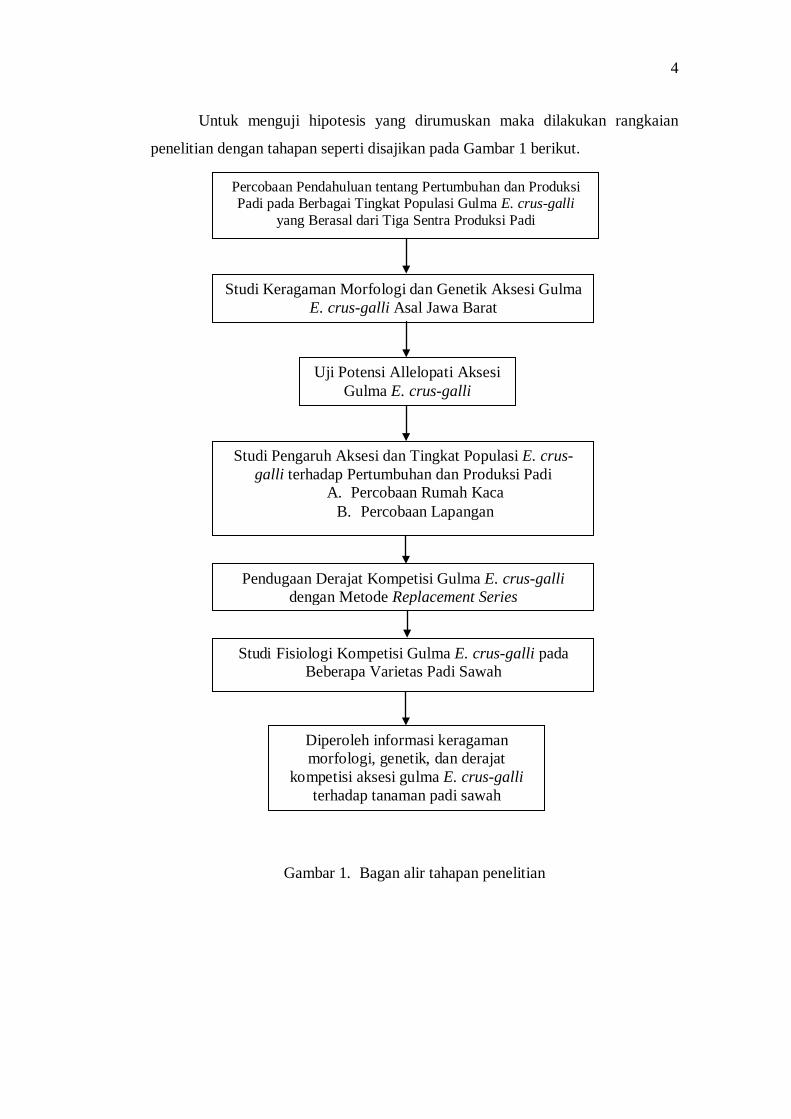
Untuk menguji hipotesis yang dirumuskan maka dilakukan rangkaian
penelitian dengan tahapan seperti disajikan pada Gambar 1 berikut.
Gambar 1. Bagan alir tahapan penelitian
Studi Keragaman Morfologi dan Genetik Aksesi Gulma E. crus-galli Asal Jawa Barat
Pendugaan Derajat Kompetisi Gulma E. crus-galli dengan Metode Replacement Series
Uji Potensi Allelopati Aksesi Gulma E. crus-galli
Percobaan Pendahuluan tentang Pertumbuhan dan Produksi Padi pada Berbagai Tingkat Populasi Gulma E. crus-galli
yang Berasal dari Tiga Sentra Produksi Padi
Studi Pengaruh Aksesi dan Tingkat Populasi E. crus-galli terhadap Pertumbuhan dan Produksi Padi
A. Percobaan Rumah Kaca B. Percobaan Lapangan
Studi Fisiologi Kompetisi Gulma E. crus-galli pada Beberapa Varietas Padi Sawah
Diperoleh informasi keragaman morfologi, genetik, dan derajat
kompetisi aksesi gulma E. crus-galli terhadap tanaman padi sawah

TINJAUAN PUSTAKA
Botani dan Morfologi Echinochloa crus-galli
Botani
Gulma E. crus-galli merupakan gulma semusim kelas Monocotyledon,
subkelas Commelinidae, ordo Cyperales, suku Poaceae/Graminae, dan marga
Echinochloa. Spesies E. crus-galli meliputi 3 subspesies, yaitu E. crus-galli var.
crus-galli, E. crus-galli var. particola dan E. crus-galli var. formosensis (Kim
1994). E. crus-galli adalah tumbuhan hexaploid (2n=6x=54). Gulma E. crus-galli
memiliki distribusi geografis yang luas dan keragaman dari morfologi dan
ekologi yang diduga terkait dengan sifat allohexaploid dari spesies ini (Yabuno
1983).
E. crus-galli diperkirakan berasal dari India, tersebar pada daerah tropis
dan sub tropis di seluruh negara Asia Tenggara dan Asia selatan serta Australia
(Holm et al. 1977). Beberapa nama umum untuk rumput ini antara lain barnyard
grass, japanese millet, cockspur, dan watergrass. Di Indonesia E. crus-galli
dikenal dengan nama gagajahan, jajagoan, jawan, jawan pari, suket ngawan
(Jawa) (Moenandir 1993; Galinato et al. 1999).
Morfologi
Rumput E. crus-galli sangat mirip dengan tanaman padi pada saat masih
muda. E. crus-galli memiliki penampilan tegak, memiliki tinggi sekitar 20-150
cm (Soerjani et al. 1987), bahkan bisa mencapai 200 cm (Galinato et al. 1999).
E. crus-galli memiliki daun yang tegak atau rebah pada dasarnya.
Daunnya memiliki ukuran panjang sampai 35 cm dan lebar 0.5-1.5 cm. Warna
daun rumput ini hijau sampai hijau keabuan. Setiap daun memiliki pelepah yang
tidak berambut dan memiliki panjang 9-13 cm. Pelepah daun umumnya berwarna
kemerahan di bagian bawahnya. Helaian daun berukuran 5-65 cm x 6-22 mm,
bersatu dengan pelepah, berbentuk linear dengan bagian dasar yang lebar dan
melingkar dan bagian ujung yang meruncing. Permukaan daun rata, agak kasar
dan menebal di bagian tepi (Duke 1996). Helaian daun memiliki beberapa rambut
halus pada bagian dasarnya dan agak lebat pada permukaan daun (Fishel 2000).

6
Daerah pangkal daun dapat digunakan untuk membedakan daun E. crus-galli dan
daun padi. Pangkal daun E. crus-galli tidak memiliki ligula dan aurikel,
sedangkan pangkal daun padi memiliki ligula yang bermembran dan aurikel yang
berbulu (Itoh 1991).
Gambar 2. Echinochloa crus-galli (L.) Beauv (Soerjani et al. 1987)
a. zona helaian daun; b. Spikelet dengan rambut pendek; c. Spikelet dengan rambut panjang dari raceme yang sama; d. Glume yang paling bawah (G1) tampak belakang (dibuka); e. Glume teratas (G2), tampak depan; f. Lemma terbawah (L1), tampak depan; g. Palea terbawah (P1), tampak depan; h. Lemma teratas (L2), tampak depan; i. Palea teratas (P2), tampak depan; j. Kariopsis, dua sisi.

7
Batang berbentuk silindris dengan pith yang menyerupai spons putih di
bagian dalamnya. Di lahan sawah, anakan pertama dari E. crus-galli muncul
10 hari setelah perkecambahan, dan biasanya sekitar 15 anakan yang terbentuk
(Galinato et al. 1999). E. crus-galli memiliki jenis akar serabut dan tebal. Akar
E. crus-galli dihasilkan pada setiap ruasnya (Soerjani et al. 1987).
Bunga berupa malai yang berada di ujung dengan 5-40 bunga majemuk
bulir yang mempunyai tipe raceme, dengan cabang-cabang pendek yang menaik.
Bunga majemuk terdiri atas banyak spikelet yang berbelok pada satu sisi,
berbentuk tegak pada awalnya tetapi selanjutnya sering membengkok ke bawah.
Panjang malai bisa mencapai 5-21 cm. Malai kaku dengan permukaan yang agak
kasar. Bulir terbawah merupakan bulir yang paling panjang, sekitar 1.75-8 cm,
sedangkan bulir yang paling atas sangat pendek. Setiap bulir terdapat susunan
spikelet yang berselang-seling di setiap sisinya (Soerjani et al. 1987).
Spikelet tersusun soliter pada bulir paling atas dan susunannya bisa
mencapai 2-4 spikelet dan pada bulir bagian bawah susunan spikelet bisa
mencapai 4-10 spikelet (Soerjani et al. 1987). Spikelet tebal dan padat, sedikit
berbentuk elips dengan panjang 3.2-3.5 mm. Spikelet biasanya sedikit berambut
dan terkadang terdapat rambut yang tebal dan kaku yang panjangnya dapat
mencapai 13 mm. Spikelet berwarna kehijauan dan sedikit berwarna ungu
(Ampong-Nyarko dan De Datta 1991).
Stamen pada E. crus-galli berjumlah 3 dengan anther berwarna kuning.
Jumlah putik ada 2 dengan stigma berbulu, berwarna ungu, menonjol keluar di
bawah ujung spikelet. Buah E. crus-galli disebut caryopsis, berbentuk lonjong
dengan panjang 1.5-2 mm, berbentuk ovoid sampai obovoid (Galinato et al.
1999). Lemma dari floret yang pertama memiliki permukaan yang datar atau
sedikit cembung atau tumpul. Glume bagian bawah memiliki panjang sekitar 1.5-
2.5 mm, berbentuk ovate, memendek dan memiliki ujung yang memendek secara
bertahap. Glume bagian atas memiliki panjang yang sama dengan spikelet,
berbentuk ovate-oblong, runcing, memiliki rambut yang tebal dan kaku sepanjang
0.5-3 mm serta berambut pendek (Galinato et al. 1999).
Biji yang tua berwarna kecoklat-coklatan sampai kehitaman. Produksi biji
bervariasi dari 2 000 – 40 000 benih per tanaman pada daerah bergulma.

8
(Ampong-Nyarko dan De Datta 1991). E. crus-galli mampu menghasilkan lebih
dari 1 000 kg benih/ha (Galinato et al. 1999).
Perbanyakan dan Penyebaran
Gulma E. crus-galli berperan sebagai gulma pada 36 jenis tanaman
budidaya di 61 negara. Jenis rumput ini memperbanyak diri secara generatif
melalui biji yang seringkali tercampur dengan benih padi (Galinato et al. 1999).
Gulma ini bereproduksi dengan cara penyerbukan sendiri atau penyerbukan
silang. E. crus-galli melakukan penyerbukan silang dengan menggunakan bantuan
angin. Biji E. crus-galli dapat menyebar melalui saluran irigasi, hewan, burung,
pengangkutan biji padi dan mesin pertanian atau peralatan pertanian lainnya (Itoh
1991).
Perkembangan yang sangat cepat dan agresif dari E. crus-galli terkait
dengan pertumbuhannya yang sangat cepat, produksi benih yang tinggi, dormansi
benih dan daya adaptasi yang tinggi di bawah kondisi lahan pertanian yang
berbeda (Bahrendt dan Hanf 1979). E. crus-galli adalah spesies yang sangat
bervariasi, memiliki banyak bentuk dan variasi dengan waktu berbunga dan
menghasilkan biji yang sangat berbeda antara satu dengan yang lain.
Ekologi Gulma E. crus-galli
Lingkungan Tumbuh
E. crus-galli merupakan gulma yang beradaptasi pada daerah berair, dan
tumbuh baik pada kelembaban tanah 80 % dari water capasity (Ampong-Nyarko
dan De Datta 1991). Pertumbuhan E. crus-galli sangat baik pada jenis tanah
berpasir dan berlempung terutama apabila kandungan nitrogennya tinggi (Kropff
dan Van Laar 1993).
Gulma ini mampu terus tumbuh walaupun hanya sebagian dari benih yang
terendam. Perkecambahan 30% lebih baik di tanah padat daripada di tanah yang
kurang padat. E. crus-galli juga mampu tumbuh dengan baik pada tanah yang
lebih kering, tetapi memiliki pertumbuhan yang lebih kecil dan menghasilkan
jumlah malai, anakan dan jumlah biji yang lebih sedikit dibandingkan pada tanah
berair (Galinato et al. 1999).

9
Menurut Soerjani et al. (1987) benih E. crus-galli tidak dapat berkecambah
pada kedalaman air lebih dari 12 cm, sedangkan menurut Kropff dan Van Laar (1993)
kedalaman air maksimum bagi perkecambahan benih E. crus-galli adalah 15 cm. Benih
yang terendam pada kedalaman lebih dari 15 cm tidak dapat berkecambah.
Benih E. crus-galli dapat hidup terus dalam waktu yang lama. Benih yang
terdapat di dalam tanah dapat hidup terus sampai 1 tahun. Benih yang disimpan di
tempat penyimpanan dalam kondisi kering dapat hidup terus sampai 7 tahun.
Kelembaban optimum untuk perkecambahan benih E. crus-galli tergantung dari
karakteristik tanah, tetapi umumnya pada 70-90% kapasitas lapang. Benih E. crus-galli
yang berada dekat dengan permukaan tanah akan berkecambah baik pada hari yang
panas (Galinato et al. 1999).
E. crus-galli dapat tumbuh pada daerah dataran rendah sampai sedang. Gulma
ini tumbuh baik pada tempat dengan penyinaran penuh sepanjang tepi perairan
(Soerjani et al. 1987). E. crus-galli membutuhkan waktu 42-64 hari untuk melengkapi
siklus hidupnya. Benih akan langsung tumbuh setelah ditanam tetapi sebagian lagi
mengalami dormansi yang bisa mencapai selama 4-48 bulan. Fotoperiodisme
mempengaruhi jumlah benih yang dorman dan intensitas dari dormansi tersebut
(Zimdahl et al. 2004).
Pembungaan dipengaruhi oleh panjang hari. Pada hari pendek pembungaan
lebih cepat terjadi. Jumlah malai dan anakan lebih besar pada hari pendek, tetapi
ukurannya kecil. Pada hari panjang (16 jam), gulma ini menghasilkan malai dengan
ukuran yang lebih besar dan jumlah benih yang lebih banyak. E. crus-galli yang
tumbuh pada daerah dengan penyinaran penuh memiliki bobot kering empat kali lebih
besar serta jumlah malai dan anakan dua kali lebih banyak daripada E. crus-galli yang
tumbuh pada daerah dengan naungan 50% (Galinato et al. 1999).
Suhu lingkungan optimum untuk perkecambahan biji adalah 32-37°C. Tingkat
perkecambahan akan menurun drastis pada suhu lingkungan di bawah 10°C atau di atas
40°C dan berhenti berkecambah pada suhu 5o
Distribusi geografis dari E. crus-galli yaitu dari 50 °LU sampai 40 °LS (Holm
et al. 1977). Di Kanada telah dilaporkan bahwa E. crus-galli ditemukan pada tempat
C. Pemanjangan kecambah sangat
tergantung pada persediaan oksigen. Benih tetap memiliki viabilitas yang tinggi
walaupun sudah disimpan dalam waktu yang lama (Galinato et al. 1999).

10
di atas 50 °LU, seperti di Edmonton (53° 33’ LU), Saskatoon (52° 07’ LU) dan Prince
Albert (53° 12’ LU) (Maun dan Barret 1986).
Plastisitas Fenotipik
Spesies tumbuhan dapat menyebar pada kondisi lingkungan secara luas, namun
kemampuan penyebaran setiap genotip terbatas. Kemampuan spesies menyebar secara
luas ditandai oleh kemampuan plastisitas fenotipik dan tingkat variasi genetik yang
tinggi (Santamaria et al. 2003). Plastisitas fenotipik adalah kemampuan suatu
organisme untuk mengubah morfologi atau fisiologinya setelah terpapar atau berada
pada kondisi lingkungan yang berbeda atau ekspresi fenotipik yang tergantung pada
lingkungan (Schlichting 1986; Thompson 1991; Sultan 2000; deWitt & Scheiner
2004). Plastisitas fenotipik memainkan peranan yang penting dalam distribusi ekologi
suatu organisme (Sultan 2003). Plastisitas fenotipik membiarkan organisme untuk
menerima kondisi lingkungan yang berbeda dengan mengubah fenotipiknya untuk
menyesuaikan diri dengan lingkungannya sehingga menjadi adaptif (Plante & Hendry
2011). Sumber perbedaan lingkungan tumbuh seperti iklim, cahaya, dan ketinggian
tempat dapat menyebabkan adanya plastisitas fenotipik (Santamaria 2003; Sultan
2003). Kemampuan plastisitas fenotipik gulma E. crus-galli telah dilaporkan oleh
banyak peneliti. Menurut Yamasue (2003) gulma E. crus-galli memiliki plastisitas
fenotipik yang luas pada karakter tinggi tanaman yang tergantung dari tanaman
tetangganya dan membentuk daun dan malai yang lebih tinggi daripada kanopi
tanaman padi.
E. crus-galli dapat memiliki kemampuan “mimikri” yaitu menyerupai tanaman
padi pada tahapan pertumbuhan tertentu dalam siklus hidupnya. Sifat ini muncul
melalui seleksi alami akibat tindakan penyiangan pada sistem pertanian yang intensif
(Barret 1983; Baki et al. 2003). Kemampuan mimikri membantu gulma E. crus-galli
untuk menghindar dari penyiangan manual (Yamasue 2003).
Fisiologi E. crus-galli
Gulma E. crus-galli termasuk tumbuhan C4. Sebagai gulma C4, E. crus-
galli menunjukkan tingkat fotosíntesis bersih yang lebih tinggi, efisiensi
penggunaan air dan nitrogen yang lebih baik dibandingkan dengan

11
tanaman/tumbuhan C3 (Ampong-Nyarko dan De Datta 1991). Gulma dengan
siklus C4 mempunyai kapasitas tinggi dalam berproduksi dan berkompetisi serta
mempunyai kebutuhan air yang lebih rendah. Pada kondisi ketersediaan air yang
rendah, gulma ini secara umum mampu bersaing terhadap tanaman padi (Baki dan
Azmi 2003).
Alelopati Gulma E. crus-galli
Tumbuh-tumbuhan juga dapat bersaing antar sesamanya secara interaksi
biokimiawi, yaitu salah satu tumbuhan mengeluarkan senyawa beracun ke
lingkungan sekitarnya dan dapat mengakibatkan gangguan pertumbuhan pada
tumbuhan yang ada di dekatnya. Interaksi biokimiawi antara gulma dan
pertanamanan antara lain menyebabkan gangguan perkecambahan biji, kecambah
jadi abnormal, pertumbuhan memanjang akar terhambat, perubahan susunan sel-
sel akar dan lain sebagainya. Beberapa species gulma menyaingi pertanaman
dengan mengeluarkan senyawa beracun dari akarnya (root exudates atau lechates)
atau dari pembusukan bagian vegetatifnya. Persaingan yang timbul akibat
dikeluarkannya zat yang meracuni tumbuhan lain disebut alelopati dan zat
kimianya disebut alelopat.
Menurut Sastroutomo (1990) senyawa alelopati dapat mempengaruhi
aktivitas tumbuhan antara lain menghambat penyerapan hara oleh akar tanaman,
pembelahan sel-sel akar, pertumbuhan tanaman, aktivitas fotosintesis,
mempengaruhi respirasi, sitesis protein, menurunkan daya permeabilitas membran
sel dan menghambat aktivitas enzim. Senyawa-senyawa kimia yang mempunyai
potensi alelopati dapat ditemukan di semua jaringan tumbuhan termasuk daun,
batang, akar, rizoma, umbi, bunga, buah, dan biji. Senyawa-senyawa alelopati itu
dapat dilepaskan dari jaringan-jaringan tumbuhan dalam berbagai cara termasuk
melalui penguapan, eksudat akar, pencucian dan pembusukan organ tumbuhan.
Beberapa gulma yang berpotensi alelopati baik yang masih hidup atau yang sudah
mati sama-sama dapat melepaskan senyawa alelopati melalui organ yang berada
di atas tanah maupun yang di bawah tanah. Menurut Putnam dan Weston (1986)
gulma E. crus-galli berpotensi mengeluarkan senyawa alelopati yang dapat
menghambat pertumbuhan tanaman.

12
Selama perkecambahan dan awal pertumbuhan, E. crus-galli menekan
pertumbuhan beberapa tanaman pertanian termasuk padi dan E. crus-galli itu
sendiri. Eksudat akar dari E. crus-galli muda menunjukkan efek alelopati dan
menekan pemanjangan akar padi. Senyawa p-Hidroxymandelic acid merupakan
allelochemical yang dikeluarkan dari akar E. crus-galli muda yang dapat menekan
kuat pertumbuhan tajuk dan pemanjangan akar padi (Yamamoto et al. 1999).
Eksudat akar E. crus-galli menekan perkecambahan dan pertumbuhan tanaman
padi, lettuce dan monochoria. Komponen yang berpotensi terlibat dalam aktivitas
phytotoxic E. crus-galli telah teridentifikasi antara lain phenolic, long-chain fatty
acids, loctones, diethyl phthalate, acenaphthene, phthalic acids, benzoic acid dan
decane. Penghambat pertumbuhan terbesar ditunjukkan oleh lactones, diikuti oleh
phenolic dan phthalic acid. Phytotoxins yang dikeluarkan akar E. crus-galli
memperlihatkan hambatan terhadap pertumbuhan tanaman indikator berdaun
lebar, tetapi kurang efektif pada tanaman padi dan E. crus-galli itu sendiri (Xuan
et al. 2006).
Produksi senyawa alelopati sangat dipengaruhi oleh beberapa faktor
lingkungan, diantaranya : 1) kualitas, intensitas dan lamanya masa penyinaran
cahaya dapat mempengaruhi produksi. Senyawa alelopati lebih banyak dihasilkan
pada kondisi cahaya ultraviolet dan periode penyinaran yang panjang; 2) jumlah
senyawa alelopati akan lebih banyak dihasilkan pada keadaan dengan kondisi
yang kekurangan hara; 3) senyawa alelopati lebih banyak dihasilkan dalam
keadaan yang mengalami gangguan kekeringan; 4) senyawa alelopati lebih
banyak dihasilkan dalam keadaan dengan suhu yang lebih rendah jika
dibandingkan dengan suhu normal bagi pertumbuhannya; 5) penggunaan hormon
seperti 2,4-D atau hidrasid maleat dapat meningkatkan senyawa alelopati (Rice
1974).
Kompetisi Padi dengan Gulma E. crus-galli
Gulma menurunkan pertumbuhan dan produksi tanaman melalui kompetisi
dalam memperebutkan cahaya, hara, air, dan ruang tumbuh (Kropff dan van Laar
1993). Tingkat kehilangan hasil akibat kompetisi tergantung pada tingkat
infestasi dan komposisi gulma. Apabila tanaman menguasai sarana tumbuh, maka

13
pertumbuhan gulma akan terhambat. Sebaliknya, apabila tanaman kurang vigor
dan tidak dapat menguasai sarana tumbuh, maka gulma akan tumbuh dengan
subur. Kompetisi tanaman dengan gulma tergantung pada beberapa faktor antara
lain lingkungan, pertumbuhan tanaman, kepadatan tanaman, fase pertumbuhan
tanaman, jenis gulma dan kepadatan gulma, fase pertumbuhan gulma, dan teknik
budidaya yang dilakukan (De Datta 1981).
Gulma E. crus-galli merupakan kompetitor yang sangat kuat terhadap
tanaman padi sehingga menurunkan pertumbuhan dan produksi tanaman padi
(Chin 2003). Pertumbuhan tinggi tanaman padi menurun akibat kompetisi dengan
gulma E. crus-galli (Perera et al. 1992; Sultana 2000). Trung et al. (1995)
melaporkan bahwa pada pertanaman padi pindah tanam, kepadatan gulma E. crus-
galli 5-10 gulma per m2 menyebabkan kehilangan hasil 7-13% dan kepadatan 15-
35 gulma per m2
menyebabkan kehilangan hasil sebesar 23-27%. Islam et al.
(2003) juga melaporkan bahwa ketika tanaman padi berkompetisi dengan 8 gulma
E. crus-galli per pot, tinggi tanaman padi menurun hingga 42.9%, indeks luas
daun (ILD) menurun sekitar 92%, jumlah anakan per tanaman menurun sekitar
72.7%, jumlah malai per rumpun menurun hingga 88.5%, dan jumlah gabah per
malai menurun hingga 63.8%.
Peubah-peubah Kompetisi
Terdapat dua jenis kompetisi yang biasa terjadi di alam yaitu kompetisi
intra spesifik dan interspesifik. Kompetisi intraspesifik adalah interaksi negatif
yang terjadi pada tumbuhan dengan jenis yang sama. Kompetisi interspesifik
adalah interaksi negatif yang terjadi pada tumbuhan yang berbeda jenis atau
disebut kompetisi antar jenis tumbuhan (Sastroutomo 1990). Menurut Eussen dan
Zulfadli (1981) kompetisi total merupakan gabungan dari kompetisi dan pengaruh
alelopati. Beberapa peubah kompetisi antara lain total hasil relatif (THR),
koefisien pendesakan, penguasaan sarana tumbuh (PST), dan agresivitas.
Total Hasil Relatif (THR) merupakan salah satu model untuk
mempelajari kompetisi antara tumbuhan dengan tumbuhan lainnya (De Wit 1960).
THR adalah peubah yang dapat digunakan untuk mengetahui adanya kompetisi
pada pertanaman campuran yang diperoleh melalui pendekatan kompetisi dengan

14
percobaan replacement series atau seri penggantian yaitu pengaturan populasi
relatif agar campurannya tetap satu (satu). Pengaturan populasi dimulai dengan
menyusun suatu seri kerapatan monokultur tumbuhan pertama dengan populasi
relatif satu, kemudian berangsur-angsur diganti sampai menjadi monokultur
tumbuhan kedua dengan populasi relatif satu. Nilai THR dirumuskan :
THR = tII
cII
tI
cI
YY
YY
+
Keterangan :
YcI
Y
= bobot kering tumbuhan 1 pada pertanaman campuran
tI
Y
= bobot kering tumbuhan 1 pada pertanaman tunggal
cII
Y
= bobot kering tumbuhan 2 pada pertanaman campuran
tII
= bobot kering tumbuhan 2 pada pertanaman tunggal
Kelebihan pendekatan ini adalah bahwa terjadi kompetisi atau tidak dapat
diketahui dari nilai THR yang diperoleh. Nilai THR < 1 menunjukkan pengaruh
yang saling merugikan atau interaksi negatif diantara kedua tumbuhan atau
menunjukkan adanya kejadian kompetisi. Nilai THR = 1 menunjukkan tidak ada
kompetisi atau menunjukkan adanya pemakaian sarana tumbuh secara bersama-
sama. Nilai THR = 1 dapat pula menunjukkan kejadian kompetisi jika salah satu
tumbuhan lebih dominan menguasai sarana tumbuh yang ada. Nilai THR >1
menunjukkan kejadian penambahan sarana tumbuh yang tidak terukur,
penggunaan sarana tumbuh yang berbeda dari masing-masing tumbuhan, kejadian
simbiosis mutualisme atau interaksi positif diantara kedua tumbuhan, dan
ketiadaan kompetisi.
Relative Crowding Coefficient (Koefisien Pendesakan). Koefisien
Pendesakan (KP) dapat menunjukkan kemampuan kompetisi suatu tumbuhan
terhadap tumbuhan lain (De Wit, 1960) yang dirumuskan sebagai berikut :
ijc = ZicampuranYimonoYi
ZjcampuranYi))()((
)(−

15
Keterangan :
Cij
Y
= koefisien pendesakan tumbuhan i terhadap tumbuhan j
i
Y
(campuran) = hasil tumbuhan i pada pertanaman campuran
i
Z
(monokultur) = hasil tumbuhan i pada pertanaman monokultur
i
Z
= rasio densitas tumbuhan i pada pertanaman campuran
j
= rasio densitas tumbuhan j pada pertanaman campuran
Nilai KP yang lebih besar menunjukkan kemampuan kompetisi suatu
tanaman terhadap tanaman lain. Persamaan di atas berlaku pula sebaliknya, yaitu
koefisien pendesakan tumbuhan II terhadap tumbuhan I. Perbandingan koefisien
pendesakan antara kedua tumbuhan dapat menunjukkan tumbuhan yang lebih
kompetitif. Nilai koefisien pendesakan lebih tinggi menunjukkan derajat
kompetisi lebih besar.
Penguasaan Sarana Tumbuh (PST). Kompetisi antara gulma dan
tanaman terjadi karena faktor tumbuh yang terbatas. Faktor yang dikompetisikan
berupa air, hara, CO2
, cahaya, dan ruang tumbuh, namun sulit bagi kita untuk
menjelaskan faktor mana yang berperan dalam peristiwa kompetisi tersebut. Studi
kompetisi dari salah satu faktor penunjang pertumbuhan (unsur kompetisi) secara
terpisah sangat sulit dilakukan karena banyaknya faktor lingkungan yang terlibat
dalam pertumbuhan. De Wit (1960) memperkenalkan konsep penguasaan sarana
tumbuh yang mencakup semua faktor yang mempengaruhi kompetisi dan
merumuskan pertumbuhan tanaman dalam persamaan berikut :
Keterangan : Y = hasil nyata d = densitas tumbuhan Ymax b = penguasaan sarana/faktor tumbuh (space occupation) atau
kemampuan memanfaatkan sarana tumbuh
= hasil maksimal pada densitas tinggi

16
Persamaan garis tersebut masih bersifat hiperbolik. Resiprokal dari
persamaan tersebut akan memberikan persamaan linear sebagai berikut :
= Angka Penguasaan Sarana Tumbuh ditetapkan dengan rumus : PST = (Y/Ymax) x 100% PST menyatakan besarnya penguasaan sarana tumbuh dan dinyatakan
dalam (%). Nilai PST yang lebih besar menunjukkan dominasi suatu tumbuhan
terhadap tumbuhan lain.
Agresivitas merupakan salah satu pendekatan untuk mengukur
peningkatan hasil relatif dari suatu spesies dalam sistem tumpangsari yang
dikembangkan oleh McGilchrist dan disimbolkan dengan A (Whiley 1979).
Konsep ini didasarkan atas percobaan rangkaian substitusi dan untuk suatu
kombinasi substitusi. Nilai agresivitas dirumuskan sebagai berikut :
Nilai agresivitas sama dengan nol berarti semua spesies dalam pertanaman
campuran mempunyai daya kompetisi yang sama besarnya. Pada sisi lain, nilai
agresivitas mempunyai angka yang sama, tetapi spesies yang dominan memiliki
nilai positif sedangkan spesies yang didominasi memiliki nilai negatif.
Marka Molekuler Simple Sequence Repeat (SSR)
Penanda morfologi telah digunakan untuk mengatasi masalah duplikasi
plasma nutfah di lapang, penentuan jarak genetik dan hubungan kekerabatan antar
plasma nutfah/klon/kultivar (Vuylsteke et al. 1988; Ortiz et al. 1993; Swennen
et al. 1995; Soejono et al. 2001). Namun, penanda morfologi sering menunjukkan
hasil yang bias karena pengaruh lingkungan terhadap penampakan fenotipik.
Untuk memperkuat informasi data penanda morfologi, diperlukan dukungan
penanda molekuler yang memperjelas perbedaan dan hubungan kekerabatan antar

17
aksesi berdasarkan karakteristik molekulernya (DNA) (Jarret dan Gawel 1995).
Penanda molekuler didasarkan pada polimorfisme yang dideteksi pada tingkat
makromolekul dalam sel. Pengertian ini akhir-akhir ini banyak digunakan untuk
mendiskripsikan hanya pada DNA. Penanda DNA ini dapat tidak terbatas dalam
jumlah dan dapat memberikan kegunaan yang besar untuk beragam tujuan yang
relevan terhadap perbaikan tanaman.
Penanda DNA yang memiliki tingkat akurasi cukup tinggi salah satunya
adalah simple sequen repeat (SSR) atau short tandem repeat (STR) atau
mikrosatelit. SSR memiliki keunggulan mudah dan murah (pada tahapan setelah
ditemukan primer spesifiknya), keberadaannya melimpah dan tersebar di seluruh
genom tanaman, dan dengan sampel dalam jumlah sedikit, mencukupi untuk
amplifikasi dengan PCR (Ribaut et al 2002). Salah satu teknik yang
memanfaatkan mikrosatelit adalah Sequence-tagged microsatellite sites (STMSs)
atau Sequence-tagged sites (STS). Keuntungan STMSs adalah menggunakan
sepasang primer yang sudah didisain khusus untuk tiap spesies (Sulyo 1997),
sehingga menyebabkan penanda ini bersifat ko-dominan (Hiu LB 1998;
Sulyo 1997). Penanda STMS memungkinkan mendapat derajat polimorfisme dan
variasi yang tinggi karena sekuen DNA mikrosatelit mengandung urutan basa
berulang-ulang secara bergandengan dengan panjang berbeda-beda pada genom.
Bentuk berulang yang umum adalah dinukleotida sederhana. Frekuensinya yang
tinggi dalam genom lebih mudah dideteksi dibandingkan mikrosatelit dengan
ulangan tri- dan tetranukleotida (Hiu Liu 1998). Mikrosatelit tri- dan
tetranukleotida lebih sedikit dalam genom dan tingkat keragamannya lebih rendah
dari dinukleotida (Scotti et al. 2002). Variasi terjadi dalam ukuran panjang
mikrosatelit pada lokus-lokus individu yang spesifik, sehingga penanda ini
polialelik dan ko-dominan secara alami, yang menjadikan penanda ini mempunyai
manfaat lebih banyak (Puspendra et al. 2002) dan memiliki tingkat
reprodusibilitas tinggi dibandingkan penanda RAPD dan RFLP (McGregor et al.
2000; Powell et al. 1996).
Studi yang membandingkan reprodusibilitas dari beberapa penanda DNA
pada tanaman kedelai menyimpulkan bahwa SSR mempunyai nilai informasi
yang tinggi (tingkat polimorfik, indeks keanekaragaman genetik, tingkat

18
heterosigositas yang diharapkan) dan spesifik lokus, sehingga menjadi penanda
pilihan untuk beragam tujuan termasuk pemuliaan tanaman (Powell et al. 1996).
Pada studi kesamaan dan keanekaragaman genetik anggur, dari data molekuler
yang membandingkan tingkat efektivitas SSR dan AFLP, menunjukkan bahwa
SSR sama efektifnya dengan AFLP (Fossati et al. 2001).

PERTUMBUHAN DAN PRODUKSI PADI PADA BERBAGAI TINGKAT POPULASI GULMA
Echinochloa crus-galli YANG BERASAL DARI TIGA LOKASI
ABSTRAK
E. crus-galli merupakan gulma utama pada pertanaman padi sawah yang dapat menurunkan hasil panen. Tujuan penelitian adalah mempelajari pengaruh aksesi dan populasi gulma E. crus-galli terhadap pertumbuhan dan produksi tanaman padi. Penelitian dilaksanakan di rumah kaca, Kebun Percobaan IPB, Cikabayan, Bogor mulai bulan Mei 2006 hingga September 2006. Penelitian dilakukan dengan menggunakan rancangan petak terbagi dengan tiga ulangan. Aksesi gulma E. crus-galli sebagai petak utama terdiri atas aksesi Karawang, Cikampek, dan Sukabumi. Populasi gulma E. crus-galli sebagai anak petak terdiri atas lima taraf yaitu 0, 1, 2, 3, dan 4 gulma E. crus-galli/pot. Hasil penelitian menunjukkan bahwa aksesi gulma E. crus-galli berpengaruh terhadap tinggi tanaman, jumlah anakan, dan kepadatan malai. Daya kompetisi gulma E. crus-galli aksesi Cikampek lebih tinggi dibandingkan dengan aksesi Sukabumi dan Karawang. Populasi E. crus-galli berpengaruh terhadap terhadap pertumbuhan dan produksi tanaman padi. Populasi 4 gulma E. crus-galli/pot menurunkan berat kering gabah total sebesar 48.0% dan berat kering gabah isi sebesar 46.2%. Interaksi antara aksesi dan populasi gulma E. crus-galli tidak berpengaruh terhadap pertumbuhan dan produksi tanaman padi. Kata kunci : Echinochloa crus-galli, kompetisi, populasi, gulma, padi sawah.

20
GROWTH AND PRODUCTION OF RICE PLANT ON SOME POPULATION DENSITY OF Echinochloa crus-galli WEED
FROM THREE LOCATIONS
ABSTRACT
Echinochloa crus-galli is a major weed in paddy field that reduces rice yield. The objective of the research was to study the effect of E. crus-galli accession and population on rice growth and production. The research was conducted in a green house using split plot design with three replications. The main plot consisted of three E. crus-galli accession i.e. Karawang, Cikampek, and Sukabumi accession. E. crus-galli population as sub plot consisted of 0, 1, 2, 3, and 4 E. crus-galli per pot. The results showed that accession of E. crus-galli affected plant height, number of tiller, and panicle density. The competitiveness against rice of E. crus-galli accession Cikampek was higher than that of Sukabumi dan Karawang accession. Population E. crus-galli affected rice growth and production. Population of 4 E. crus-galli/pot decreased spikelets weigth about 48.0% and filled spikelets weigth about 46.2%. Interaction of accession and population of E. crus-galli did not affected rice growth and production. Key words : Echinochloa crus-galli, competition, population, weed, rice.

21
Pendahuluan
Kebutuhan beras semakin meningkat sejalan dengan peningkatan jumlah
penduduk. Pada tahun 2030 nanti, kebutuhan beras Indonesia diperkirakan
mencapai 41.7 juta ton (BPS 2008). Upaya peningkatan produksi beras pada
masa yang akan datang dihadapkan pada berbagai kendala seperti alih fungsi
lahan pertanian ke non pertanian, degradasi kesuburan lahan, dan serangan
organisme pengganggu tanaman (OPT).
Salah satu OPT yang dapat menurunkan produksi tanaman padi adalah
gulma. Penurunan produksi yang diakibatkan oleh gulma pada beberapa situasi
secara ekonomis lebih penting daripada penurunan produksi yang disebabkan oleh
insekta, cendawan, atau organisme pengganggu lainnya (Savary et al. 1997;
2000). Selain penurunan produksi, adanya gulma di pertanaman padi sawah juga
menyebabkan biaya pengendalian yang besar sehingga menurunkan pendapatan
petani (Tungate et.al. 2007).
Salah satu spesies gulma dominan pada lahan sawah adalah Echinochloa
crus-galli (Ali dan Sankaran, 1984). Kehadiran gulma E. crus-galli di
pertanaman padi sawah dapat menurunkan produksi tanaman padi hingga 50-59%
(Sultana 2000; Chin 2001), 57-95% (Ahn dan Chung 2000), dan bahkan dapat
menurunkan produksi gabah hingga 97% (Islam dan Karim 2003). Penurunan
produksi tersebut disebabkan oleh adanya kompetisi antara gulma dan tanaman
padi terhadap sumberdaya yang tersedia (Zimdahl 2004). Gulma ini juga dapat
menjadi tumbuhan inang bagi Leptocorisa oratorius, Acrocylindricum oryzae,
Corticium sasakii, dan Rhynchosporium oryzae (Tjitrosemito 1994).
Gulma E. crus-galli memiliki daya adaptasi yang luas pada kondisi
lingkungan yang beragam (Galinato et al. 1999). Karena kemampuan adaptasi
yang luas, maka gulma E. crus-galli dari tiap aksesi diduga memiliki daya
kompetisi yang berbeda pula. Perubahan praktek agronomis pada berbagai lokasi
dari waktu ke waktu seperti penggunaan herbisida baru, inovasi cara pengolahan
tanah, penggunaan kultivar baru dapat mempengaruhi distribusi gulma dan
kemampuan kompetisi gulma terhadap tanaman budidaya (Froud-Williams et al.
1984; Clement et al. 1996). Perbedaan karakter daya kompetisi dari aksesi gulma
E. crus-galli dalam menurunkan produksi tanaman padi belum diteliti di

22
Indonesia. Penelitian bertujuan untuk mempelajari pengaruh aksesi gulma E.
crus-galli pada beberapa tingkat populasi terhadap pertumbuhan dan produksi
tanaman padi sawah.
Bahan dan Metode
Penelitian dilakukan dalam pot di rumah kaca Kebun Percobaan Cikabayan,
Dramaga, Bogor, pada bulan Mei 2006 sampai dengan September 2006. Bahan
yang digunakan antara lain benih padi varietas IR-64, pupuk urea, SP-36, dan
KCl, dan biji E. crus-galli. Peralatan yang digunakan antara lain pot berukuran 30
cm - 40 cm (diameter - tinggi), tray, neraca, oven, dan leaf area meter.
Percobaan menggunakan rancangan petak terbagi (split plot design) dalam
rancangan acak kelompok dengan tiga ulangan. Percobaan terdiri atas dua faktor,
yaitu aksesi E. crus-galli sebagai petak utama yang terdiri atas tiga aksesi (aksesi
Karawang, Cikampek, dan Sukabumi) dan populasi E. crus-galli sebagai anak
petak yang terdiri atas lima taraf, yaitu 0, 1, 2, 3, dan 4 bibit gulma E. crus-galli
per pot. Satuan percobaan terdiri atas 3 pot sehingga total terdapat 135 pot
percobaan.
Media tanam yang digunakan adalah tanah latosol dramaga yang berasal
dari lahan sawah kebun percobaan IPB Sawah Baru. Sebelum digunakan sebagai
media, tanah dikeringanginkan terlebih dahulu, kemudian dihaluskan dan diayak.
Tiap pot diisi media tanah sebanyak 10 kg/pot. Media tanah dalam pot
selanjutnya dilumpurkan dan digenangi dengan air hinga ketinggian 5 cm dari
permukaan media. Benih padi varietas IR-64 dan biji E. crus-galli disemai
sebelum penanaman di pot dengan menggunakan bak semai. Bibit padi
dipindahtanam ke dalam pot pada saat berumur 21 hari setelah semai (HSS).
Bibit padi ditanam tepat di tengah-tengah pot. Bibit gulma E. crus-galli yang
berumur 14 HSS ditanam pada jarak 7 cm dari tanaman padi dengan jumlah bibit
sesuai dengan perlakuan.
Pemeliharaan tanaman yang dilakukan meliputi pemupukan, penyiraman,
pengendalian hama penyakit. Pupuk SP-36 dan KCl dengan dosis masing-masing
sebesar 0.5 g/pot diberikan seluruhnya pada saat tanam, sedangkan pupuk urea
dengan dosis 1.5 g/pot diberikan 3 kali yaitu 1/3 dosis pada saat tanam, 1/3 dosis

23
pada 4 minggu setelah tanam (MST), dan 1/3 dosis pada 8 MST. Penyiraman
dilakukan 2 hari sekali sampai ketinggian genangan sekitar 5 cm. Penyiangan
gulma selain E. crus-galli dilakukan secara manual. Pengendalian penyakit
tungro dilakukan dengan cara membuang bagian tanaman yang terserang. Panen
padi dan gulma E. crus-galli dilakukan bersamaan pada 13 MST.
Peubah yang diamati antara lain tinggi tanaman, jumlah anakan, bobot
biomassa tajuk dan akar, panjang dan lebar daun bendera, luas daun bendera,
kadar nitrogen daun bendera, jumlah anakan produktif, panjang malai, kepadatan
malai, dan produksi gabah. Data hasil pengamatan dianalisis menggunakan
analisis ragam (uji F) dengan uji lanjut DMRT (Duncan’s Multiple Range Test)
pada taraf nyata 5%.
Hasil dan Pembahasan
Pertumbuhan Tanaman Padi
Aksesi gulma E. crus-galli berpengaruh terhadap tinggi tanaman padi pada
saat 7 MST (Tabel 1). Tinggi tanaman padi yang ditanam dengan gulma E. crus-
galli aksesi Sukabumi lebih pendek dibandingkan dengan yang ditanam dengan
gulma aksesi lainnya.
Tabel 1. Pengaruh aksesi gulma terhadap tinggi tanaman padi
Aksesi Tinggi Tanaman Padi (cm) 2 MST 3 MST 4 MST 5 MST 6 MST 7 MST 8 MST
Karawang 39.8 54.6 67.0 80.9 89.4 92.4a 95.1 Cikampek 38.7 53.0 66.4 80.2 87.6 91.4a 93.7 Sukabumi 40.2 52.5 67.7 80.4 86.6 89.4b 92.0
Keterangan: angka yang diikuti huruf yang sama pada kolom yang sama, tidak berbeda nyata pada uji DMRT taraf 5%.
Populasi dan interaksi antara aksesi dengan populasi E. crus-galli tidak
berpengaruh terhadap tinggi tanaman padi. Hasil berbeda dilaporkan oleh Islam et
al. (2003) bahwa keberadaan gulma E. crus-galli mulai populasi 2 per pot
menurunkan tinggi tanaman padi. Perera et al. (1992), Sultana (2000) dan Purba
(2007) juga melaporkan adanya penurunan tinggi tanaman padi akibat kompetisi
E. crus-galli.
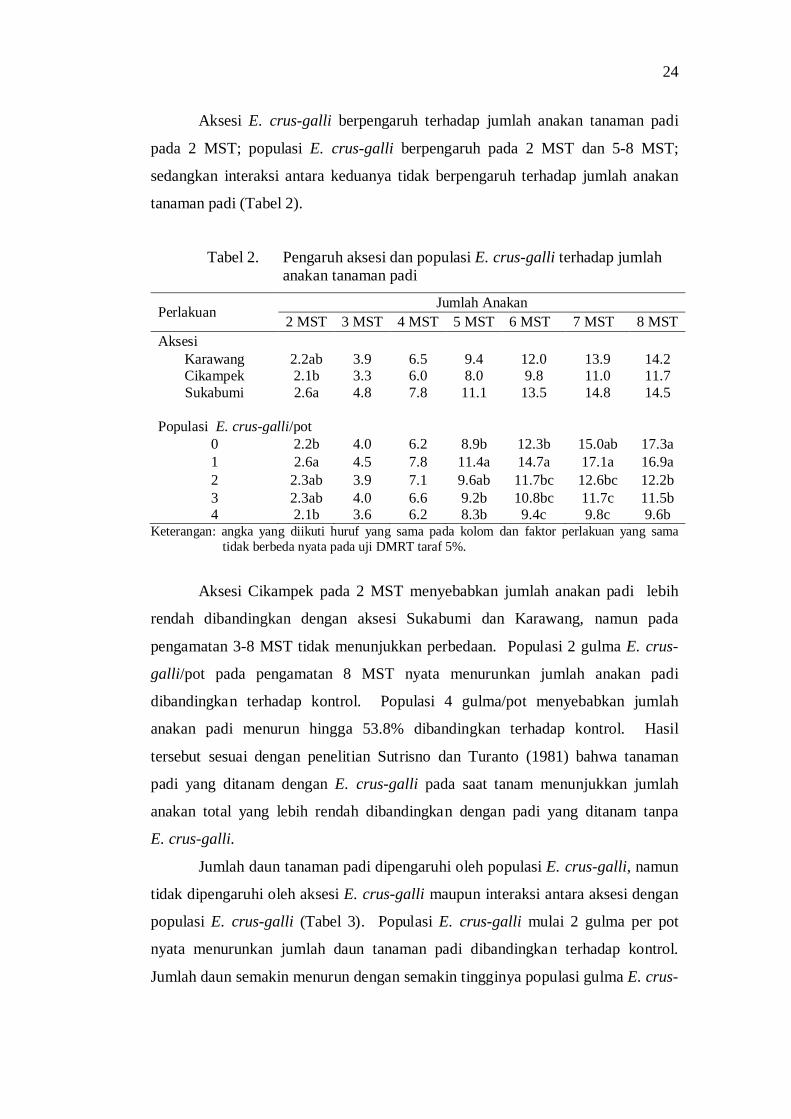
24
Aksesi E. crus-galli berpengaruh terhadap jumlah anakan tanaman padi
pada 2 MST; populasi E. crus-galli berpengaruh pada 2 MST dan 5-8 MST;
sedangkan interaksi antara keduanya tidak berpengaruh terhadap jumlah anakan
tanaman padi (Tabel 2).
Tabel 2. Pengaruh aksesi dan populasi E. crus-galli terhadap jumlah anakan tanaman padi
Perlakuan Jumlah Anakan 2 MST 3 MST 4 MST 5 MST 6 MST 7 MST 8 MST
Aksesi Karawang 2.2ab 3.9 6.5 9.4 12.0 13.9 14.2 Cikampek 2.1b 3.3 6.0 8.0 9.8 11.0 11.7 Sukabumi 2.6a 4.8 7.8 11.1 13.5 14.8 14.5
Populasi E. crus-galli/pot
0 2.2b 4.0 6.2 8.9b 12.3b 15.0ab 17.3a 1 2.6a 4.5 7.8 11.4a 14.7a 17.1a 16.9a 2 2.3ab 3.9 7.1 9.6ab 11.7bc 12.6bc 12.2b 3 2.3ab 4.0 6.6 9.2b 10.8bc 11.7c 11.5b 4 2.1b 3.6 6.2 8.3b 9.4c 9.8c 9.6b
Keterangan: angka yang diikuti huruf yang sama pada kolom dan faktor perlakuan yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Aksesi Cikampek pada 2 MST menyebabkan jumlah anakan padi lebih
rendah dibandingkan dengan aksesi Sukabumi dan Karawang, namun pada
pengamatan 3-8 MST tidak menunjukkan perbedaan. Populasi 2 gulma E. crus-
galli/pot pada pengamatan 8 MST nyata menurunkan jumlah anakan padi
dibandingkan terhadap kontrol. Populasi 4 gulma/pot menyebabkan jumlah
anakan padi menurun hingga 53.8% dibandingkan terhadap kontrol. Hasil
tersebut sesuai dengan penelitian Sutrisno dan Turanto (1981) bahwa tanaman
padi yang ditanam dengan E. crus-galli pada saat tanam menunjukkan jumlah
anakan total yang lebih rendah dibandingkan dengan padi yang ditanam tanpa
E. crus-galli.
Jumlah daun tanaman padi dipengaruhi oleh populasi E. crus-galli, namun
tidak dipengaruhi oleh aksesi E. crus-galli maupun interaksi antara aksesi dengan
populasi E. crus-galli (Tabel 3). Populasi E. crus-galli mulai 2 gulma per pot
nyata menurunkan jumlah daun tanaman padi dibandingkan terhadap kontrol.
Jumlah daun semakin menurun dengan semakin tingginya populasi gulma E. crus-

25
galli. Namun demikian, populasi 4 gulma E. crus-galli menghasilkan jumlah
daun yang sebanding dengan populasi 3 gulma E. crus-galli mulai pengamatan
9 MST sampai dengan 13 MST.
Tabel 3. Jumlah daun tanaman padi pada perlakuan populasi E. crus-galli
Populasi E. crus-galli/pot
Jumlah Daun Tanaman Padi (helai) 9 MST 10 MST 11 MST 12 MST 13 MST
0 78.3a 76.0a 60.4a 56.2a 53.9a 1 79.1a 71.7ab 50.0ab 45.3ab 43.7ab 2 66.1a 59.3b 42.7b 38.2b 36.6b 3 49.9b 42.9c 29.2c 25.4c 24.6c 4 43.1b 35.6c 23.0c 19.7c 18.9c
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Jumlah daun tanaman padi terkait dengan jumlah anakan per rumpun.
Semakin tinggi populasi E. crus-galli jumlah anakan tanaman padi semakin
menurun dan akhirnya menurunkan jumlah daun per rumpun. Penurunan jumlah
anakan dan jumlah daun tanaman padi diduga disebabkan oleh adanya kompetisi
antar spesies antara tanaman padi dengan gulma E. crus-galli dalam
memperebutkan hara. Kekurangan hara P dapat menyebabkan penurunan jumlah
anakan tanaman padi. Selain itu, penurunan jumlah anakan dan jumlah daun
diduga disebabkan oleh adanya zat alelopati yang dikeluarkan oleh gulma E. crus-
galli. Yamamoto et al. (1999) dan Xuan et al. (2006) menyatakan bahwa eksudat
akar E. crus-galli yaitu senyawa hidroxymandelic acid dan lactones,
menyebabkan penurunan perkecambahan dan pertumbuhan tanaman padi.
Bobot kering tajuk tanaman padi pada 2 bulan setelah tanam (BST) dan
3 BST dipengaruhi oleh populasi E. crus-galli. Aksesi E. crus-galli maupun
interaksi antara aksesi dengan populasi E. crus-galli tidak berpengaruh terhadap
bobot kering tajuk tanaman padi (Tabel 4). Gulma E. crus-galli pada pengamatan
3 BST menurunkan bobot kering tajuk tanaman padi mulai populasi 2 gulma/pot
dengan penurunan sebesar 30.9% dibandingkan terhadap kontrol. Eussen dan
Zulfadli (1981) menyatakan bahwa produksi bahan kering tanaman menurun
akibat ditanam bersama dengan gulma pada seluruh siklus pertumbuhannya.
Penurunan bobot kering tersebut diduga terjadi akibat adanya kompetisi antara
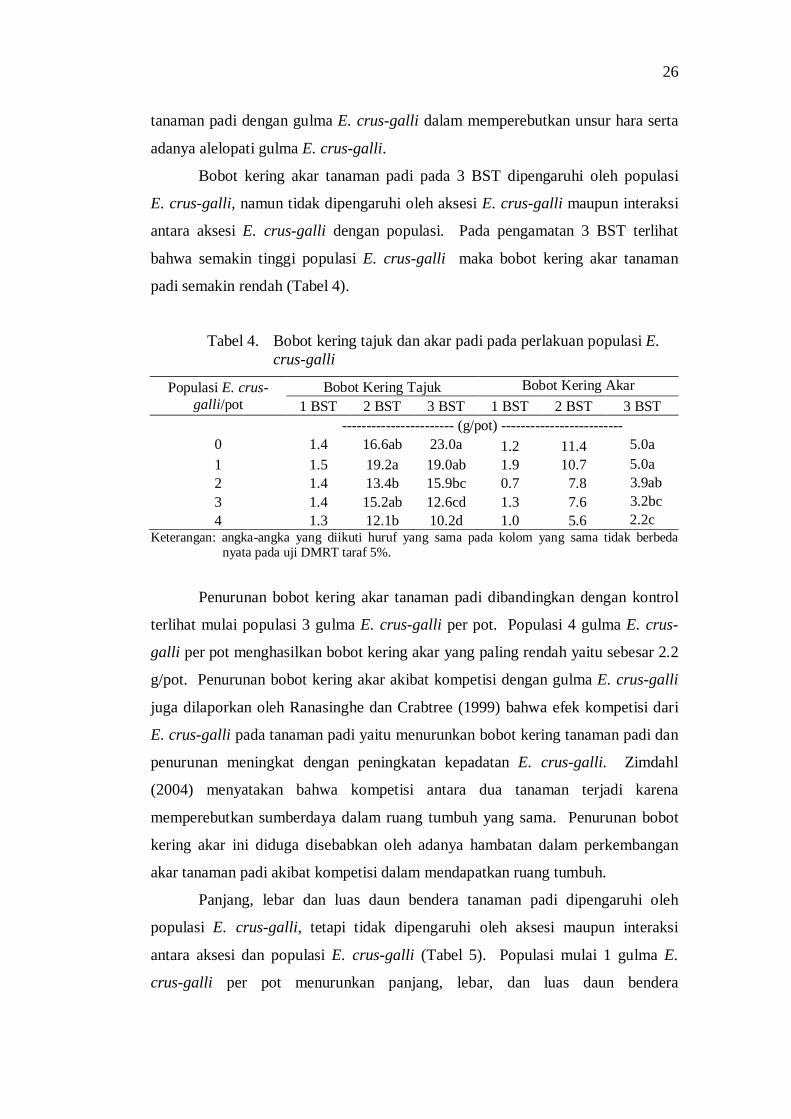
26
tanaman padi dengan gulma E. crus-galli dalam memperebutkan unsur hara serta
adanya alelopati gulma E. crus-galli.
Bobot kering akar tanaman padi pada 3 BST dipengaruhi oleh populasi
E. crus-galli, namun tidak dipengaruhi oleh aksesi E. crus-galli maupun interaksi
antara aksesi E. crus-galli dengan populasi. Pada pengamatan 3 BST terlihat
bahwa semakin tinggi populasi E. crus-galli maka bobot kering akar tanaman
padi semakin rendah (Tabel 4).
Tabel 4. Bobot kering tajuk dan akar padi pada perlakuan populasi E. crus-galli
Populasi E. crus-galli/pot
Bobot Kering Tajuk Bobot Kering Akar 1 BST 2 BST 3 BST 1 BST 2 BST 3 BST
----------------------- (g/pot) ------------------------- 0 1.4 16.6ab 23.0a 1.2 11.4 5.0a 1 1.5 19.2a 19.0ab 1.9 10.7 5.0a 2 1.4 13.4b 15.9bc 0.7 7.8 3.9ab 3 1.4 15.2ab 12.6cd 1.3 7.6 3.2bc 4 1.3 12.1b 10.2d 1.0 5.6 2.2c
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Penurunan bobot kering akar tanaman padi dibandingkan dengan kontrol
terlihat mulai populasi 3 gulma E. crus-galli per pot. Populasi 4 gulma E. crus-
galli per pot menghasilkan bobot kering akar yang paling rendah yaitu sebesar 2.2
g/pot. Penurunan bobot kering akar akibat kompetisi dengan gulma E. crus-galli
juga dilaporkan oleh Ranasinghe dan Crabtree (1999) bahwa efek kompetisi dari
E. crus-galli pada tanaman padi yaitu menurunkan bobot kering tanaman padi dan
penurunan meningkat dengan peningkatan kepadatan E. crus-galli. Zimdahl
(2004) menyatakan bahwa kompetisi antara dua tanaman terjadi karena
memperebutkan sumberdaya dalam ruang tumbuh yang sama. Penurunan bobot
kering akar ini diduga disebabkan oleh adanya hambatan dalam perkembangan
akar tanaman padi akibat kompetisi dalam mendapatkan ruang tumbuh.
Panjang, lebar dan luas daun bendera tanaman padi dipengaruhi oleh
populasi E. crus-galli, tetapi tidak dipengaruhi oleh aksesi maupun interaksi
antara aksesi dan populasi E. crus-galli (Tabel 5). Populasi mulai 1 gulma E.
crus-galli per pot menurunkan panjang, lebar, dan luas daun bendera
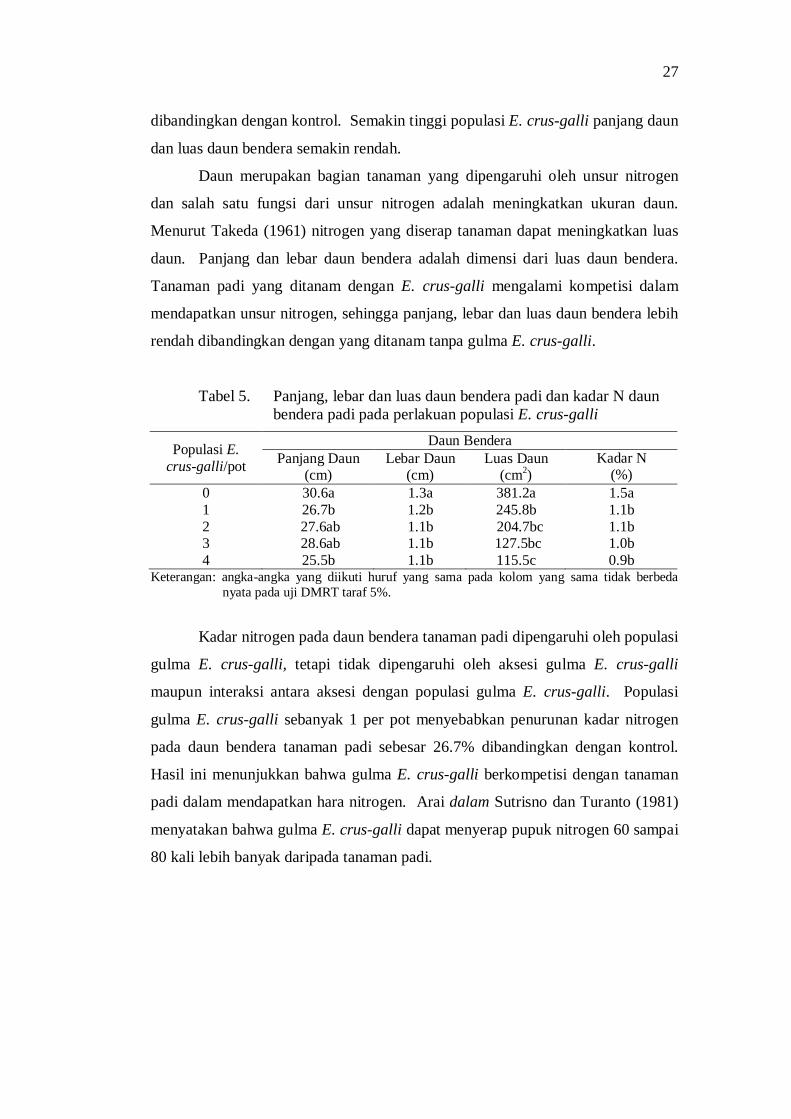
27
dibandingkan dengan kontrol. Semakin tinggi populasi E. crus-galli panjang daun
dan luas daun bendera semakin rendah.
Daun merupakan bagian tanaman yang dipengaruhi oleh unsur nitrogen
dan salah satu fungsi dari unsur nitrogen adalah meningkatkan ukuran daun.
Menurut Takeda (1961) nitrogen yang diserap tanaman dapat meningkatkan luas
daun. Panjang dan lebar daun bendera adalah dimensi dari luas daun bendera.
Tanaman padi yang ditanam dengan E. crus-galli mengalami kompetisi dalam
mendapatkan unsur nitrogen, sehingga panjang, lebar dan luas daun bendera lebih
rendah dibandingkan dengan yang ditanam tanpa gulma E. crus-galli.
Tabel 5. Panjang, lebar dan luas daun bendera padi dan kadar N daun bendera padi pada perlakuan populasi E. crus-galli
Populasi E. crus-galli/pot
Daun Bendera Panjang Daun
(cm) Lebar Daun
(cm) Luas Daun
(cm2Kadar N
) (%) 0 30.6a 1.3a 381.2a 1.5a 1 26.7b 1.2b 245.8b 1.1b 2 27.6ab 1.1b 204.7bc 1.1b 3 28.6ab 1.1b 127.5bc 1.0b 4 25.5b 1.1b 115.5c 0.9b
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Kadar nitrogen pada daun bendera tanaman padi dipengaruhi oleh populasi
gulma E. crus-galli, tetapi tidak dipengaruhi oleh aksesi gulma E. crus-galli
maupun interaksi antara aksesi dengan populasi gulma E. crus-galli. Populasi
gulma E. crus-galli sebanyak 1 per pot menyebabkan penurunan kadar nitrogen
pada daun bendera tanaman padi sebesar 26.7% dibandingkan dengan kontrol.
Hasil ini menunjukkan bahwa gulma E. crus-galli berkompetisi dengan tanaman
padi dalam mendapatkan hara nitrogen. Arai dalam Sutrisno dan Turanto (1981)
menyatakan bahwa gulma E. crus-galli dapat menyerap pupuk nitrogen 60 sampai
80 kali lebih banyak daripada tanaman padi.
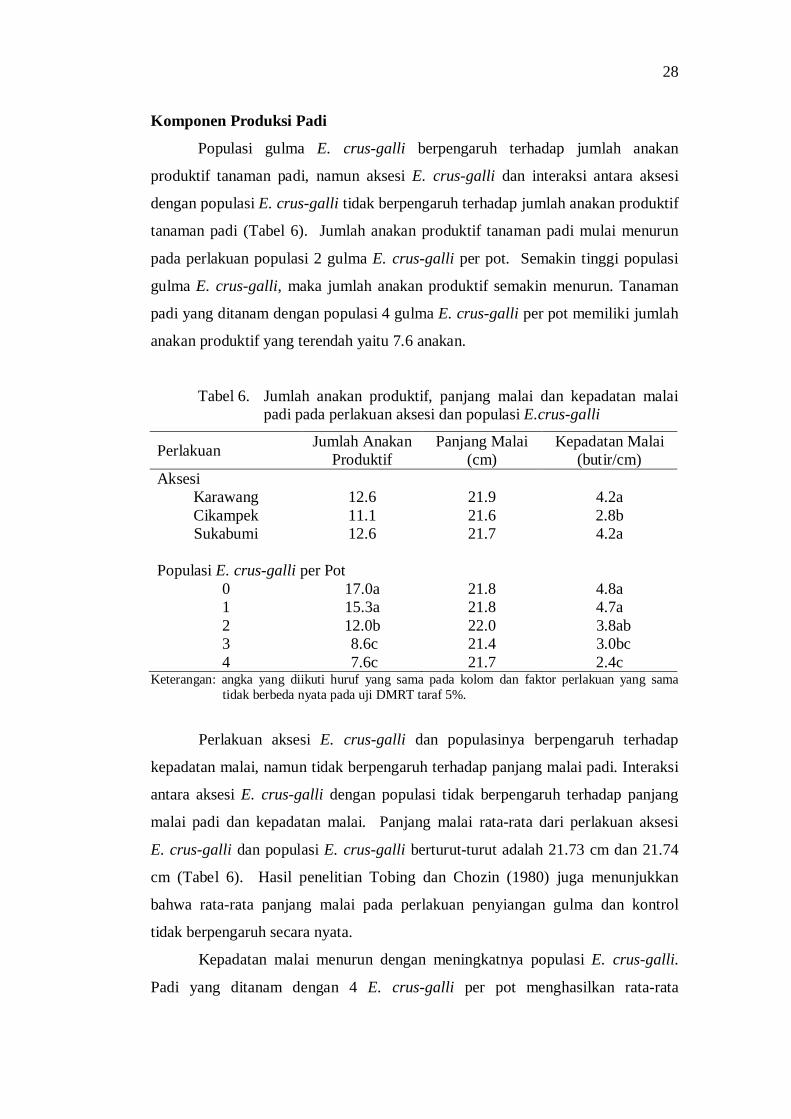
28
Komponen Produksi Padi
Populasi gulma E. crus-galli berpengaruh terhadap jumlah anakan
produktif tanaman padi, namun aksesi E. crus-galli dan interaksi antara aksesi
dengan populasi E. crus-galli tidak berpengaruh terhadap jumlah anakan produktif
tanaman padi (Tabel 6). Jumlah anakan produktif tanaman padi mulai menurun
pada perlakuan populasi 2 gulma E. crus-galli per pot. Semakin tinggi populasi
gulma E. crus-galli, maka jumlah anakan produktif semakin menurun. Tanaman
padi yang ditanam dengan populasi 4 gulma E. crus-galli per pot memiliki jumlah
anakan produktif yang terendah yaitu 7.6 anakan.
Tabel 6. Jumlah anakan produktif, panjang malai dan kepadatan malai padi pada perlakuan aksesi dan populasi E.crus-galli
Perlakuan Jumlah Anakan Produktif
Panjang Malai (cm)
Kepadatan Malai (butir/cm)
Aksesi Karawang 12.6 21.9 4.2a Cikampek 11.1 21.6 2.8b Sukabumi 12.6 21.7 4.2a
Populasi E. crus-galli per Pot
0 17.0a 21.8 4.8a 1 15.3a 21.8 4.7a 2 12.0b 22.0 3.8ab 3 8.6c 21.4 3.0bc 4 7.6c 21.7 2.4c
Keterangan: angka yang diikuti huruf yang sama pada kolom dan faktor perlakuan yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Perlakuan aksesi E. crus-galli dan populasinya berpengaruh terhadap
kepadatan malai, namun tidak berpengaruh terhadap panjang malai padi. Interaksi
antara aksesi E. crus-galli dengan populasi tidak berpengaruh terhadap panjang
malai padi dan kepadatan malai. Panjang malai rata-rata dari perlakuan aksesi
E. crus-galli dan populasi E. crus-galli berturut-turut adalah 21.73 cm dan 21.74
cm (Tabel 6). Hasil penelitian Tobing dan Chozin (1980) juga menunjukkan
bahwa rata-rata panjang malai pada perlakuan penyiangan gulma dan kontrol
tidak berpengaruh secara nyata.
Kepadatan malai menurun dengan meningkatnya populasi E. crus-galli.
Padi yang ditanam dengan 4 E. crus-galli per pot menghasilkan rata-rata

29
kepadatan malai terendah yaitu 2.4 butir/cm. Penurunan kepadatan malai pada
populasi 4 E. crus-galli per pot sebesar 50.1% dibandingkan terhadap kontrol
(Tabel 6). Gulma E. crus-galli aksesi Cikampek memiliki daya kompetisi yang
lebih besar dalam menurunkan kepadatan malai yang ditunjukkan dengan
kepadatan malai yang lebih rendah dibandingkan dengan aksesi Karawang dan
Sukabumi (Tabel 6). Daya kompetisi yang lebih besar ini diduga sebagai akibat
praktek budidaya tanaman padi sawah di lokasi Cikampek yang dinamis
dibandingkan dengan aksesi Sukabumi dan aksesi Karawang. Menurut Froud-
Williams et al. (1984) dan Clement et al. (1996) praktik agronomi tanaman yang
tidak statis dalam waktu dan ruang seperti penggunaan herbisida kelas baru,
kultivar, inovasi pengolahan tanah, penggunaan irigasi, dapat mempengaruhi
distribusi geografis gulma dan daya kompetisi gulma dalam menurunkan
produksi.
Produksi Gabah
Populasi gulma E. crus-galli berpengaruh terhadap bobot gabah total,
bobot gabah isi, bobot gabah hampa, dan persentase gabah hampa. Aksesi gulma
tidak berpengaruh terhadap bobot gabah total, bobot gabah isi, bobot gabah
hampa, dan persentase gabah hampa (Tabel 7).
Tabel 7. Pengaruh aksesi dan populasi E. crus-galli terhadap bobot gabah dan persentase kehampaan
Perlakuan Bobot Gabah (g/pot) Persen Hampa (% w/w) Isi Hampa Total
Aksesi Karawang 16.1 1.4 17.5 7.8 Cikampek 12.1 1.0 13.1 8.0 Sukabumi 17.2 1.4 18.6 7.1
Populasi per Pot
0 18.4a 2.0a 20.4a 10.1a 1 18.8a 1.8a 20.7a 9.8ab 2 15.9ab 1.1b 17.0ab 6.8abc 3 12.6ab 0.7b 13.3b 5.3c 4 9.9b 0.7b 10.6b 6.2bc
Keterangan: angka yang diikuti huruf yang sama pada kolom dan faktor perlakuan yang sama tidak berbeda nyata pada uji DMRT taraf 5%.

30
Populasi gulma E. crus-galli sebanyak 4 gulma per pot menurunkan bobot
gabah isi sebesar 46.2% dan menurunkan bobot gabah total sebesar 48.0%
dibandingkan terhadap kontrol. Namun demikian, persen kehampaan pada
populasi gulma E. crus-galli sebanyak 4 gulma per pot menurun dibandingkan
dengan tanpa gulma. Bobot gabah isi yang rendah diduga disebabkan oleh adanya
kompetisi dalam mendapatkan unsur nitrogen. Menurut De Data (1981) salah satu
fungsi nitrogen pada tanaman padi adalah meningkatkan jumlah gabah isi.
Kesimpulan
Perbedaan aksesi gulma E. crus-galli menyebabkan perbedaan
pertumbuhan dan produksi tanaman padi khususnya pada peubah tinggi tanaman
pada 7 MST, jumlah anakan pada 2 MST, dan kepadatan malai pada saat panen.
Gulma E. crus-galli aksesi Cikampek menunjukkan daya kompetisi yang lebih
kuat dibandingkan aksesi Karawang dan Sukabumi berdasarkan penurunan jumlah
anakan pada 2 MST dan penurunan kepadatan malai pada saat panen.
Kepadatan polulasi gulma E. crus-galli per pot menentukan tingkat
pertumbuhan dan produksi tanaman padi sawah. Semakin tinggi populasi gulma
E. crus-galli pengaruh kompetisi terhadap tanaman padi semakin besar. Populasi
gulma E. crus-galli sebanyak 4 per pot menurunkan bobot gabah sebesar 48.0%
dan menurunkan bobot gabah isi sebesar 46.2%.
Penelitian lanjutan tentang keragaman morfologi dan genetik aksesi gulma
E. crus-galli dari berbagai lokasi di Jawa Barat penting dilakukan untuk
mengetahui apakah keragaman aksesi gulma disebabkan oleh perbedaan geografis
ataukah disebabkan oleh perbedaan genetik.

KERAGAMAN MORFOLOGI DAN GENETIK AKSESI GULMA Echinochloa crus-galli (L.) Beauv.
ASAL JAWA BARAT
ABSTRAK
Gulma E. crus-galli merupakan gulma dominan pada tanaman padi sawah yang menunjukkan variasi morfologi dan genetik. Penelitian bertujuan untuk menganalisis keragaman morfologi dan genetik aksesi gulma E. crus-galli dari berbagai lokasi di Jawa Barat. Sebanyak 16 aksesi E. crus-galli dikoleksi dari tujuh kabupaten di Jawa Barat. Hasil penelitian menunjukkan bahwa aksesi gulma E. crus-galli asal Jawa Barat menunjukkan adanya keragaman morfologi dan genetik. Aksesi gulma E. crus-galli menunjukkan keragaman morfologi baik pada habitat asal maupun pada kondisi rumah kaca. Berdasarkan marka molekuler SSR, aksesi gulma E. crus-galli dapat dikelompokkan menjadi empat sub grup, dimana hampir seluruh aksesi asal Jawa Barat mengelompok ke dalam sub grup A dan C. Pusat keragaman aksesi gulma E. crus-galli diduga berasal dari Subang, Karawang, dan Pangalengan. Implikasi penelitian adalah bahwa tindakan agronomi terpadu harus dilakukan agar penyebaran aksesi gulma dapat dikendalikan. Kata kunci : Echinochloa crus-galli, morfologi, genetik, marka molekuler SSR

32
MORPHOLOGY AND GENETICAL DIVERSITY OF Echinochloa crus-galli (l.) Beauv. WEED ACCESSION
FROM WEST JAVA
ABSTRACT
E. crus-galli is a major weed in the paddy field in Indonesia. This weed shows morphology and genetical variations. The objective of the research was to analyze the morphology and genetic diversity of E. crus-galli accession from some locations in West Java, Indonesia. Genetic diversity of E. crus-galli accession analyze by using SSR markers. The E. crus-galli was collected from 7 district in West Java, Indonesia. The results showed that genetic variations were observed among districts and within district. E. crus-galli accession showed morphology diversity both in origin habitat and in greenhouse. Based on SSR marker, accessions could be grouped into four sub groups, where sub group A and C consisted the most accessions from West Java. It is likely that center of diversity of E. crus-galli is located in Subang, Karawang and Pangalengan. This experiment implies that integrated agronomic treatments should be conducted in order to control distribution of E. crus-galli in West Java. Key word : Echinochloa crus-galli, morphology, genetic, SSR marker.

33
Pendahuluan
Echinochloa crus-galli (L.) Beauv. merupakan gulma semusim yang
memiliki kemampuan adaptasi ekologi yang luas dan daya kompetisi yang kuat
sehingga menjadi spesies gulma penting dan dominan pada tanaman padi sawah
(Altop et al. 2011). Gulma E. crus-galli (L.) Beauv. dapat menyebabkan
kehilangan hasil produksi tanaman padi sawah (Ali dan Sankaran 1984; Ali 1985;
Gealy et al. 2003; Haefele et al. 2004) dengan penurunan hasil produksi padi
mencapai 46-59% (Sultana 2000; Chin 2001; Guntoro et al. 2009), 57-95% (Ahn
dan Chung 2000), dan bahkan mencapai 97% (Islam dan Karim 2003).
Gulma E. crus-galli (L.) Beauv. memiliki distribusi yang luas, mampu
beradaptasi pada berbagai ekologi, toleran terhadap kondisi iklim kering dan
kondisi anaerob, memiliki kemampuan mimikri, perkecambahan dan pertumbuhan
yang cepat, produksi biji yang banyak, sehingga spesies ini menjadi gulma di
lebih dari 60 negara (Barret 1983; Altop et al. 2011). Aoki dan Yamaguchi
(2008) juga melaporkan bahwa gulma E. crus-galli memperlihatkan keragaman
yang sangat tinggi dalam morfologi dan kemampuan beradaptasi pada kondisi
lingkungan yang beragam. Tasrif et al. (2004) melaporkan adanya keragaman
karakter tinggi gulma, panjang malai, luas daun, jumlah biji per malai, panjang
bulu, dan karakter fenotipik lainnya dari E. crus-galli yang disebabkan oleh
perbedaan geografi lokasi aksesi. Menurut Altop et al. (2011), perubahan
morfologi seperti perubahan susunan kanopi akan mempengaruhi kemampuan
kompetisi spesies. Oleh karena itu, informasi tingkat keragaman morfologi dalam
spesies sangat penting untuk menentukan strategi pengendalian yang efektif.
Identifikasi menggunakan karakter morfologi merupakan salah satu upaya
untuk mendapatkan informasi mengenai keragaman genetik. Penanda morfologi
telah digunakan untuk mengatasi masalah duplikasi plasma nutfah di lapang,
penentuan jarak genetik dan hubungan kekerabatan antar plasma
nutfah/klon/kultivar (Vuylsteke et al. 1988; Ortiz et al. 1993; Swennen et al.
1995; Soejono et al. 2001). Untuk memperkuat informasi data
penanda morfologi, diperlukan dukungan penanda molekuler (Jarret dan Gawel
1995). Penggunaan teknik molekuler untuk mengetahui keragaman genetik aksesi
sangat penting, selain untuk mengetahui kemampuan gulma tersebut dalam

34
beradaptasi dengan lingkungannya, juga untuk merencanakan strategi
pengendalian yang tepat (Nissen et al. 1995).
Biologi molekuler saat ini memainkan peranan penting dalam studi
ekologi. Dengan berkembangnya teknik molekuler, marka molekuler yang lebih
beragam telah tersedia untuk menginvestigasi genetik ekologi gulma (Moodie
et al. 1997; Ash et al. 2003; Altop et al. 2011). Metode berdasarkan polymerase
chain reaction (PCR), seperti random amplified polymorphic DNA (RAPD)
(Williams et al. 1990) dan inter simple sequence repeats (ISSR) (Zietkewicz et al.
1994) telah banyak diaplikasikan untuk survey keanekaragaman populasi genetik.
Simple sequence repeat (SSR) atau short tandem repeat (STR) atau mikrosatelit
diyakini memiliki tingkat akurasi yang lebih tinggi dibandingkan dengan penanda
molekuler yang lain. SSR digunakan sebagai penanda karena relatif lebih mudah
dan murah, keberadaannya tersebar di seluruh genom tanaman, dan sampel yang
diperlukan lebih sedikit (Ribaut et al. 2002).
Studi variasi genetik gulma E. crus-galli di Indonesia dengan
menggunakan marka molekuler RAPD telah dilakukan oleh Tasrief et al. (2004).
Namun demikian, informasi tentang keragaman morfologi dan genetik antar
aksesi gulma E. crus-galli dari berbagai lokasi masih sangat terbatas. Penelitian
bertujuan untuk menganalisis keragaman morfologi dan genetik aksesi gulma
E. crus-galli dari berbagai lokasi di Jawa Barat.
Metode Penelitian
Evaluasi Keragaman Morfologi Aksesi E. crus-galli
Evaluasi dilakukan di rumah kaca Kebun Percobaan IPB Darmaga, Bogor
yang berada pada ketinggian 250 m dpl. Sebanyak 16 aksesi E. crus-galli
diperoleh dengan cara mengumpulkan biji-biji gulma yang telah matang dari
berbagai habitat sawah di Jawa Barat yaitu Indramayu, Subang, Cikampek,
Karawang, Sukabumi, Cianjur, dan Pangalengan-Bandung. Koleksi biji gulma
dilakukan pada bulan Maret – April 2007. Lokasi pengambilan contoh aksesi
ditetapkan berdasarkan ketinggian tempat (Tabel 8). Selanjutnya, biji disemai pada
bak semai dan pada saat 14 hari setelah semai bibit gulma dipindahtanam ke dalam
pot plastik berukuran 30-40 (diameter-tinggi pot) dengan media tanah latosol

35
Darmaga sebanyak 9 kg/pot. Setiap aksesi ditanam pada tiga pot. Pemeliharaan
gulma di dalam pot meliputi pemupukan dengan dosis pupuk 0.5 g N/pot, 0.5 g
P2O5/pot, dan 0.5 g K2
O/pot, dan penyiraman dilakukan tiap pagi hari.
Tabel 8. Deskripsi asal-usul 16 aksesi gulma E. crus-galli di Jawa Barat
Kode Aksesi Lokasi
Aksesi
Ketinggian Tempat (m dpl)
Habitat Koordinat Lokasi S E
K3 Karawang 37 Sawah 06˚ 17' 89" 107˚ 20' 101" K5 Karawang 37 Sawah 06˚ 17' 96" 107˚ 20' 178" K6 Karawang 37 Sawah 06˚ 17' 62" 107˚ 20' 118" K7 Karawang 37 Sawah 06˚ 17' 53" 107˚ 20' 150" K9 Karawang 27 Sawah 06˚ 17' 45" 107˚ 20' 70" C4 Cikampek 40 Sawah 06˚ 23' 138" 107˚ 26' 33" S3 Subang 29 Sawah 06˚ 22' 79" 107˚ 35' 126" I1 Indramayu 16 Sawah 06˚ 22' 07" 108˚ 18' 01" I2 Indramayu 16 Sawah 06˚ 22' 48" 108˚ 18' 23" I5 Indramayu 16 Sawah 06˚ 22' 63" 108˚ 18' 12"
Ta1 Cianjur 261 Sawah 06° 48' 797" 107˚ 14' 161" Tc1 Sukabumi 749 Sawah 06° 54' 381" 106˚ 58' 865" Td2 Cianjur 1031 Sawah 06˚ 59' 778" 107˚ 08' 295" Td3 Cianjur 1029 Sawah 06˚ 59' 778" 107˚ 08' 292" Te3 Pangalengan 1242 Sawah 07° 09' 145" 107˚ 33' 308" Tf3 Pangalengan 1473 Sawah 07° 07' 892" 107˚ 33' 371"
Karakter yang diamati pada habitat asal antara lain tinggi gulma, jumlah
anakan, jumlah daun, panjang dan lebar daun, serta panjang malai. Karakter yang
diamati pada kondisi rumah kaca antara lain karakter vegetatif meliput i tinggi
gulma, jumlah anakan, jumlah daun, panjang daun, lebar daun, sudut daun, dan
diameter batang. Karakter panjang daun, lebar daun, dan sudut daun diamati dari
daun ketiga dari atas yaitu daun yang pertama kali membuka sempurna. Karakter
vegetatif diamati pada 10 minggu setelah tanam.
Karakter generatif yang diamati antara lain umur berbunga, umur panen,
panjang malai, jumlah biji per malai, bobot 100 butir biji, panjang biji, diamater
biji, panjang daun bendera, lebar daun bendera, luas daun bendera. Karakter
generatif diamati pada saat panen, kecuali karakter umur berbunga yang diamati
pada saat gulma berbunga. Data morfologi dianalisis dengan cluster analysis dan
analisis komponen utama.

36
Evaluasi Keragaman Genetik Aksesi E. crus-galli
Evaluasi keragaman genetik 16 aksesi gulma E. crus-galli asal Jawa Barat
dilakukan dengan menggunakan marka molekuler Simple Sequence Repeat (SSR).
Primer SSR yang digunakan untuk evaluasi keragaman genetik adalah primer yang
dikembangkan oleh Danquah et al. (2002), yaitu EC01, EC02, EC03, EC04, dan
EC05 (Tabel 9).
Tabel 9. Primer Simple Sequence Repeat (SSR) dan sekuen nukleotida
yang digunakan dalam analisis genetik aksesi E. crus-galli
Lokus GenBank Accession no. Sekuen 5’ to 3’ Repeat Motif
EC1 AY050530 F:ATTACTGGTCAGACGGAAAC (CA)6 R:GCAGTTATCTCCGTGGGCAC EC2 AY050531 F:GGCTCCAAACAAGGCAATTC (CA)
5 R:TTCAGGGAATTTAGTACAAG
EC3 AY050532 F:GAAAGGAAATGGGTTGGCTG (CA)8(TA)3(CA)
12 R:CTTCGCACCATGATCTTCTC
EC4 AY050533 F:AGTAGAAGGCTGCAAGAAGG (GA)4AGAG(GA)
3 R:TCTCAGCCCACTTTGTATAG
EC5 AY050534 F:CAGAGCCTTCAATCATGGTG (CA)
6 R:TGCTTCAAGTTCTAGGAGAC
Bahan-bahan yang digunakan meliputi : buffer ekstraksi (10% CTAB;
0.5 M EDTA (pH 8.0); 1 M Tris-HCl (pH 8.0), 5 M NaCl; 1% β-mercaptoethanol),
buffer purifikasi/buffer CIA (Chloroform : Isoamil Alcohol = 24:1 v/v),
2-propanol, TE (1 M Tris-HCl (pH 8.0); 0.5 M EDTA (pH 8.0)), agarose,
ethidium bromida 1%, Tris-HCL (pH 9.0), MgCl2, dNTPs (1:1:1:1), 1 unit enzim
taq DNA polymerase, dan air bebas ion. Peralatan yang digunakan antara lain
tabung mikro steril (1.5 ml), vorteks, waterbath, sentrifuse, microtube steril,
vacum, bak elektroforesis, lampu UV, mesin PCR, dan Kamera Polaroid.
Isolasi DNA dilaksanakan di laboratorium Departemen Konservasi
Sumberdaya Hutan, Fakultas Kehutanan, IPB. Isolasi DNA dilakukan
berdasarkan metode Doyle & Doyle (1987) yang telah dimodifikasi oleh Sobir
(2000), metode Gawel & Jarret (1991) yang telah dimodifikasi oleh Crouch
et al. (1998a), metode Orozco-Castillo (1995) yang telah dimodifikasi oleh
Toruan-Mathius dan Hutabarat (1997) dan metode Nguyen (2002) yang
dimodifikasi. Daun muda segar (baru terbuka atau masih menggulung) gulma E.

37
crus-galli dipotong dan diambil sekitar 0.2-0.5 g, kemudian ditumbuk dalam
tabung mikro steril (1.5 ml) berisi 0.6-0.8 ml buffer ekstraksi (10% CTAB; 0.5 M
EDTA (pH 8.0); 1 M Tris-HCl (pH 8.0), 5 M NaCl; 1% β-mercaptoethanol), lalu
ditutup rapat, kemudian divorteks agar homogen. Campuran selanjutnya
diinkubasi di dalam waterbath pada suhu 65o
Pemurnian DNA dilakukan dengan penambahan 0.6-0.7 ml buffer
purifikasi/buffer CIA (Chloroform : Isoamil Alcohol = 24:1 v/v), dan pemisahan
fraksi di dalam campuran dilakukan dengan sentrifugasi 13000 rpm selama
10 menit. Fase cair (supernatan) yang diperoleh dipindahkan ke microtube steril
yang baru, lalu ditambahkan 500-600 µl 2-propanol dingin, diinkubasi 4
C selama 15 menit, lalu dikocok.
o
Pengujian kuantitas dan kualitas DNA dilakukan dengan menggunakan
metode Doyle & Doyle (1987) dengan beberapa modifikasi (Sobir 2000). DNA
hasil ekstraksi sebanyak 5 µl ditambah dengan loading dye sebanyak 1 µl
dimasukkan pada sumur agarose gel 1.2% pada bak elektroforesis selama
45 menit pada tegangan 110 volt. Hasil elektroforesis diwarnai dengan ethidium
bromida 1% dan dibilas dengan aquades, selanjutnya pita DNA hasil dilihat pada
lampu UV.
C
selama 30 menit, lalu disentrifuse 13 000 rpm selama 15 menit. Fase cair dibuang
dan fase padat/pelet dikeringkan menggunakan vacum selama 5 menit.
Selanjutnya pelet dilarutkan dalam 100 µl TE (1 M Tris-HCl (pH 8.0); 0.5 M
EDTA (pH 8.0); Aquades).
Reaksi PCR meliputi 50 ng DNA template, primer forward dan reverse
masing-masing 1.2 µM, 10 mM Tris-HCL (pH 9.0), 2.5 mM MgCl2, 0.2 mM
atau 200 µM dNTPs (1:1:1:1) gabungan dari dATP, dCTP, dGTP dan dTTP,
1 unit enzim taq DNA polymerase, dan ditambahkan air bebas ion hingga volume
total 25 µl dimasukkan ke dalam tabung PCR dan diamplifikasi pada mesin ABI
Prism 270.
PCR reaksi diatur denaturasi selama 4 menit pada 94oC, diikuti amplifikasi
sebanyak 30 siklus yaitu 1 menit denaturasi pada 94oC, annealing selama 1 menit
sesuai suhu masing-masing primer dan elongasi selama 45 detik pada 72oC dan
diakhiri 4 oC. Hasil amplifikasi dilihat dengan menggunakan gel agarose 1.4%
dalam elektroforesis yang telah diberi pewarna ethidium bromide 0.5 µg/ml.

38
DNA hasil amplifikasi dicampur dengan loading dye dengan perbandingan DNA :
loading dye adalah 10: 2, selanjutnya dielektroforesis (di-running) pada 50 V
selama 3 jam. Gel hasil elektroforesis direndam dalam 1% ethidium bromide
selama ± 30 menit, kemudian divisualisasikan menggunakan transimulator UV
(312 nm) dan didokumentasikan dengan Kamera Polaroid. Profil pita DNA hasil
amplifikasi pada laju elektroforesis yang sama untuk setiap tanaman diskor nilai
nol (0) jika tidak ada pita dan satu (1) jika ada pita pada posisi yang sama individu
yang dibandingkan. Kesamaan genetik antar dua individu dihitung menurut
koefisien Dice (Rohlf 1998). Analisis pengelompokan dilakukan berdasarkan
kesamaan genetik dan disajikan dalam bentuk dendrogram. Data SSR dianalisis
dengan menggunakan program NTSYS-pc versi 2.0.
Hasil dan Pembahasan
Karakteristik Lingkungan Habitat Asal Aksesi Gulma E. crus-galli
Kondisi iklim berbeda antar habitat asal gulma E. crus-galli. Kondisi
iklim dapat mempengaruhi distribusi aksesi gulma E. crus-galli. Suhu udara rata-
rata bervariasi antara 19 - 34 °C, curah hujan tahunan rata-rata bervariasi antara
1000 - 4000 mm/tahun, dan kelembaban udara bervariasi antara 70-91%. Jenis
tanah habitat asal aksesi gulma relatif sama antar lokasi yaitu aluvial. Ketinggian
tempat berbeda mulai 16 m dpl hingga 1473 m dpl (Tabel 10).
Tabel 10. Karakteristik lingkungan dari habitat asal aksesi gulma E. crus-galli dan lokasi penanaman gulma E. crus-galli
Aksesi Suhu Udara (°C)
Curah Hujan Tahunan
(mm/tahun)
Kelembaban Udara (%)
Jenis Tanah
Indramayu 25-34 1430 70-80 Aluvial Subang 30-33 1600-1900 72-91 Aluvial Karawang 24-28 1100-3200 80 Aluvial Cikampek 24-33 1000-2000 87 Aluvial Cianjur 23-30 1000-4000 85-89 Aluvial Sukabumi 20-30 3000-4000 85-89 Aluvial Bandung 19-24 1500-4000 78 Aluvial
Sumber: http://www.weather.yahoo.com dan http://www.jabar.go.id [2010].
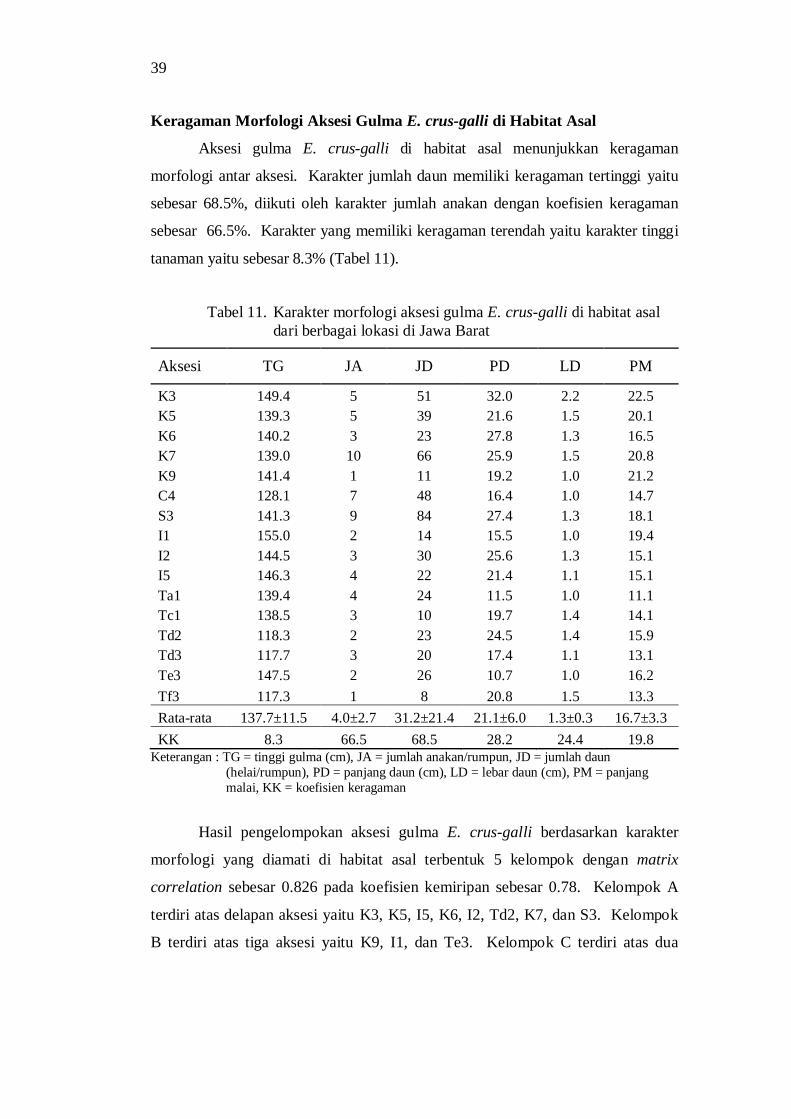
39
Keragaman Morfologi Aksesi Gulma E. crus-galli di Habitat Asal
Aksesi gulma E. crus-galli di habitat asal menunjukkan keragaman
morfologi antar aksesi. Karakter jumlah daun memiliki keragaman tertinggi yaitu
sebesar 68.5%, diikuti oleh karakter jumlah anakan dengan koefisien keragaman
sebesar 66.5%. Karakter yang memiliki keragaman terendah yaitu karakter tinggi
tanaman yaitu sebesar 8.3% (Tabel 11).
Tabel 11. Karakter morfologi aksesi gulma E. crus-galli di habitat asal
dari berbagai lokasi di Jawa Barat
Aksesi TG JA JD PD LD PM
K3 149.4 5 51 32.0 2.2 22.5 K5 139.3 5 39 21.6 1.5 20.1 K6 140.2 3 23 27.8 1.3 16.5 K7 139.0 10 66 25.9 1.5 20.8 K9 141.4 1 11 19.2 1.0 21.2 C4 128.1 7 48 16.4 1.0 14.7 S3 141.3 9 84 27.4 1.3 18.1 I1 155.0 2 14 15.5 1.0 19.4 I2 144.5 3 30 25.6 1.3 15.1 I5 146.3 4 22 21.4 1.1 15.1 Ta1 139.4 4 24 11.5 1.0 11.1 Tc1 138.5 3 10 19.7 1.4 14.1 Td2 118.3 2 23 24.5 1.4 15.9 Td3 117.7 3 20 17.4 1.1 13.1 Te3 147.5 2 26 10.7 1.0 16.2 Tf3 117.3 1 8 20.8 1.5 13.3 Rata-rata 137.7±11.5 4.0±2.7 31.2±21.4 21.1±6.0 1.3±0.3 16.7±3.3 KK 8.3 66.5 68.5 28.2 24.4 19.8
Keterangan : TG = tinggi gulma (cm), JA = jumlah anakan/rumpun, JD = jumlah daun (helai/rumpun), PD = panjang daun (cm), LD = lebar daun (cm), PM = panjang malai, KK = koefisien keragaman
Hasil pengelompokan aksesi gulma E. crus-galli berdasarkan karakter
morfologi yang diamati di habitat asal terbentuk 5 kelompok dengan matrix
correlation sebesar 0.826 pada koefisien kemiripan sebesar 0.78. Kelompok A
terdiri atas delapan aksesi yaitu K3, K5, I5, K6, I2, Td2, K7, dan S3. Kelompok
B terdiri atas tiga aksesi yaitu K9, I1, dan Te3. Kelompok C terdiri atas dua
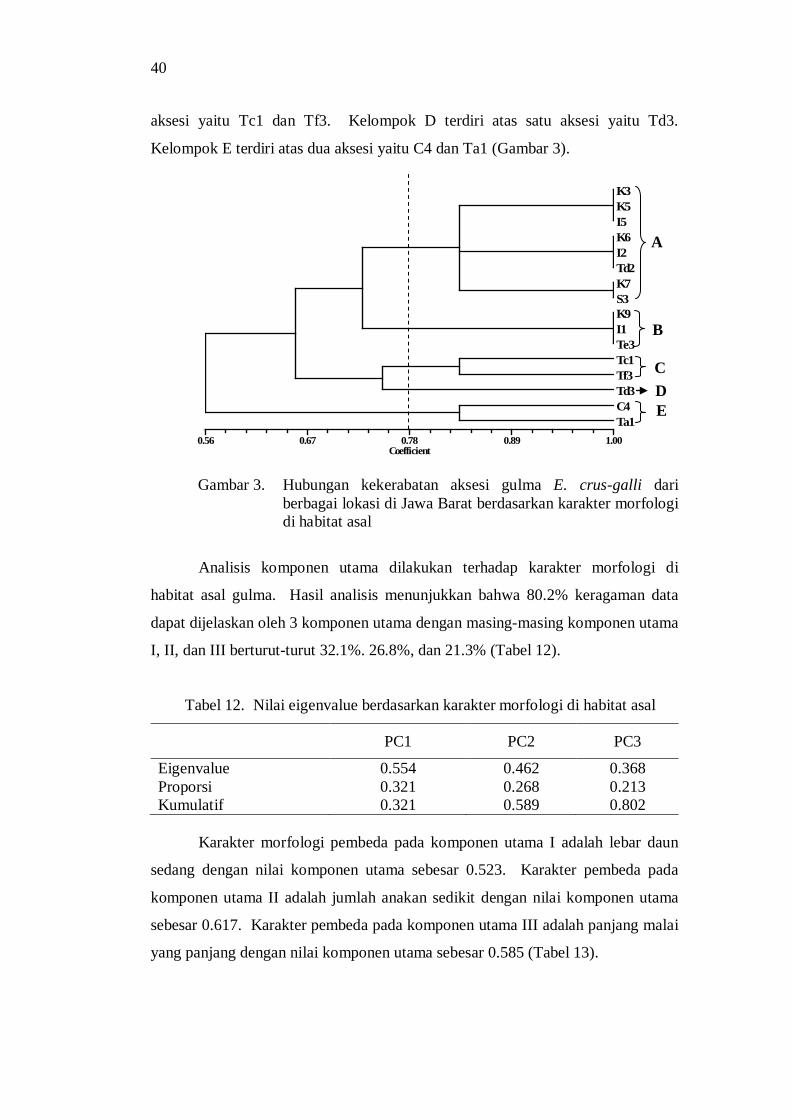
40
aksesi yaitu Tc1 dan Tf3. Kelompok D terdiri atas satu aksesi yaitu Td3.
Kelompok E terdiri atas dua aksesi yaitu C4 dan Ta1 (Gambar 3).
Gambar 3. Hubungan kekerabatan aksesi gulma E. crus-galli dari berbagai lokasi di Jawa Barat berdasarkan karakter morfologi di habitat asal
Analisis komponen utama dilakukan terhadap karakter morfologi di
habitat asal gulma. Hasil analisis menunjukkan bahwa 80.2% keragaman data
dapat dijelaskan oleh 3 komponen utama dengan masing-masing komponen utama
I, II, dan III berturut-turut 32.1%. 26.8%, dan 21.3% (Tabel 12).
Tabel 12. Nilai eigenvalue berdasarkan karakter morfologi di habitat asal
PC1 PC2 PC3
Eigenvalue 0.554 0.462 0.368 Proporsi 0.321 0.268 0.213 Kumulatif 0.321 0.589 0.802
Karakter morfologi pembeda pada komponen utama I adalah lebar daun
sedang dengan nilai komponen utama sebesar 0.523. Karakter pembeda pada
komponen utama II adalah jumlah anakan sedikit dengan nilai komponen utama
sebesar 0.617. Karakter pembeda pada komponen utama III adalah panjang malai
yang panjang dengan nilai komponen utama sebesar 0.585 (Tabel 13).
Coefficient0.56 0.67 0.78 0.89 1.00
K3 K5 I5 K6 I2 Td2 K7 S3 K9 I1 Te3 Tc1 Tf3 Td3 C4 Ta1
A
B
C D E
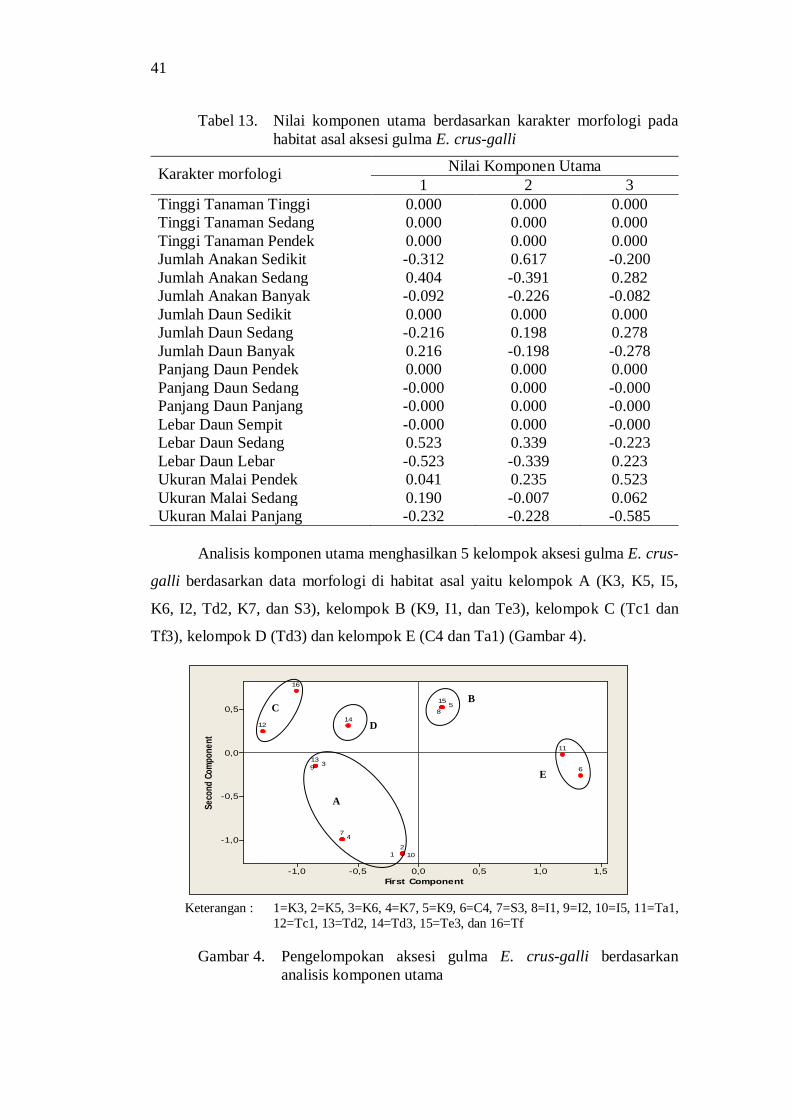
41
Tabel 13. Nilai komponen utama berdasarkan karakter morfologi pada habitat asal aksesi gulma E. crus-galli
Karakter morfologi Nilai Komponen Utama 1 2 3
Tinggi Tanaman Tinggi 0.000 0.000 0.000 Tinggi Tanaman Sedang 0.000 0.000 0.000 Tinggi Tanaman Pendek 0.000 0.000 0.000 Jumlah Anakan Sedikit -0.312 0.617 -0.200 Jumlah Anakan Sedang 0.404 -0.391 0.282 Jumlah Anakan Banyak -0.092 -0.226 -0.082 Jumlah Daun Sedikit 0.000 0.000 0.000 Jumlah Daun Sedang -0.216 0.198 0.278 Jumlah Daun Banyak 0.216 -0.198 -0.278 Panjang Daun Pendek 0.000 0.000 0.000 Panjang Daun Sedang -0.000 0.000 -0.000 Panjang Daun Panjang -0.000 0.000 -0.000 Lebar Daun Sempit -0.000 0.000 -0.000 Lebar Daun Sedang 0.523 0.339 -0.223 Lebar Daun Lebar -0.523 -0.339 0.223 Ukuran Malai Pendek 0.041 0.235 0.523 Ukuran Malai Sedang 0.190 -0.007 0.062 Ukuran Malai Panjang -0.232 -0.228 -0.585
Analisis komponen utama menghasilkan 5 kelompok aksesi gulma E. crus-
galli berdasarkan data morfologi di habitat asal yaitu kelompok A (K3, K5, I5,
K6, I2, Td2, K7, dan S3), kelompok B (K9, I1, dan Te3), kelompok C (Tc1 dan
Tf3), kelompok D (Td3) dan kelompok E (C4 dan Ta1) (Gambar 4).
Keterangan : 1=K3, 2=K5, 3=K6, 4=K7, 5=K9, 6=C4, 7=S3, 8=I1, 9=I2, 10=I5, 11=Ta1,
12=Tc1, 13=Td2, 14=Td3, 15=Te3, dan 16=Tf
Gambar 4. Pengelompokan aksesi gulma E. crus-galli berdasarkan analisis komponen utama
1,51,00,50,0-0,5-1,0
0,5
0,0
-0,5
-1,0
First Component
Seco
nd C
ompo
nent
16
15
14
13
12
11
10
9
8
7
6
5
4
3
21
C
D
B
A
E
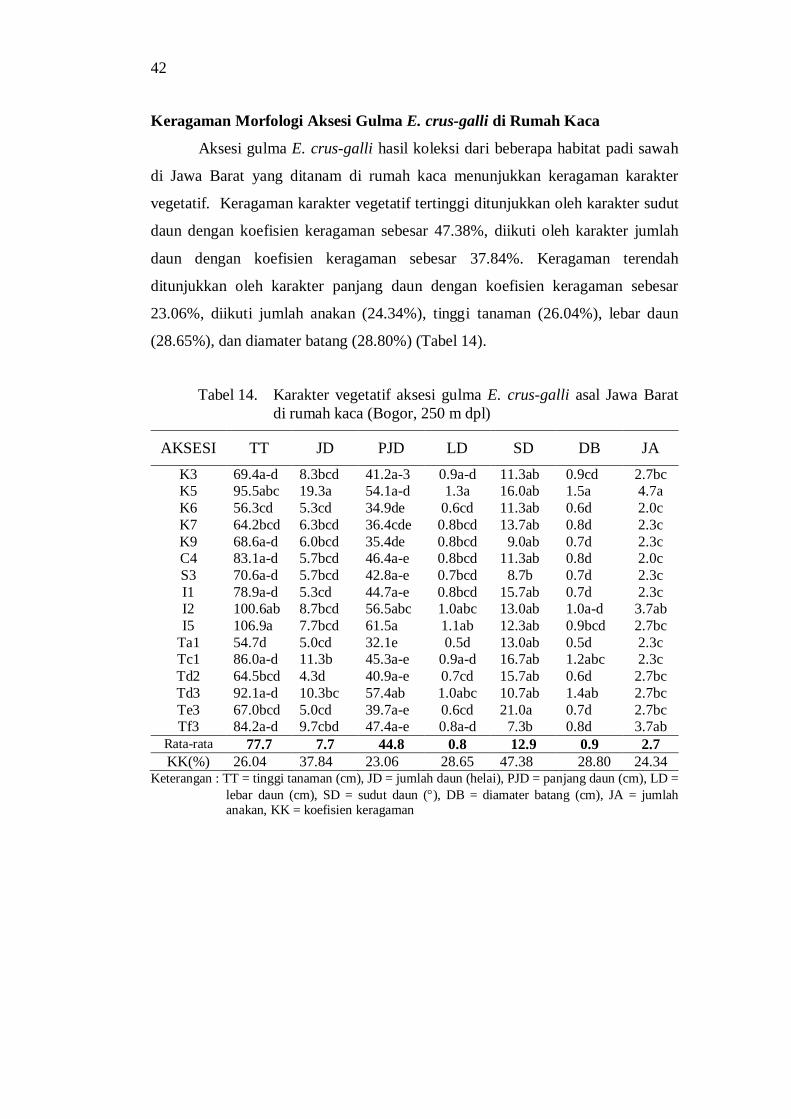
42
Keragaman Morfologi Aksesi Gulma E. crus-galli di Rumah Kaca
Aksesi gulma E. crus-galli hasil koleksi dari beberapa habitat padi sawah
di Jawa Barat yang ditanam di rumah kaca menunjukkan keragaman karakter
vegetatif. Keragaman karakter vegetatif tertinggi ditunjukkan oleh karakter sudut
daun dengan koefisien keragaman sebesar 47.38%, diikuti oleh karakter jumlah
daun dengan koefisien keragaman sebesar 37.84%. Keragaman terendah
ditunjukkan oleh karakter panjang daun dengan koefisien keragaman sebesar
23.06%, diikuti jumlah anakan (24.34%), tinggi tanaman (26.04%), lebar daun
(28.65%), dan diamater batang (28.80%) (Tabel 14).
Tabel 14. Karakter vegetatif aksesi gulma E. crus-galli asal Jawa Barat
di rumah kaca (Bogor, 250 m dpl)
AKSESI TT JD PJD LD SD DB JA K3 69.4a-d 8.3bcd 41.2a-3 0.9a-d 11.3ab 0.9cd 2.7bc K5 95.5abc 19.3a 54.1a-d 1.3a 16.0ab 1.5a 4.7a K6 56.3cd 5.3cd 34.9de 0.6cd 11.3ab 0.6d 2.0c K7 64.2bcd 6.3bcd 36.4cde 0.8bcd 13.7ab 0.8d 2.3c K9 68.6a-d 6.0bcd 35.4de 0.8bcd 9.0ab 0.7d 2.3c C4 83.1a-d 5.7bcd 46.4a-e 0.8bcd 11.3ab 0.8d 2.0c S3 70.6a-d 5.7bcd 42.8a-e 0.7bcd 8.7b 0.7d 2.3c I1 78.9a-d 5.3cd 44.7a-e 0.8bcd 15.7ab 0.7d 2.3c I2 100.6ab 8.7bcd 56.5abc 1.0abc 13.0ab 1.0a-d 3.7ab I5 106.9a 7.7bcd 61.5a 1.1ab 12.3ab 0.9bcd 2.7bc
Ta1 54.7d 5.0cd 32.1e 0.5d 13.0ab 0.5d 2.3c Tc1 86.0a-d 11.3b 45.3a-e 0.9a-d 16.7ab 1.2abc 2.3c Td2 64.5bcd 4.3d 40.9a-e 0.7cd 15.7ab 0.6d 2.7bc Td3 92.1a-d 10.3bc 57.4ab 1.0abc 10.7ab 1.4ab 2.7bc Te3 67.0bcd 5.0cd 39.7a-e 0.6cd 21.0a 0.7d 2.7bc Tf3 84.2a-d 9.7cbd 47.4a-e 0.8a-d 7.3b 0.8d 3.7ab
Rata-rata 77.7 7.7 44.8 0.8 12.9 0.9 2.7 KK(%) 26.04 37.84 23.06 28.65 47.38 28.80 24.34
Keterangan : TT = tinggi tanaman (cm), JD = jumlah daun (helai), PJD = panjang daun (cm), LD = lebar daun (cm), SD = sudut daun (°), DB = diamater batang (cm), JA = jumlah anakan, KK = koefisien keragaman
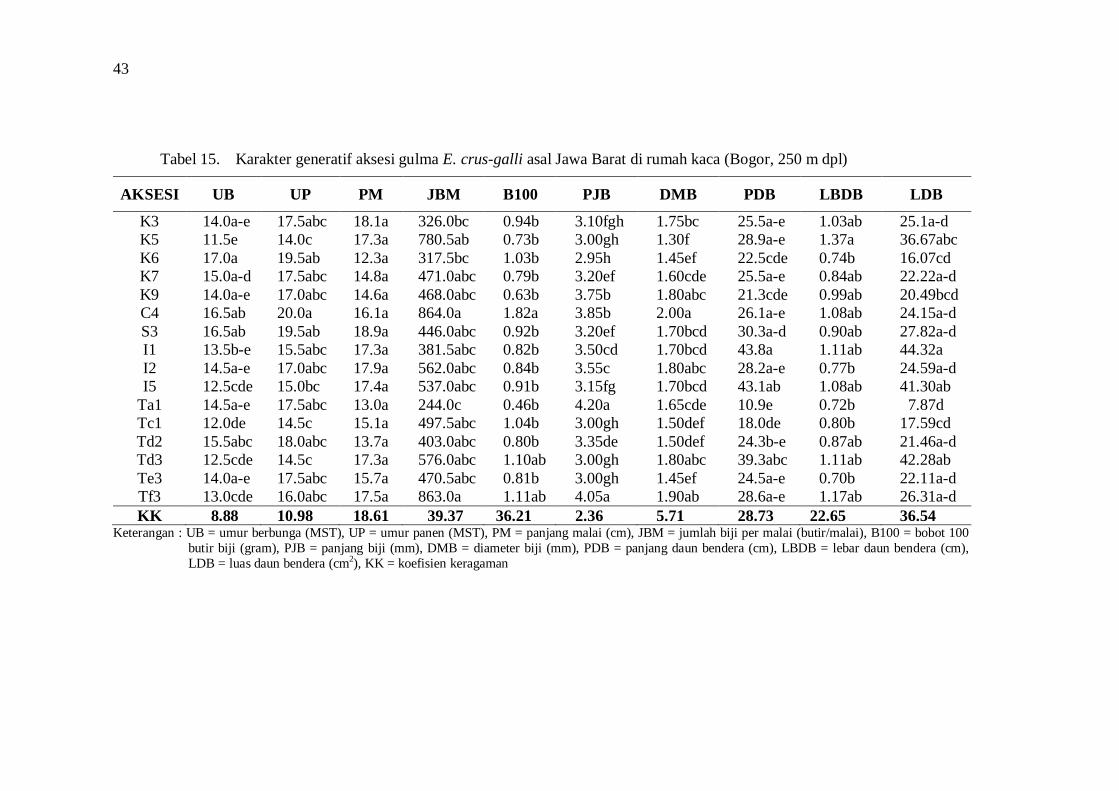
43
Tabel 15. Karakter generatif aksesi gulma E. crus-galli asal Jawa Barat di rumah kaca (Bogor, 250 m dpl)
AKSESI UB UP PM JBM B100 PJB DMB PDB LBDB LDB
K3 14.0a-e 17.5abc 18.1a 326.0bc 0.94b 3.10fgh 1.75bc 25.5a-e 1.03ab 25.1a-d K5 11.5e 14.0c 17.3a 780.5ab 0.73b 3.00gh 1.30f 28.9a-e 1.37a 36.67abc K6 17.0a 19.5ab 12.3a 317.5bc 1.03b 2.95h 1.45ef 22.5cde 0.74b 16.07cd K7 15.0a-d 17.5abc 14.8a 471.0abc 0.79b 3.20ef 1.60cde 25.5a-e 0.84ab 22.22a-d K9 14.0a-e 17.0abc 14.6a 468.0abc 0.63b 3.75b 1.80abc 21.3cde 0.99ab 20.49bcd C4 16.5ab 20.0a 16.1a 864.0a 1.82a 3.85b 2.00a 26.1a-e 1.08ab 24.15a-d S3 16.5ab 19.5ab 18.9a 446.0abc 0.92b 3.20ef 1.70bcd 30.3a-d 0.90ab 27.82a-d I1 13.5b-e 15.5abc 17.3a 381.5abc 0.82b 3.50cd 1.70bcd 43.8a 1.11ab 44.32a I2 14.5a-e 17.0abc 17.9a 562.0abc 0.84b 3.55c 1.80abc 28.2a-e 0.77b 24.59a-d I5 12.5cde 15.0bc 17.4a 537.0abc 0.91b 3.15fg 1.70bcd 43.1ab 1.08ab 41.30ab
Ta1 14.5a-e 17.5abc 13.0a 244.0c 0.46b 4.20a 1.65cde 10.9e 0.72b 7.87d Tc1 12.0de 14.5c 15.1a 497.5abc 1.04b 3.00gh 1.50def 18.0de 0.80b 17.59cd Td2 15.5abc 18.0abc 13.7a 403.0abc 0.80b 3.35de 1.50def 24.3b-e 0.87ab 21.46a-d Td3 12.5cde 14.5c 17.3a 576.0abc 1.10ab 3.00gh 1.80abc 39.3abc 1.11ab 42.28ab Te3 14.0a-e 17.5abc 15.7a 470.5abc 0.81b 3.00gh 1.45ef 24.5a-e 0.70b 22.11a-d Tf3 13.0cde 16.0abc 17.5a 863.0a 1.11ab 4.05a 1.90ab 28.6a-e 1.17ab 26.31a-d KK 8.88 10.98 18.61 39.37 36.21 2.36 5.71 28.73 22.65 36.54
Keterangan : UB = umur berbunga (MST), UP = umur panen (MST), PM = panjang malai (cm), JBM = jumlah biji per malai (butir/malai), B100 = bobot 100 butir biji (gram), PJB = panjang biji (mm), DMB = diameter biji (mm), PDB = panjang daun bendera (cm), LBDB = lebar daun bendera (cm), LDB = luas daun bendera (cm2
), KK = koefisien keragaman

44
Karakter generatif dari tiap aksesi gulma E. crus-galli dari beberapa
habitat sawah di Jawa Barat menunjukkan keragaman. Karakter generatif dengan
keragaman tertinggi ditunjukkan oleh karakter jumlah biji per malai dengan
koefisien keragaman sebesar 39.37%, diikuti karakter luas daun bendera
(36.54%), bobot biji 100 butir (36.21%), panjang daun bendera (28.73%) dan
lebar daun bendera (22.65%). Keragaman terendah ditunjukkan oleh karakter
panjang biji dengan koefisien keragaman sebesar 2.36%, disusul oleh karakter
diameter biji dengan koefisien keragaman sebesar 5.71% (Tabel 15).
Hasil analisis pengelompokan berdasarkan karakter morfologi yang
meliputi gabungan antara karakter vegetatif dan generatif diperoleh 5 grup pada
koefisien kemiripan sebesar 0.5, yaitu grup A, grup B, grup C, grup D, dan grup
E. Grup A terdiri atas 7 aksesi yaitu aksesi K7, K9, K3, Te3, Td2, S3, dan I2.
Grup B terdiri atas dua aksesi yaitu K6 dan Ta1. Grup C terdiri atas tiga aksesi
yaitu K5, Td3 dan I5. Grup D terdiri atas tiga aksesi yaitu aksesi Tf3, C4, dan I1.
Grup E terdiri atas satu aksesi yaitu Tc1 (Gambar 5).
Gambar 5. Dendogram kekerabatan aksesi gulma E. crus-galli
berdasarkan karakter morfologi di rumah kaca
Analisis komponen utama dilakukan untuk mengetahui ciri atau karakter
yang membedakan setiap aksesi, dimana dengan analisis gerombol (cluster
analysis) hanya mengetahui pengelompokan berdasarkan karakter tertentu, tetapi
Coefficient0.30 0.45 0.59 0.73 0.88
K7 K9 Te3 Td2 K3 S3 I2 K6 Ta1 K5 Td3 I5 Tf3 C4 I1 Tc1
D
E
C
B
A
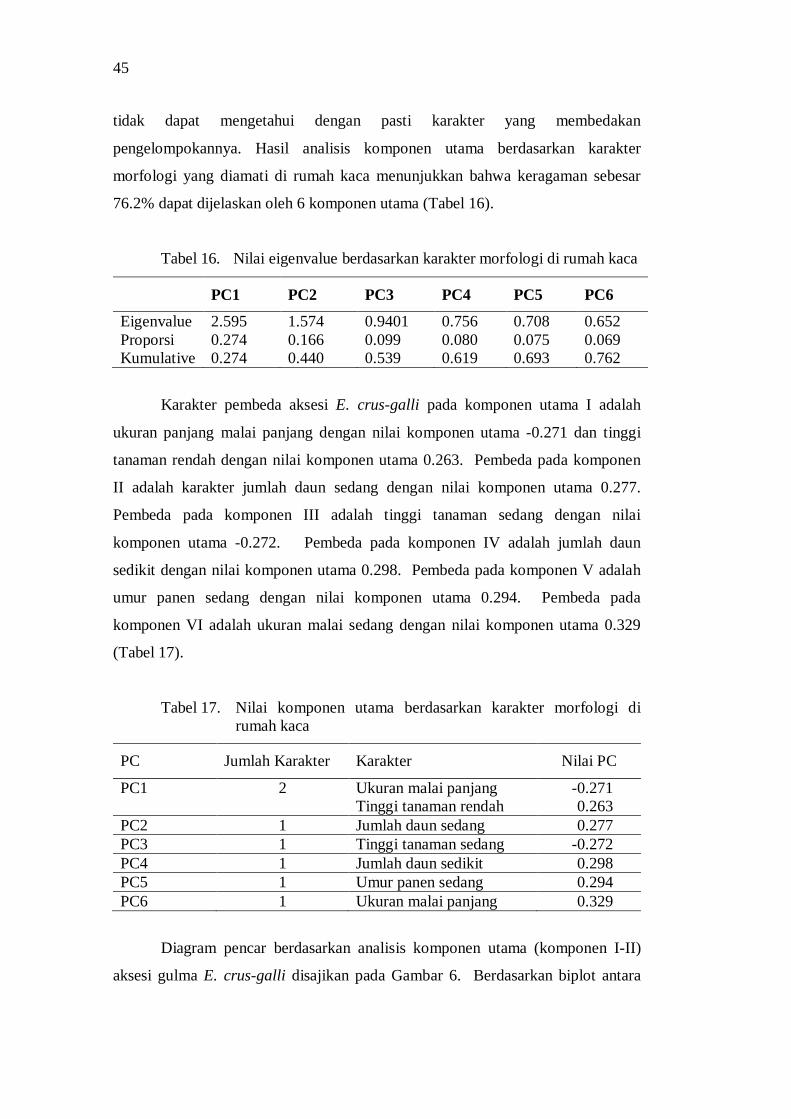
45
tidak dapat mengetahui dengan pasti karakter yang membedakan
pengelompokannya. Hasil analisis komponen utama berdasarkan karakter
morfologi yang diamati di rumah kaca menunjukkan bahwa keragaman sebesar
76.2% dapat dijelaskan oleh 6 komponen utama (Tabel 16).
Tabel 16. Nilai eigenvalue berdasarkan karakter morfologi di rumah kaca
PC1 PC2 PC3 PC4 PC5 PC6
Eigenvalue 2.595 1.574 0.9401 0.756 0.708 0.652 Proporsi 0.274 0.166 0.099 0.080 0.075 0.069 Kumulative 0.274 0.440 0.539 0.619 0.693 0.762
Karakter pembeda aksesi E. crus-galli pada komponen utama I adalah
ukuran panjang malai panjang dengan nilai komponen utama -0.271 dan tinggi
tanaman rendah dengan nilai komponen utama 0.263. Pembeda pada komponen
II adalah karakter jumlah daun sedang dengan nilai komponen utama 0.277.
Pembeda pada komponen III adalah tinggi tanaman sedang dengan nilai
komponen utama -0.272. Pembeda pada komponen IV adalah jumlah daun
sedikit dengan nilai komponen utama 0.298. Pembeda pada komponen V adalah
umur panen sedang dengan nilai komponen utama 0.294. Pembeda pada
komponen VI adalah ukuran malai sedang dengan nilai komponen utama 0.329
(Tabel 17).
Tabel 17. Nilai komponen utama berdasarkan karakter morfologi di rumah kaca
PC Jumlah Karakter Karakter Nilai PC
PC1 2 Ukuran malai panjang -0.271 Tinggi tanaman rendah 0.263 PC2 1 Jumlah daun sedang 0.277 PC3 1 Tinggi tanaman sedang -0.272 PC4 1 Jumlah daun sedikit 0.298 PC5 1 Umur panen sedang 0.294 PC6 1 Ukuran malai panjang 0.329
Diagram pencar berdasarkan analisis komponen utama (komponen I-II)
aksesi gulma E. crus-galli disajikan pada Gambar 6. Berdasarkan biplot antara

46
komponen utama I-II aksesi gulma E. crus-galli asal Jawa Barat memencar dalam
empat kelompok, yaitu kelompok A, B, C, dan D.
Keterangan : 1=K3, 2=K5, 3=K6, 4=K7, 5=K9, 6=C4, 7=S3, 8=I1, 9=I2, 10=I5, 11=Ta1,
12=Tc1, 13=Td2, 14=Td3, 15=Te3, dan 16=Tf
Gambar 6. Diagram pencar (komponen I-II) aksesi gulma E. crus-galli berdasarkan karakter vegetatif di rumah kaca
Keragaman Genetik Aksesi Gulma E. crus-galli
Aksesi gulma E. crus-galli yang berasal dari berbagai habitat padi sawah
di Jawa Barat menunjukkan adanya keragaman genetik berdasarkan marka
molekuler Simple Sequence Repeat (SSR). Keragaman genetik antar aksesi gulma
E. crus-galli menunjukkan keragaman yang rendah. Hal ini diduga karena gulma
E. crus-galli merupakan gulma yang menyerbuk sendiri (Honk et al. 1996).
Secara umum, spesies yang menyerbuk sendiri memiliki variasi genetik yang
rendah, sebaliknya untuk spesies yang menyerbuk silang akan memiliki variasi
genetik yang tinggi (Lin et al. 2005).
Dendogram menunjukkan bahwa aksesi mengelompok menjadi 4 sub
grup, yaitu A, B, C dan D. Sub Grup A memiliki anggota paling besar yaitu
11 aksesi diikuti dengan sub grup C (3 aksesi). Sub Grup B dan D memiliki satu
anggota, yaitu secara berturut-turut aksesi dari Karawang (K3) dan aksesi dari
Pangalengan (Tf3).
3210-1-2-3
3
2
1
0
-1
-2
First Component
Seco
nd C
ompo
nent
16
15
14
13
12
11
10
9
8
7 6
5
4
3
2
1
C B
A
D
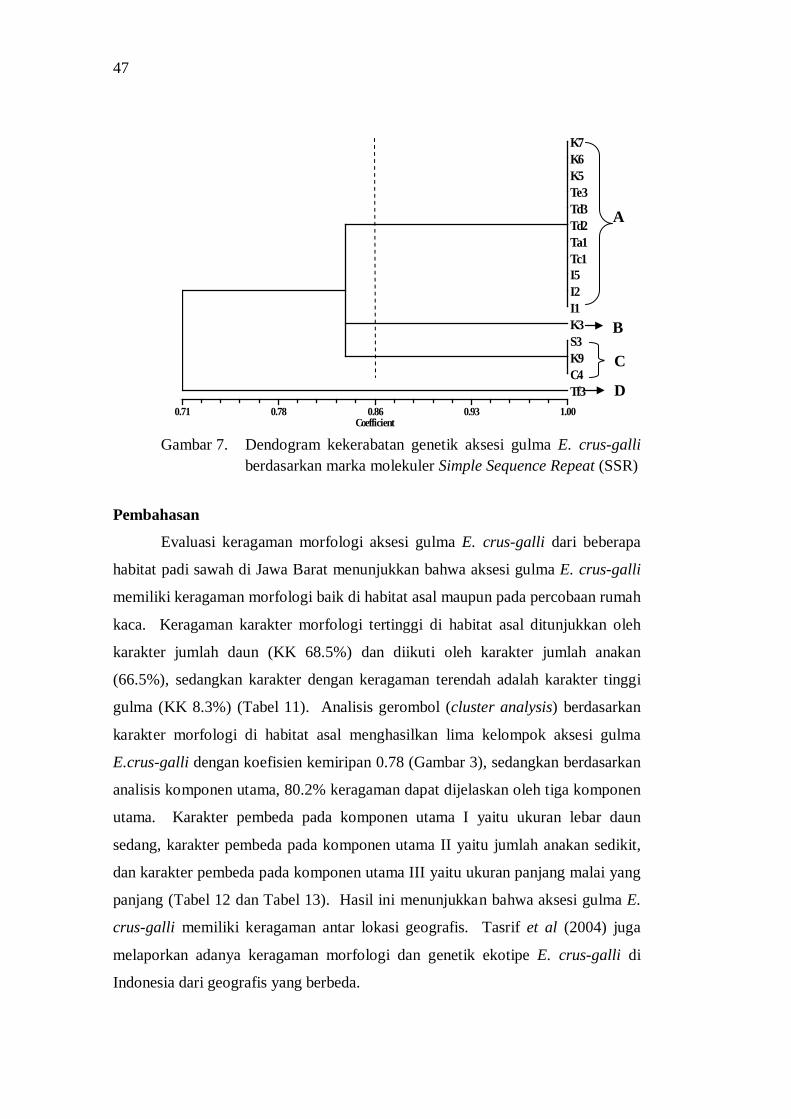
47
Gambar 7. Dendogram kekerabatan genetik aksesi gulma E. crus-galli
berdasarkan marka molekuler Simple Sequence Repeat (SSR)
Pembahasan
Evaluasi keragaman morfologi aksesi gulma E. crus-galli dari beberapa
habitat padi sawah di Jawa Barat menunjukkan bahwa aksesi gulma E. crus-galli
memiliki keragaman morfologi baik di habitat asal maupun pada percobaan rumah
kaca. Keragaman karakter morfologi tertinggi di habitat asal ditunjukkan oleh
karakter jumlah daun (KK 68.5%) dan diikuti oleh karakter jumlah anakan
(66.5%), sedangkan karakter dengan keragaman terendah adalah karakter tinggi
gulma (KK 8.3%) (Tabel 11). Analisis gerombol (cluster analysis) berdasarkan
karakter morfologi di habitat asal menghasilkan lima kelompok aksesi gulma
E.crus-galli dengan koefisien kemiripan 0.78 (Gambar 3), sedangkan berdasarkan
analisis komponen utama, 80.2% keragaman dapat dijelaskan oleh tiga komponen
utama. Karakter pembeda pada komponen utama I yaitu ukuran lebar daun
sedang, karakter pembeda pada komponen utama II yaitu jumlah anakan sedikit,
dan karakter pembeda pada komponen utama III yaitu ukuran panjang malai yang
panjang (Tabel 12 dan Tabel 13). Hasil ini menunjukkan bahwa aksesi gulma E.
crus-galli memiliki keragaman antar lokasi geografis. Tasrif et al (2004) juga
melaporkan adanya keragaman morfologi dan genetik ekotipe E. crus-galli di
Indonesia dari geografis yang berbeda.
Coefficient0.71 0.78 0.86 0.93 1.00
K7 K6 K5 Te3 Td3 Td2 Ta1 Tc1 I5 I2 I1 K3 S3 K9 C4 Tf3
A
C
B
D

48
Hasil evaluasi keragaman morfologi di rumah kaca juga menunjukkan
adanya keragaman morfologi antar aksesi. Keragaman tertinggi ditunjukkan oleh
karakter sudut daun (KK 47.38%), diikuti oleh karakter jumlah biji per malai
(39.37%) dan jumlah daun (KK 37.84%), sedangkan keragaman terendah
ditunjukkan oleh karakter panjang biji (KK 2.36%) dan diameter biji (KK 5.71%)
(Tabel 14 dan Tabel 15). Berdasarkan analisis gerombol, terbentuk lima
kelompok aksesi gulma E. crus-galli pada koefisien kemiripan 0.5 (Gambar 5)
dan berdasarkan analisis komponen utama 76.2% keragaman dapat dijelaskan
oleh 6 komponen utama (Tabel 16). Karakter pembeda pada komponen utama I
adalah ukuran panjang malai yang panjang dan tinggi tanaman rendah. Karakter
pembeda pada komponen utama II – VI berturut-turut yaitu jumlah daun sedang,
tinggi tanaman sedang, jumlah daun sedikit, umur panen sedang, dan ukuran
malai panjang (Tabel 17).
Pengelompokan yang berbeda pada beberapa aksesi gulma E. crus-galli
antara kondisi habitat asal dengan habitat rumah kaca menunjukkan bahwa gulma
E. crus-galli memiliki kemampuan plastisitas fenotipik. Plastisitas fenotipik ini
ditunjukkan oleh karakter morfologi yang memiliki keragaman tinggi di habitat
asal seperti karakter jumlah daun rata-rata 31.2 helai (KK 68.5%), pada kondisi
rumah kaca rata-rata 7.7 helai daun. Karakter panjang daun pada habitat asal rata-
rata sebesar 21.1 cm, pada kondisi rumah kaca menjadi 44.8 cm. Menurut Altop
et al. (2011) keragaman fenotip dipengaruhi praktek budidaya tanaman,
karakteristik tanaman, geografis, dan adanya tekanan penggunaan herbisida.
Dendogram berdasarkan karakter genetik menunjukkan bahwa sub grup A
memiliki aksesi yang berasal dari seluruh aksesi Indramayu (I1, I2, dan I5),
Cianjur (Ta1, Td2, Td3), Sukabumi (Tc1), Pangalengan (Te3), dan sebagian besar
aksesi dari Karawang (K5, K6, K7). Aksesi asal Subang (S3), aksesi asal
Cikampek (C4), serta 1 aksesi asal Karawang (K9) tergabung dalam sub grup C
(Gambar 7). Pengelompokan A dan C menunjukkan adanya pengelompokan
berdasarkan zona geografi. Secara geografis, Cianjur, Sukabumi dan Karawang
merupakan wilayah Jawa Barat bagian barat yang berbatasan langsung, dan
berdasarkan dendogram mengelompok menjadi satu cluster. Demikian juga
Subang dan Cikampek serta Karawang juga merupakan satu cluster C yang dapat

49
dimasukkkan dalam zona geografis Jawa Barat bagian utara. Ketiga wilayah
tersebut secara administratif berbatasan langsung. Perkecualian dari sub grup B
dan D yang memisah dari sub grup lain secara geografis berjauhan. Selain itu,
secara altitude, kedua tempat tersebut mewakili dataran rendah dan dataran tinggi
(Tabel 8). Dengan demikian, perkecualian tersebut masih mengindikasikan
pewilayahan gulma E. crus-galli secara geografis yang kuat.
Adanya keragaman genetik pada aksesi dari Karawang dan dari
Pangalengan yang terbagi menjadi dua sub grup sangat menarik untuk dikaji. Dari
sisi geografik, Karawang memiliki dataran tinggi yang berbatasan langsung
dengan dataran tinggi Cianjur dan Cikampek (Kabupaten Purwakarta). Asal aksesi
K3 dan K9 dari Kabupaten Karawang terhubungkan langsung dengan jaringan
irigasi sawah asal Waduk Jatiluhur di Purwakarta. Dengan demikian, ada dua
kemungkinan asal off type dari aksesi Karawang yang masuk dalam sub grup B
dan C, memisah menjadi sub grup tersendiri dari sebagian besar anggota di sub
grup A. Pertama, adalah karena perpindahan material genetik melalui jaringan
irigasi dan kedua, adalah karena perpindahan hasil panen padi atau terbawa oleh
pekerja pertanian. Kedua kejadian tersebut sangat mungkin, namun dalam
penelitian ini tidak dapat ditentukan secara tegas, mana yang paling mungkin
terjadi. Pada kasus Pangalengan, aksesi Tf3 yang merupakan sub grup D, lebih
dimungkinkan karena adanya isolasi jarak. Gulma E. cruss-galli kode Tf3 diambil
dari sawah di ketinggian di atas 1400 m di atas permukaan laut. Ada
kemungkinan, pada ketinggian tersebut mode penyerbukan yang secara umum
bagi E. cruss-galli adalah menyerbuk sendiri, diduga dapat berubah dengan
adanya porsi menyerbuk silang. Namun demikian, perlu adanya kajian lebih lanjut
apakah hipotesis tersebut benar. Seperti diketahui secara umum bahwa keragaman
genetik penyerbuk di daerah pertanian intensif akan lebih rendah dibandingkan
dengan daerah pertanian yang kurang intensif. Sejarah pengusahaan padi sawah
yang sangat panjang di daerah pantai utara Jawa, terutama penggunaan pestisida
yang sangat intensif diduga dapat mengurangi keragaman penyerbuk, termasuk
serangga penyerbuk E. crus-galli.
Dengan demikian, keragaman genetik E. crus-galli yang diamati pada
penelitian ini diduga disebabkan oleh beberapa faktor seperti perpindahan material

50
genetik secara tidak sengaja melalui perpindahan hasil panen padi atau bahan
tanaman padi, perpindahan melalui air irigasi, dan keragaman genetik akibat
adanya isolasi jarak, serta kemungkinan adanya mutasi. Menurut Altop et al.
(2011) adanya keragaman genetik E. crus-galli pada lokasi yang sama disebabkan
oleh perpindahan biji melalui alat pengolahan tanah, transportasi, dan perpindahan
hasil panen.
Berdasarkan dendogram karakter morfologi di habitat asal, karakter
morfologi di rumah kaca, dan karakter genetik terlihat bahwa aksesi K3, K7, S3,
I2, dan Td2 mengelompok dalam satu sub grup yang sama. Hal ini membuktikan
bahwa keragaman morfologi disebabkan oleh karakter genetik aksesi tersebut.
Sebaliknya, aksesi I5 dan I1 yang secara genetik sama dengan aksesi I2, ternyata
memperlihatkan fenotipik yang berbeda. Pada kondisi habitat asal I5
mengelompok bersama aksesi I2 ke dalam sub grup A, sedangkan I1 berada di
kelompok lain (sub grup B). Pada kondisi rumah kaca, I1, I2, dan I5 terpisah ke
dalam kelompok yang berbeda. Hasil ini menunjukkan bahwa keragaman
morfologi muncul sebagai akibat adanya sifat plastisitas fenotipik akibat kondisi
lingkungan yang berbeda.
Kesimpulan
Aksesi gulma E. crus-galli asal Jawa Barat menunjukkan keragaman
morfologi yang tinggi. Perbedaan antar asesi tidak dapat ditunjukkan oleh
karakter tunggal, tetapi oleh beberapa karakter morfologi yang utama yaitu lebar
daun, jumlah anakan, panjang malai, jumlah daun, tinggi tanaman, dan umur
panen.
Berdasarkan marka molekuler Simple Sequence Repeat (SSR), asesi gulma
E. crus-galli asal Jawa Barat menunjukkan adanya keragaman genetik.
Keragaman genetik ini diduga disebabkan oleh perpindahan material genetik
melalui hasil panen ataupun melalui irigasi, adanya isolasi jarak, dan
kemungkinan adanya mutasi.
Keragaman morfologi asesi gulma E. crus-galli disebabkan oleh
kemampuan plastisitas fenotipik dan faktor genetik.

IDENTIFIKASI POTENSI ALELOPATI BEBERAPA AKSESI GULMA Echinochloa crus-galli (L.) Beauv.
ASAL JAWA BARAT
ABSTRAK
Salah satu cara gulma E. crus-galli untuk mengganggu pertumbuhan dan produksi tanaman padi adalah melalui alelopati. Penelitian ini bertujuan untuk mengidentifikasi potensi alelopati aksesi gulma E. crus-galli asal Jawa Barat terhadap perkecambahan tanaman padi sawah. Percobaan dilakukan dengan rancangan acak kelompok dengan satu faktor yaitu larutan ekstrak akar segar 16 aksesi gulma E. crus-galli asal Jawa Barat. Sebagai kontrol, kertas merang diberi aquades sebanyak 2 ml. Satuan percobaan berupa cawan petri dengan 10 benih padi hibrida. Hasil percobaan menunjukkan bahwa aksesi gulma E. crus-galli asal Jawa Barat memiliki potensi alelopati yang ditunjukkan dengan adanya penghambatan ataupun pemanjangan plumula dan radikula kecambah padi. Penghambatan atau pemanjangan plumula dan radikula dipengaruhi oleh jumlah jenis dan konsentrasi senyawa yang terkandung dalam ekstrak akar tiap aksesi gulma. Senyawa alelopati yang teridentifikasi dari ekstrak akar aksesi E. crus-galli diantaranya adalah golongan senyawa phenolic, phtalic acid, decanoid acid, propanoid, quinon, dan sterol. Aksesi asal Karawang (K5, K6, K3) dan asal Cianjur (Ta1) lebih menekan plumula sedangkan aksesi asal Pangalengan (Td3) lebih menekan radikula. Kata kunci : Echinochloa crus-galli, alelopati, plumula, radikula.

52
IDENTIFICATION OF ALLELOPHATIC POTENTIAL OF SOME Echinochloa crus-galli (L.) Beauv. WEED
ACCESSION FROM WEST JAVA
ABSTRACT One way of E. crus-galli weed to interfere the growth and production of rice plants is through allelophaty. The research objective was to identify the allelophatic potential of E.crus-galli weed accession from West Java on the germination of rice. The experiments were conducted in randomized block design with one factor that was solution of fresh root extract of 16 E. crus-galli weed accessions from West Java. The experimental unit was petridish with 10 seeds of hybrid rice. The results showed that the accession of E. crus-galli weed from West Java has the allelophatic potential indicated by the inhibition or elongation of plumula and radicle. Inhibition or elongation of plumula and radicle was influenced by a number of types and concentrations of the compounds contained in extracts of the roots of each weed accessions. Allelophatic compounds identified from the root extract of E. crus-galli accession including phenolics, phtalic acid, decanoid acid, propanoid, quinone, and sterols. Accession of Karawang (K5, K6, K3) and Cianjur (Ta1) more suppressed plumula while the accession of Pangalengan (Td3) more suppressed radicle
.
Keywords : Echinochloa crus-galli, allelophaty, plumula, radicle.

53
Pendahuluan
Salah satu cara gulma untuk mengganggu pertumbuhan dan produksi
tanaman adalah melalui alelopati. Alelopati didefinisikan sebagai pengaruh
langsung ataupun tidak langsung dari suatu tumbuhan terhadap yang lainnya,
termasuk mikroorganisme, baik yang bersifat positif atau perangsangan maupun
negatif atau penghambatan terhadap pertumbuhan, melalui pelepasan senyawa
kimia ke lingkungannya (Rice 1984; 1995; Inderjit & Keating 1999; Singh et al.
2003). Senyawa alelopati dapat mempengaruhi aktivitas tumbuhan antara lain
dengan cara menghambat penyerapan hara oleh akar tanaman, pembelahan sel-sel
akar, pertumbuhan tanaman, aktivitas fotosintesis, mempengaruhi respirasi,
sintesis protein, menurunkan daya permeabilitas membran sel dan menghambat
aktivitas enzim (Sastroutomo 1990).
Gulma Echinochloa crus-galli memiliki potensi alelopati. Adanya potensi
alelopati pada gulma E. crus-galli telah dilaporkan oleh Yamamoto et al. (1999),
Xuan et al. (2006) dan Son (2010). Selama perkecambahan dan awal
pertumbuhan, E. crus-galli menekan pertumbuhan beberapa tanaman pertanian
termasuk padi dan E. crus-galli itu sendiri. Senyawa p-Hidroxymandelic acid
merupakan allelochemical dari eksudat akar E. crus-galli muda yang dapat
menekan kuat pertumbuhan tajuk dan pemanjangan akar padi (Yamamoto et al.
1999). Eksudat akar E. crus-galli dapat menekan perkecambahan dan
pertumbuhan tanaman padi, lettuce dan monochoria. Komponen yang terlibat
dalam aktivitas phytotoxic E. crus-galli antara lain phenolic, long-chain fatty
acids, lactones, diethyl phthalate, acenaphthene, phthalic acids, benzoic acid dan
decane. Penghambat pertumbuhan terbesar ditunjukkan oleh lactones, diikut i
oleh phenolic dan phthalic acid (Xuan et al. 2006).
Produksi senyawa alelopati sangat dipengaruhi oleh faktor lingkungan,
seperti kualitas, intensitas dan lamanya masa penyinaran cahaya, kondisi
kekurangan hara, dan gangguan kekeringan (Rice 1974). Penelitian tentang
interaksi padi dan gulma E. crus-galli saat ini terfokus pada potensi penekanan
gulma E. crus-galli oleh tanaman padi dan konsentrasi alelopati yang dihasilkan
oleh tanaman padi yang digunakan untuk mengendalikan gulma E. crus-galli
(Chung et al. 2003).

54
Penelitian tentang alelopati pada aksesi gulma E. crus-galli di Indonesia
masih sangat terbatas. Aksesi gulma E. crus-galli asal Jawa Barat diduga
memiliki potensi alelopati yang berbeda yang dapat menghambat pertumbuhan
tanaman padi dan menurunkan produksi padi. Penelitian ini bertujuan untuk
mengidentifikasi potensi alelopati aksesi gulma E. crus-galli asal Jawa Barat
terhadap perkecambahan tanaman padi sawah.
Bahan dan Metode
Penelitian dilaksanakan dari bulan Mei - Desember 2007 di Laboratorium
Umum, Departemen Agronomi dan Hortikultura, Fakultas Pertanian. Bahan
tanaman berupa akar gulma E. crus-galli hasil koleksi dari beberapa lokasi di
Jawa Barat, benih padi hibrida, dan aquadest. Alat yang digunakan antara lain
mortar, kertas merang, cawan petri, beaker, timbangan analitik, dan oven.
Percobaan dilakukan dengan Rancangan Acak Kelompok dengan satu
faktor yaitu 16 aksesi gulma E. crus-galli asal Jawa Barat. Potensi alelopati
diukur dari ekstrak akar segar gulma E. crus-galli menurut metode Yamamoto et.
al. (1999). Sebanyak 10 g akar gulma dihaluskan menggunakan mortar,
selanjutnya dilarutkan dalam 200 ml aquades dan disimpan selama 24 jam.
Setelah 24 jam, larutan ektrak akar disaring menggunakan kertas saring.
Selanjutnya, kertas merang (10 cm x 10 cm) diletakkan pada cawan petri,
kemudian diberi larutan ekstrak akar gulma sebanyak 2 ml (konsentrasi 5 % w/v)
untuk setiap perlakuan. Sebagai kontrol, kertas merang diberi aquades sebanyak 2
ml. Benih padi hibrida sebelum perlakuan direndam dalam larutan NaClO 1%
selama 7 menit, kemudian dibilas dengan aquades. Setelah itu, benih padi hibrida
disemai sebanyak 10 benih/perlakuan.
Peubah yang diamati yaitu tingkat penghambatan (Inhibition Rate=IR)
terhadap plumula dan radikula pada saat 14 hari setelah semai. IR plumula dan IR
radikula dihitung berdasarkan rumus :
Kandungan alelopati pada ekstrak akar aksesi gulma E. crus-galli dianalisis
dengan GCMS menurut metode Xuan et. al. ( 2006).
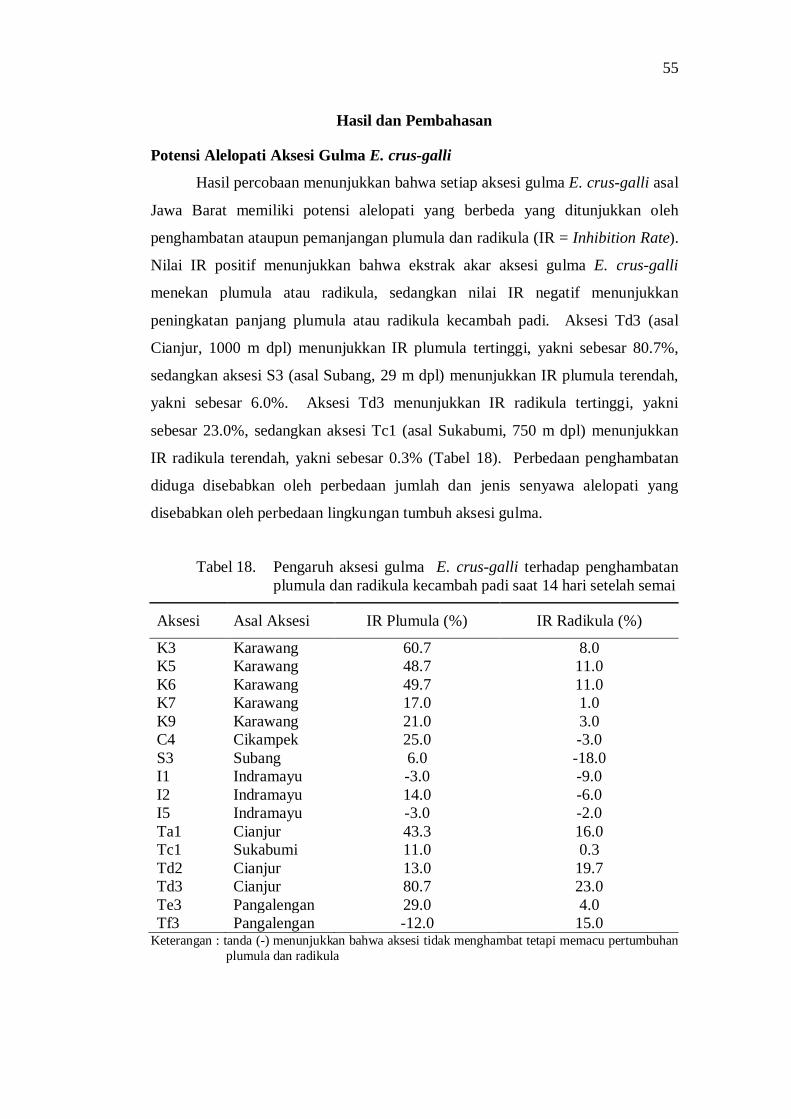
55
Hasil dan Pembahasan
Potensi Alelopati Aksesi Gulma E. crus-galli
Hasil percobaan menunjukkan bahwa setiap aksesi gulma E. crus-galli asal
Jawa Barat memiliki potensi alelopati yang berbeda yang ditunjukkan oleh
penghambatan ataupun pemanjangan plumula dan radikula (IR = Inhibition Rate).
Nilai IR positif menunjukkan bahwa ekstrak akar aksesi gulma E. crus-galli
menekan plumula atau radikula, sedangkan nilai IR negatif menunjukkan
peningkatan panjang plumula atau radikula kecambah padi. Aksesi Td3 (asal
Cianjur, 1000 m dpl) menunjukkan IR plumula tertinggi, yakni sebesar 80.7%,
sedangkan aksesi S3 (asal Subang, 29 m dpl) menunjukkan IR plumula terendah,
yakni sebesar 6.0%. Aksesi Td3 menunjukkan IR radikula tertinggi, yakni
sebesar 23.0%, sedangkan aksesi Tc1 (asal Sukabumi, 750 m dpl) menunjukkan
IR radikula terendah, yakni sebesar 0.3% (Tabel 18). Perbedaan penghambatan
diduga disebabkan oleh perbedaan jumlah dan jenis senyawa alelopati yang
disebabkan oleh perbedaan lingkungan tumbuh aksesi gulma.
Tabel 18. Pengaruh aksesi gulma E. crus-galli terhadap penghambatan
plumula dan radikula kecambah padi saat 14 hari setelah semai
Aksesi Asal Aksesi IR Plumula (%) IR Radikula (%)
K3 Karawang 60.7 8.0 K5 Karawang 48.7 11.0 K6 Karawang 49.7 11.0 K7 Karawang 17.0 1.0 K9 Karawang 21.0 3.0 C4 Cikampek 25.0 -3.0 S3 Subang 6.0 -18.0 I1 Indramayu -3.0 -9.0 I2 Indramayu 14.0 -6.0 I5 Indramayu -3.0 -2.0 Ta1 Cianjur 43.3 16.0 Tc1 Sukabumi 11.0 0.3 Td2 Cianjur 13.0 19.7 Td3 Cianjur 80.7 23.0 Te3 Pangalengan 29.0 4.0 Tf3 Pangalengan -12.0 15.0
Keterangan : tanda (-) menunjukkan bahwa aksesi tidak menghambat tetapi memacu pertumbuhan plumula dan radikula

56
Tidak semua aksesi menghambat pemanjangan plumula. Beberapa diantara
aksesi justru meningkatkan panjang plumula dan panjang radikula padi yang
ditunjukkan dengan nilai IR yang negatif, artinya panjang plumula dan panjang
radikula padi justru lebih panjang dibandingkan terhadap kontrol. Aksesi yang
meningkatkan panjang plumula antara lain I1 (IR -3.0), I5 (IR -3.0), dan Tf3
(IR -12.0). Aksesi yang meningkatkan panjang radikula antara lain aksesi C4
(IR -3.0), S3 (-18.0), I1 (IR -9.0), I2 (IR -6.0), dan I5 (IR -2.0) (Tabel 18).
Menurut Sastroutomo (1990), tumbuh-tumbuhan bervariasi dalam menghasilkan
senyawa alelopati (alelokimia) bergantung pada keadaan lingkungan tempat ia
tumbuh, dan gangguan serta tekanan lingkungan yang dialaminya. Menurut
Junaedi et al. (2006) perbedaan potensi alelopati dan pengaruhnya terhadap
tumbuhan lain dipengaruhi oleh genetik dan lingkungan. Perbedaan potensi
alelopati karena lingkungan dapat disebabkan oleh adanya perbedaan populasi,
siklus hidup, waktu tanam, tanah dan iklim, serta adanya cekaman biotik maupun
abiotik.
Hubungan Antara Potensi Alelopati dengan Keragaman Genetik Aksesi
Gulma E. crus-galli
Potensi alelopati aksesi gulma E. crus-galli dipengaruhi oleh faktor genetik
dan lingkungan tumbuh gulma. Analisis cluster berdasarkan potensi alelopatinya
menghasilkan enam kelompok aksesi pada koefisien kemiripan 0.72. Kelompok
A terdiri atas lima aksesi yaitu aksesi K7, S3, I5, I2, dan I1. Kelompok B terdiri
atas satu aksesi yaitu aksesi K3. Kelompok C terdiri atas tiga aksesi yaitu aksesi
Te3, K9, dan C4. Kelompok D terdiri atas tiga aksesi yaitu aksesi K6, K5, dan
Ta1. Kelompok E terdiri atas tiga aksesi yaitu aksesi Tf3, Td2, dan Tc1.
Kelompok F terdiri atas satu aksesi yaitu aksesi Td3. Aksesi asal Indramayu
terlihat mengelompok dalam satu kelompok yaitu kelompok A, sedangkan aksesi
asal Karawang terlihat menyebar ke dalam empat kelompok, yakni kelompok A
(K7), kelompok B (K3), kelompok C (K9), dan kelompok D (K6 dan K5).
Sementara aksesi Td3 asal Cianjur berada pada kelompok sendiri yaitu kelompok
F (Gambar 8).
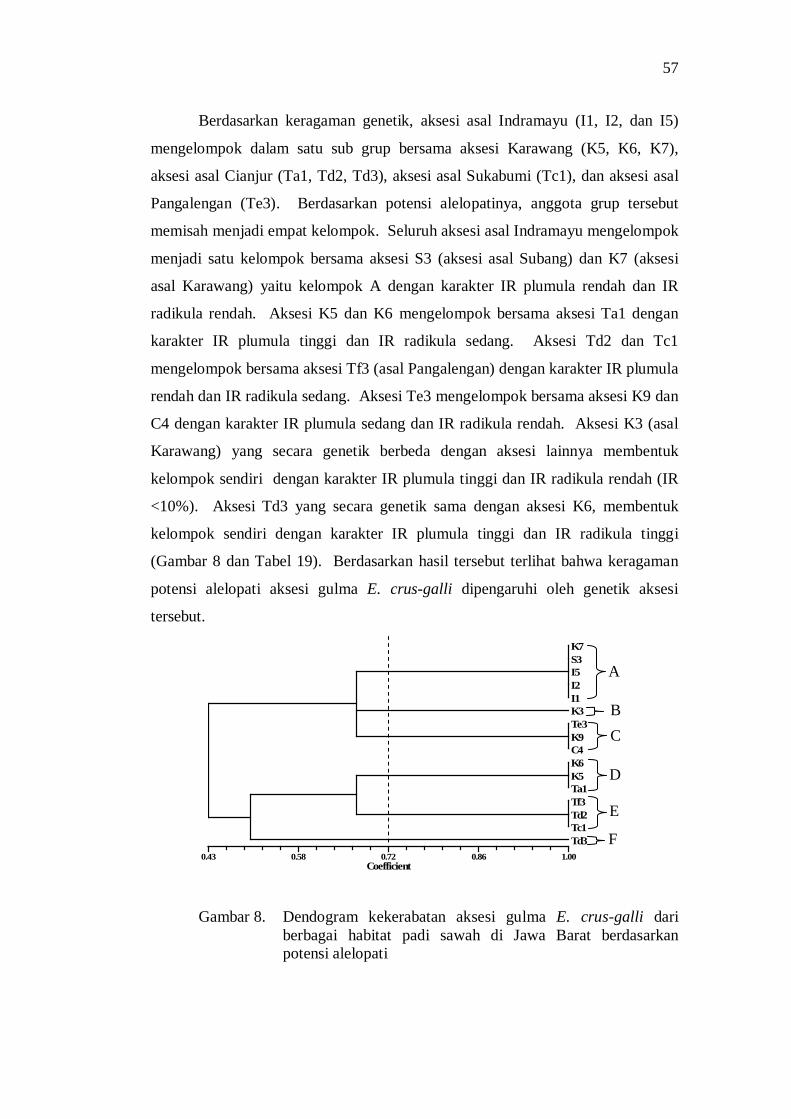
57
Berdasarkan keragaman genetik, aksesi asal Indramayu (I1, I2, dan I5)
mengelompok dalam satu sub grup bersama aksesi Karawang (K5, K6, K7),
aksesi asal Cianjur (Ta1, Td2, Td3), aksesi asal Sukabumi (Tc1), dan aksesi asal
Pangalengan (Te3). Berdasarkan potensi alelopatinya, anggota grup tersebut
memisah menjadi empat kelompok. Seluruh aksesi asal Indramayu mengelompok
menjadi satu kelompok bersama aksesi S3 (aksesi asal Subang) dan K7 (aksesi
asal Karawang) yaitu kelompok A dengan karakter IR plumula rendah dan IR
radikula rendah. Aksesi K5 dan K6 mengelompok bersama aksesi Ta1 dengan
karakter IR plumula tinggi dan IR radikula sedang. Aksesi Td2 dan Tc1
mengelompok bersama aksesi Tf3 (asal Pangalengan) dengan karakter IR plumula
rendah dan IR radikula sedang. Aksesi Te3 mengelompok bersama aksesi K9 dan
C4 dengan karakter IR plumula sedang dan IR radikula rendah. Aksesi K3 (asal
Karawang) yang secara genetik berbeda dengan aksesi lainnya membentuk
kelompok sendiri dengan karakter IR plumula tinggi dan IR radikula rendah (IR
<10%). Aksesi Td3 yang secara genetik sama dengan aksesi K6, membentuk
kelompok sendiri dengan karakter IR plumula tinggi dan IR radikula tinggi
(Gambar 8 dan Tabel 19). Berdasarkan hasil tersebut terlihat bahwa keragaman
potensi alelopati aksesi gulma E. crus-galli dipengaruhi oleh genetik aksesi
tersebut.
Gambar 8. Dendogram kekerabatan aksesi gulma E. crus-galli dari berbagai habitat padi sawah di Jawa Barat berdasarkan potensi alelopati
Coefficient0.43 0.58 0.72 0.86 1.00
K7 S3 I5 I2 I1 K3 Te3 K9 C4 K6 K5 Ta1 Tf3 Td2 Tc1 Td3
A
B
D
F
C
E
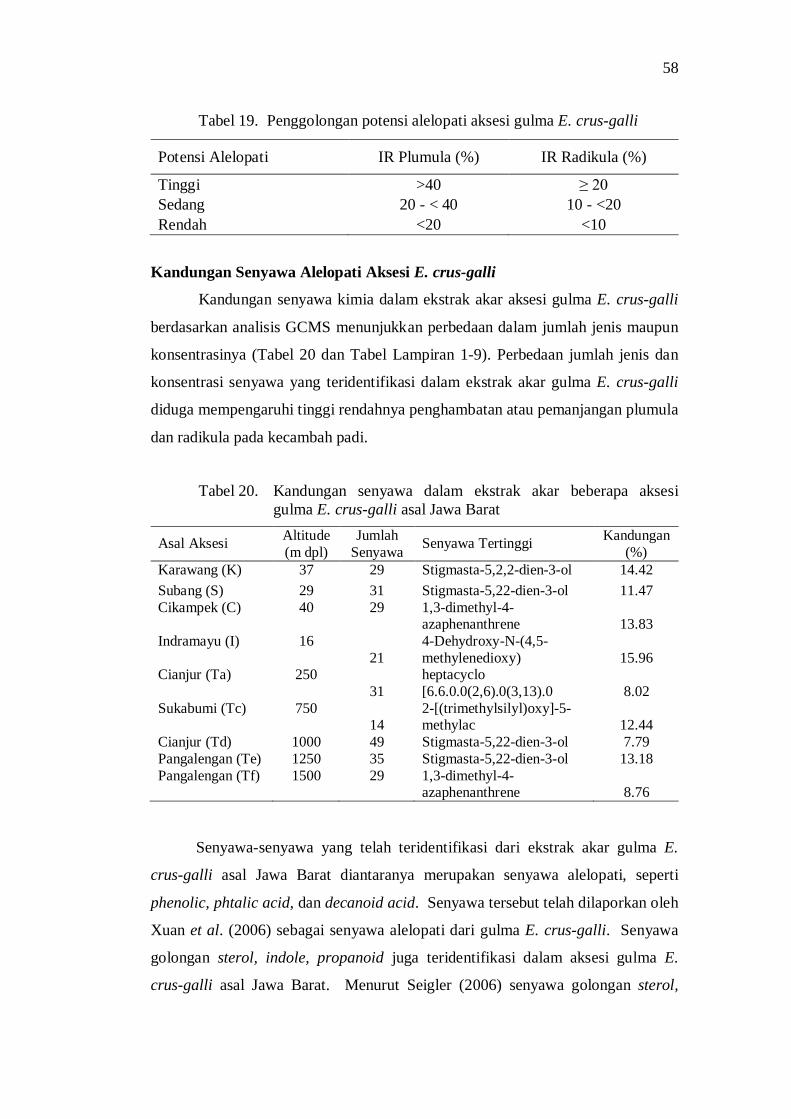
58
Tabel 19. Penggolongan potensi alelopati aksesi gulma E. crus-galli
Potensi Alelopati IR Plumula (%) IR Radikula (%)
Tinggi >40 ≥ 20 Sedang 20 - < 40 10 - <20 Rendah <20 <10
Kandungan Senyawa Alelopati Aksesi E. crus-galli
Kandungan senyawa kimia dalam ekstrak akar aksesi gulma E. crus-galli
berdasarkan analisis GCMS menunjukkan perbedaan dalam jumlah jenis maupun
konsentrasinya (Tabel 20 dan Tabel Lampiran 1-9). Perbedaan jumlah jenis dan
konsentrasi senyawa yang teridentifikasi dalam ekstrak akar gulma E. crus-galli
diduga mempengaruhi tinggi rendahnya penghambatan atau pemanjangan plumula
dan radikula pada kecambah padi.
Tabel 20. Kandungan senyawa dalam ekstrak akar beberapa aksesi
gulma E. crus-galli asal Jawa Barat
Asal Aksesi Altitude (m dpl)
Jumlah Senyawa Senyawa Tertinggi Kandungan
(%) Karawang (K) 37 29 Stigmasta-5,2,2-dien-3-ol 14.42 Subang (S) 29 31 Stigmasta-5,22-dien-3-ol 11.47 Cikampek (C) 40 29 1,3-dimethyl-4-
azaphenanthrene 13.83 Indramayu (I) 16
21 4-Dehydroxy-N-(4,5-methylenedioxy) 15.96
Cianjur (Ta) 250 31
heptacyclo [6.6.0.0(2,6).0(3,13).0 8.02
Sukabumi (Tc) 750 14
2-[(trimethylsilyl)oxy]-5-methylac 12.44
Cianjur (Td) 1000 49 Stigmasta-5,22-dien-3-ol 7.79 Pangalengan (Te) 1250 35 Stigmasta-5,22-dien-3-ol 13.18 Pangalengan (Tf) 1500 29 1,3-dimethyl-4-
azaphenanthrene 8.76 Senyawa-senyawa yang telah teridentifikasi dari ekstrak akar gulma E.
crus-galli asal Jawa Barat diantaranya merupakan senyawa alelopati, seperti
phenolic, phtalic acid, dan decanoid acid. Senyawa tersebut telah dilaporkan oleh
Xuan et al. (2006) sebagai senyawa alelopati dari gulma E. crus-galli. Senyawa
golongan sterol, indole, propanoid juga teridentifikasi dalam aksesi gulma E.
crus-galli asal Jawa Barat. Menurut Seigler (2006) senyawa golongan sterol,

59
indole, dan propanoid merupakan senyawa alelopati. Senyawa alelopati tersebut
diduga mempengaruhi pertumbuhan dan perkembangan serta produksi tanaman
padi di lapangan.
Pembahasan
Aksesi gulma E. crus-galli asal Jawa Barat memiliki perbedaan potensi
alelopati yang ditunjukkan oleh adanya penghambatan ataupun peningkatan
panjang plumula dan radikula kecambah padi. Penghambatan pajang plumula dan
radikula disebabkan oleh zat alelopati yang ada dalam ekstrak akar gulma yang
bersifat sebagai penghambat tumbuh. Hasil ini sejalan dengan penelitian
Yamamoto et al. (1999) bahwa selama perkecambahan dan pertumbuhan awal,
gulma E. crus-galli dapat menghambat beberapa tanaman uji, termasuk tanaman
padi. Salah satu senyawa alelopati yang teridentifikasi yaitu p-hydroxymandelic
acid pada konsentrasi 59.5-178 µM sangat kuat menghambat pertumbuhan
tanaman padi.
Pemanjangan plumula dan radikula yang terjadi pada beberapa aksesi
gulma E. crus-galli asal Jawa Barat diduga disebabkan oleh kurangnya
konsentrasi zat alelopati sehingga tidak mampu memberikan efek penghambatan
plumula dan radikula ataupun memang terdapat zat pemacu pertumbuhan yang
bekerja pada konsentrasi rendah, sehingga plumula dan radikula lebih panjang
dibandingkan terhadap kontrol. Fenomena ini perlu dikaji lebih lanjut untuk
menjelaskan dugaan tersebut.
Identifikasi terhadap senyawa kimia yang terkandung dalam ekstrak akar
gulma E. crus-galli menunjukkan adanya perbedaan jumlah jenis dan konsentrasi
tiap jenis senyawa dari masing-masing aksesi. Perbedaan jumlah jenis dan
konsentrasi tiap jenis senyawa tersebut diduga menjadi penyebab adanya
perbedaan dalam pemanjangan atau penghambatan plumula dan radikula
kecambah padi.
Hasil studi ini menunjukkan bahwa aksesi gulma E. crus-galli asal Jawa
Barat memiliki potensi alelopati. Beberapa senyawa potensial alelopati telah
teridentifikasi dari ekstrak akar aksesi gulma E. crus-galli asal Jawa Barat.
Senyawa-senyawa tersebut dilepaskan ke dalam lingkungan tumbuhnya selama

60
berkecambah dan pertumbuhan awal gulma. Menurut Xuan et al. (2006) senyawa
potensial alelopati yang dikeluarkan oleh gulma E. crus-galli menyebabkan
tanaman padi terhambat pertumbuhannya, tetapi senyawa tersebut kurang
menghambat pertumbuhan gulma itu sendiri. Pelepasan senyawa potensial
alelopati diduga ikut berperan dalam suksesnya gulma ini menjadi gulma invasive.
Adanya senyawa alelopati yang dilepaskan oleh gulma E. crus-galli di lapangan
pada perbedaan tahapan pertumbuhan dan interaksinya terhadap kondisi
lingkungan tumbuh tanaman sangat menarik untuk dikaji lebih jauh.
Kesimpulan
Aksesi gulma E. crus-galli asal Jawa Barat memiliki potensi alelopati
yang ditunjukkan dengan adanya penghambatan ataupun pemanjangan plumula
dan radikula kecambah padi. Penghambatan atau pemanjangan plumula dan
radikula dipengaruhi oleh jumlah jenis dan konsentrasi senyawa yang terkandung
dalam ekstrak akar tiap aksesi gulma. Senyawa alelopati yang teridentifikasi dari
ekstrak akar aksesi E. crus-galli diantaranya adalah golongan senyawa phenolic,
phtalic acid, decanoid acid, propanoid, quinon, indole dan sterol.
Berdasarkan potensi alelopatinya, setiap aksesi menunjukkan potensi
alelopati yang berbeda. Perbedaan potensi alelopati tersebut disebabkan oleh asal
habitat dan sifat genetiknya. Aksesi asal Karawang (K5, K6, K3) dan asal Cianjur
(Ta1) lebih menekan plumula sedangkan aksesi asal Pangalengan (Td3) lebih
menekan radikula.

PENGARUH AKSESI GULMA Echinochloa crus-galli TERHADAP PERTUMBUHAN DAN PRODUKSI PADI
ABSTRAK
Aksesi gulma E. crus-galli dari beberapa habitat padi sawah di Jawa Barat diduga memiliki potensi yang berbeda dalam menurunkan pertumbuhan dan produksi tanaman padi sawah. Penelitian bertujuan untuk mempelajari pengaruh beberapa aksesi gulma E. crus-galli asal Jawa Barat terhadap pertumbuhan dan produksi tanaman padi sawah pada percobaan pot di rumah kaca. Percobaan dilakukan dengan menggunakan rancangan acak kelompok dengan satu faktor dan tiga ulangan. Faktor yang dicobakan yaitu aksesi gulma E. crus-galli yang terdiri atas 5 aksesi terpilih (K3, K6, K9, Te3, Tf3) dan kontrol tanpa gulma. Satu satuan percobaan berupa pot berukuran diameter 30 cm dan tinggi 45 cm. Setiap pot ditanam 1 bibit padi dan 2 bibit gulma E. crus-galli. Hasil penelitian menunjukkan bahwa aksesi gulma E. crus-galli memiliki potensi yang berbeda dalam menurunkan pertumbuhan dan hasil tanaman padi. Aksesi K6 (Karawang) merupakan aksesi yang paling potensial dalam menurunkan pertumbuhan dan produksi padi. Potensi aksesi K6 dalam menurunkan pertumbuhan dan produksi tanaman padi sawah terkait dengan potensi alelopati dan karakter morfologinya yaitu panjang daun panjang (>60 cm), lebar daun sempit (<0.7 cm), sudut daun kecil (<16°), dan umur berbunga lambat (>16 MST). Kata kunci : aksesi Echinochloa crus-galli, pertumbuhan, produksi, padi.

62
THE EFFECT OF Echinochloa crus-galli (L.) Beauv. WEED ACCESSION ON GROWTH AND PRODUCTION
OF RICE PLANT
ABSTRACT
E. crus-galli weed accession from several habitats of lowland rice in West Java supposedly has a potential different in reducing the growth and production of rice plants. The research objective was to study the effect of some accessions of E. crus-galli weed from West Java on the growth and production of rice plants in greenhouse experiment. The experiments were conducted in randomized block design with one factor and three replications. The accession of E. crus-galli weed consisted of five elected accession (K3, K6, K9, Te3, TF3) and without weed as control. The experiment unis was pots (diameter 30 cm and a height of 40 cm). Each pot was planted one rice seedling and 2 E. crus-galli seedling. The results showed that the E. crus-galli weed accession had potential different in reducing growth and yield of rice plants. Accession K6 (Karawang) was the most potential accession in decreasing the growth and production of rice. Decreasing of the rice growth and production of K6 accession related to the allelophatic potential and morphological character of K6 accession i.e. lenght leaf (> 60 cm), width leaf (<0.7 cm), leaf angle (<16 °), and flowering age (> 16 MST
).
Keywords: Echinochloa crus-galli accession, growth, production, rice
.

63
Pendahuluan
Upaya untuk meningkatkan produksi padi dipengaruhi oleh beberapa hal
diantaranya luas lahan yang menurun akibat konversi lahan, menurunnya
kesuburan tanah dan serangan organisme pengganggu tanaman (OPT). Salah satu
OPT yang dapat menurunkan produksi tanaman padi yaitu gulma. Kerugian
akibat gulma di negara yang sedang berkembang tidak saja tinggi, tetapi juga
mempengaruhi persediaan pangan dunia.
Gulma merupakan tumbuhan yang tumbuh pada tempat yang tidak
dikehendaki. Gulma yang tumbuh di sekitar tanaman yang dibudidayakan dapat
menghambat pertumbuhan dan menekan hasil akhir. Salah satu jenis gulma utama
pada lahan sawah yang dapat menurunkan produksi tanaman padi adalah gulma
Echinochloa crus-galli (L.) Beauv. Gulma ini dapat menurunkan hasil padi
sampai 72% (Lubigan dan Vega dalam Suardi dan Pane 1983).
Gulma dapat menurunkan pertumbuhan dan produksi tanaman melalui
mekanisme kompetisi, alelopati, dan alelomediasi. Kompetisi dapat terjadi antara
gulma dengan tanaman padi dalam memperebutkan sumber daya yang terbatas,
yaitu air, unsur hara, dan sarana tumbuh lainnya. Mekanisme alelopati terjadi
dengan adanya zat alelopati yang bersifat menghambat yang dikeluarkan oleh
gulma ke dalam lingkungan dan mempengaruhi pertumbuhan dan produksi
tanaman padi. Alelomediasi terjadi karena tanaman padi menjadi inang bagi
hama dan penyakit tanaman padi sehingga menyebabkan penurunan pertumbuhan
dan produksi tanaman padi.
Aksesi gulma E. crus-galli dari beberapa habitat padi sawah di Jawa Barat
menunjukkan potensi alelopati yang berbeda dalam menghambat perkecambahan
tanaman padi yang ditunjukkan dengan perbedaan dalam penghambatan plumula
dan radikula. Perbedaan potensi alelopati ini diduga menyebabkan tinggi
rendahnya penurunan pertumbuhan dan produksi tanaman padi. Penelitian
bertujuan mempelajari pengaruh beberapa aksesi gulma E. crus-galli asal Jawa
Barat terhadap pertumbuhan dan produksi tanaman padi sawah pada percobaan
pot di rumah kaca.

64
Bahan dan Metode
Penelitian dilaksanakan di rumah kaca Kebun Percobaan IPB, Cikabayan,
Bogor mulai Februari 2007 hingga Desember 2007. Bahan tanaman berupa biji
aksesi E. crus-galli asal di Jawa Barat, benih padi varietas IR-64, pupuk Urea,
KCl, SP-36, furadan 3G, aquadest dan media tanah sawah jenis latosol. Alat yang
digunakan antara lain pot, peralatan tanam, timbangan analitik, dan oven.
Percobaan dilakukan dengan menggunakan rancangan acak kelompok
dengan satu faktor dan tiga ulangan. Faktor yang dicobakan yaitu aksesi gulma E.
crus-galli yang terdiri atas 5 aksesi terpilih yaitu aksesi K3, K6, K9, Te3, Tf3 dan
kontrol tanpa gulma. Satu satuan percobaan berupa pot berukuran 30-40
(diameter-tinggi). Setiap pot ditanam 1 bibit padi dan 2 bibit gulma E. crus-galli.
Media tanah sawah latosol darmaga dikeringanginkan dan dihaluskan
kemudian dimasukkan ke dalam pot sebanyak 9 kg per pot. Tanah dilumpurkan
dengan cara menyiramkan air hingga jenuh dan diaduk. Benih padi IR-64 dan biji
E. crus-galli disemai dalam bak semai. Bibit padi dan gulma E. crus-galli
dipindahtanam ke dalam pot pada saat 2 minggu setelah semai. Bibit padi
ditanam tepat di tengah-tengah pot sebanyak satu bibit per pot dan bibit gulma E.
crus-galli sebanyak dua bibit per pot yang ditanam di sisi kanan dan kiri tanaman
padi dengan jarak sekitar 7 cm dari tanaman padi.
Penyiraman tanaman dilakukan setiap dua hari sekali dengan ketinggian
genangan 5 cm. Pemupukan dilakukan pada saat 0, 4, dan 6 minggu setelah tanam
(MST) dengan dosis pupuk 1.88 g urea/pot, 1.25 g SP-36/pot, dan 1.25 g KCl/pot.
Pengendalian gulma dilakukan secara manual dengan cara mencabut gulma selain
gulma E. crus-galli yang tumbuh di dalam pot. Panen dilakukan pada saat 80 %
malai padi sudah menguning.
Peubah yang diamati antara lain tinggi tanaman, panjang dan lebar daun
ke-3 dari atas, jumlah daun, jumlah anakan, jumlah anakan produktif, panjang dan
lebar daun bendera, panjang akar pada saat panen, panjang malai, kepadatan
malai, produksi gabah, bobot gabah isi dan hampa, dan persentase gabah hampa,
bobot biomassa akar dan tajuk, dan bobot 100 butir gabah kering. Data hasil
pengamatan dianalisis menggunakan analisis ragam dengan uji lanjut Duncan’s
Multiple Range Test pada taraf 5%.
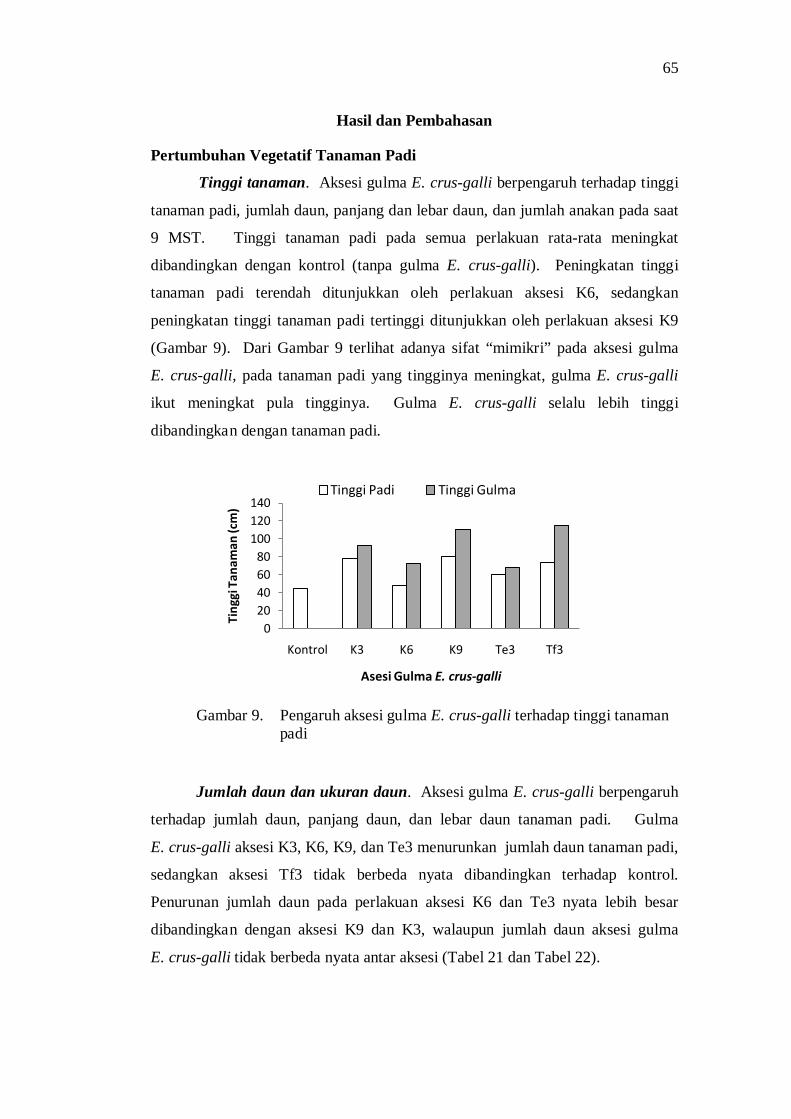
65
Hasil dan Pembahasan
Pertumbuhan Vegetatif Tanaman Padi
Tinggi tanaman. Aksesi gulma E. crus-galli berpengaruh terhadap tinggi
tanaman padi, jumlah daun, panjang dan lebar daun, dan jumlah anakan pada saat
9 MST. Tinggi tanaman padi pada semua perlakuan rata-rata meningkat
dibandingkan dengan kontrol (tanpa gulma E. crus-galli). Peningkatan tinggi
tanaman padi terendah ditunjukkan oleh perlakuan aksesi K6, sedangkan
peningkatan tinggi tanaman padi tertinggi ditunjukkan oleh perlakuan aksesi K9
(Gambar 9). Dari Gambar 9 terlihat adanya sifat “mimikri” pada aksesi gulma
E. crus-galli, pada tanaman padi yang tingginya meningkat, gulma E. crus-galli
ikut meningkat pula tingginya. Gulma E. crus-galli selalu lebih tinggi
dibandingkan dengan tanaman padi.
Gambar 9. Pengaruh aksesi gulma E. crus-galli terhadap tinggi tanaman padi
Jumlah daun dan ukuran daun. Aksesi gulma E. crus-galli berpengaruh
terhadap jumlah daun, panjang daun, dan lebar daun tanaman padi. Gulma
E. crus-galli aksesi K3, K6, K9, dan Te3 menurunkan jumlah daun tanaman padi,
sedangkan aksesi Tf3 tidak berbeda nyata dibandingkan terhadap kontrol.
Penurunan jumlah daun pada perlakuan aksesi K6 dan Te3 nyata lebih besar
dibandingkan dengan aksesi K9 dan K3, walaupun jumlah daun aksesi gulma
E. crus-galli tidak berbeda nyata antar aksesi (Tabel 21 dan Tabel 22).
020406080
100120140
Kontrol K3 K6 K9 Te3 Tf3
Ting
gi T
anam
an (c
m)
Asesi Gulma E. crus-galli
Tinggi Padi Tinggi Gulma
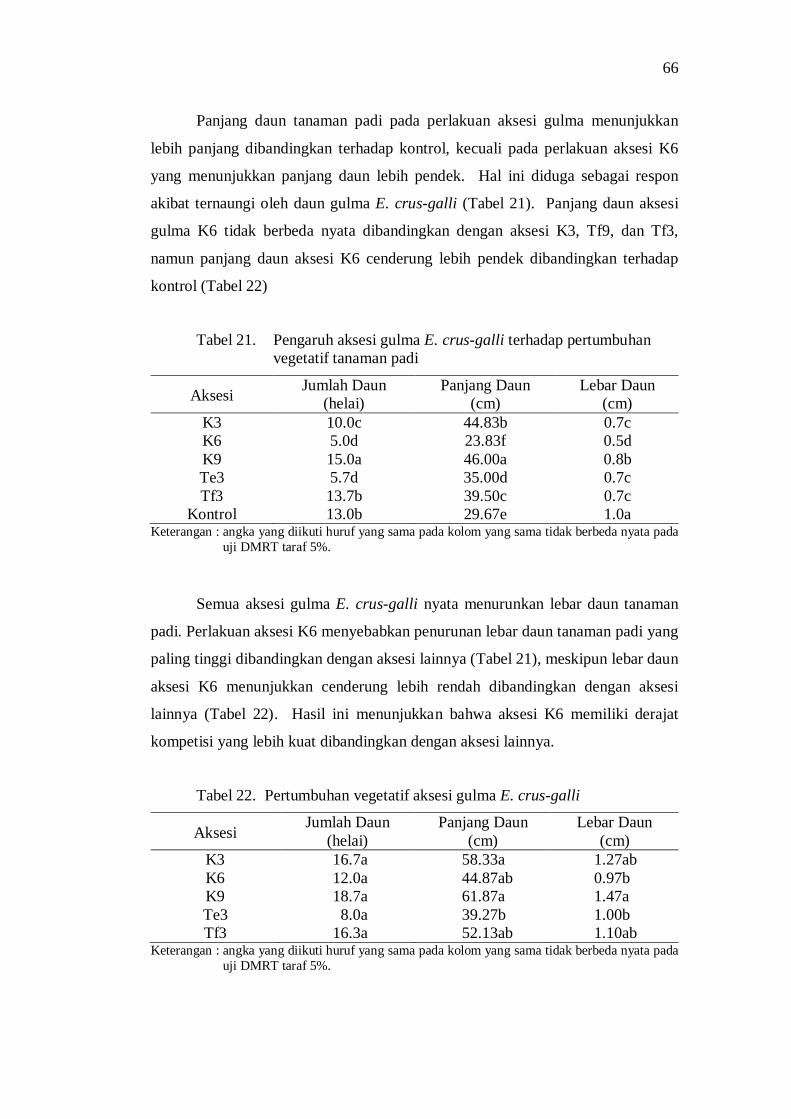
66
Panjang daun tanaman padi pada perlakuan aksesi gulma menunjukkan
lebih panjang dibandingkan terhadap kontrol, kecuali pada perlakuan aksesi K6
yang menunjukkan panjang daun lebih pendek. Hal ini diduga sebagai respon
akibat ternaungi oleh daun gulma E. crus-galli (Tabel 21). Panjang daun aksesi
gulma K6 tidak berbeda nyata dibandingkan dengan aksesi K3, Tf9, dan Tf3,
namun panjang daun aksesi K6 cenderung lebih pendek dibandingkan terhadap
kontrol (Tabel 22)
Tabel 21. Pengaruh aksesi gulma E. crus-galli terhadap pertumbuhan vegetatif tanaman padi
Aksesi Jumlah Daun (helai)
Panjang Daun (cm)
Lebar Daun (cm)
K3 10.0c 44.83b 0.7c K6 5.0d 23.83f 0.5d K9 15.0a 46.00a 0.8b Te3 5.7d 35.00d 0.7c Tf3 13.7b 39.50c 0.7c
Kontrol 13.0b 29.67e 1.0a Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada
uji DMRT taraf 5%.
Semua aksesi gulma E. crus-galli nyata menurunkan lebar daun tanaman
padi. Perlakuan aksesi K6 menyebabkan penurunan lebar daun tanaman padi yang
paling tinggi dibandingkan dengan aksesi lainnya (Tabel 21), meskipun lebar daun
aksesi K6 menunjukkan cenderung lebih rendah dibandingkan dengan aksesi
lainnya (Tabel 22). Hasil ini menunjukkan bahwa aksesi K6 memiliki derajat
kompetisi yang lebih kuat dibandingkan dengan aksesi lainnya.
Tabel 22. Pertumbuhan vegetatif aksesi gulma E. crus-galli
Aksesi Jumlah Daun
(helai) Panjang Daun
(cm) Lebar Daun
(cm) K3 16.7a 58.33a 1.27ab K6 12.0a 44.87ab 0.97b K9 18.7a 61.87a 1.47a Te3 8.0a 39.27b 1.00b Tf3 16.3a 52.13ab 1.10ab
Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
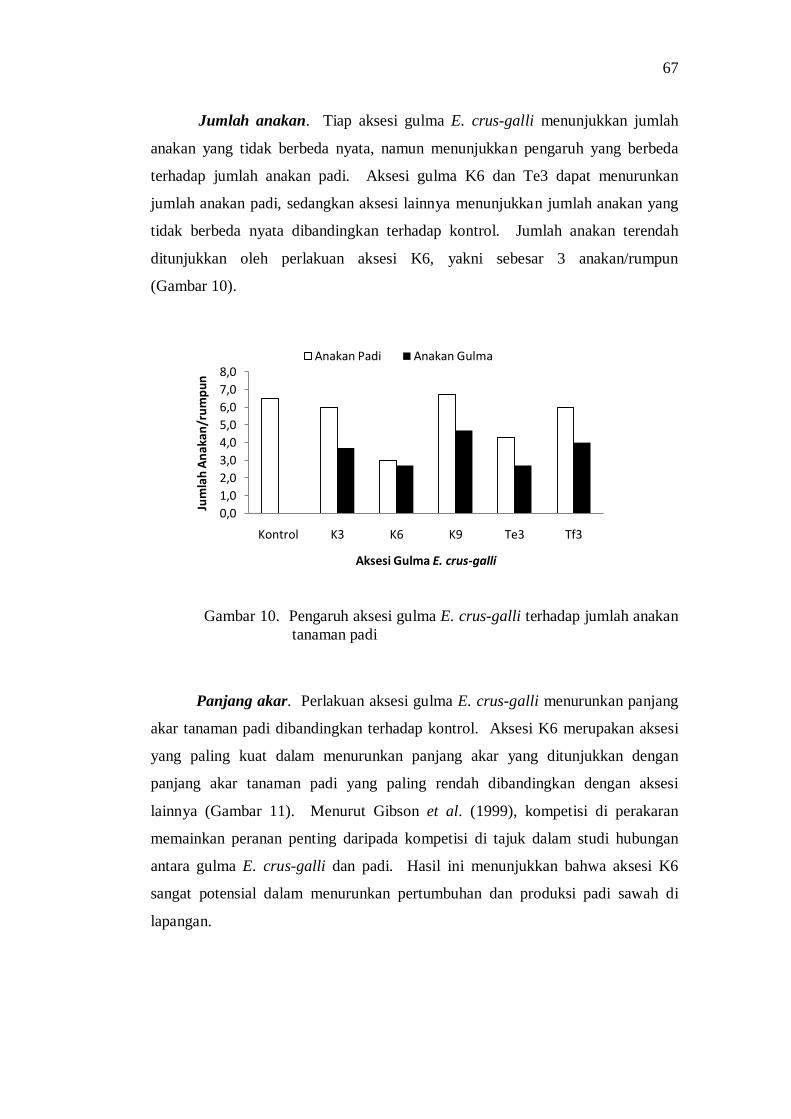
67
Jumlah anakan. Tiap aksesi gulma E. crus-galli menunjukkan jumlah
anakan yang tidak berbeda nyata, namun menunjukkan pengaruh yang berbeda
terhadap jumlah anakan padi. Aksesi gulma K6 dan Te3 dapat menurunkan
jumlah anakan padi, sedangkan aksesi lainnya menunjukkan jumlah anakan yang
tidak berbeda nyata dibandingkan terhadap kontrol. Jumlah anakan terendah
ditunjukkan oleh perlakuan aksesi K6, yakni sebesar 3 anakan/rumpun
(Gambar 10).
Gambar 10. Pengaruh aksesi gulma E. crus-galli terhadap jumlah anakan tanaman padi
Panjang akar. Perlakuan aksesi gulma E. crus-galli menurunkan panjang
akar tanaman padi dibandingkan terhadap kontrol. Aksesi K6 merupakan aksesi
yang paling kuat dalam menurunkan panjang akar yang ditunjukkan dengan
panjang akar tanaman padi yang paling rendah dibandingkan dengan aksesi
lainnya (Gambar 11). Menurut Gibson et al. (1999), kompetisi di perakaran
memainkan peranan penting daripada kompetisi di tajuk dalam studi hubungan
antara gulma E. crus-galli dan padi. Hasil ini menunjukkan bahwa aksesi K6
sangat potensial dalam menurunkan pertumbuhan dan produksi padi sawah di
lapangan.
0,01,02,03,04,05,06,07,08,0
Kontrol K3 K6 K9 Te3 Tf3
Jum
lah
Ana
kan/
rum
pun
Aksesi Gulma E. crus-galli
Anakan Padi Anakan Gulma
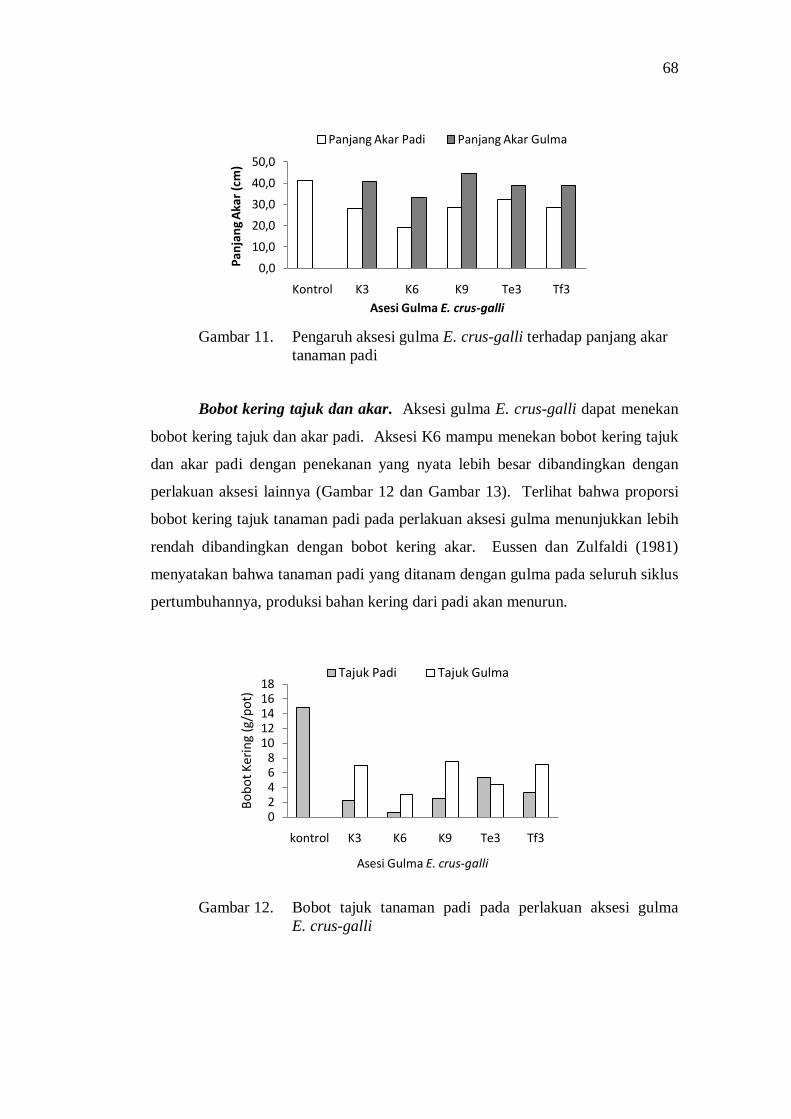
68
Gambar 11. Pengaruh aksesi gulma E. crus-galli terhadap panjang akar tanaman padi
Bobot kering tajuk dan akar. Aksesi gulma E. crus-galli dapat menekan
bobot kering tajuk dan akar padi. Aksesi K6 mampu menekan bobot kering tajuk
dan akar padi dengan penekanan yang nyata lebih besar dibandingkan dengan
perlakuan aksesi lainnya (Gambar 12 dan Gambar 13). Terlihat bahwa proporsi
bobot kering tajuk tanaman padi pada perlakuan aksesi gulma menunjukkan lebih
rendah dibandingkan dengan bobot kering akar. Eussen dan Zulfaldi (1981)
menyatakan bahwa tanaman padi yang ditanam dengan gulma pada seluruh siklus
pertumbuhannya, produksi bahan kering dari padi akan menurun.
Gambar 12. Bobot tajuk tanaman padi pada perlakuan aksesi gulma E. crus-galli
0,0
10,0
20,0
30,0
40,0
50,0
Kontrol K3 K6 K9 Te3 Tf3
Panj
ang
Aka
r (cm
)
Asesi Gulma E. crus-galli
Panjang Akar Padi Panjang Akar Gulma
02468
1012141618
kontrol K3 K6 K9 Te3 Tf3
Bobo
t Ker
ing
(g/p
ot)
Asesi Gulma E. crus-galli
Tajuk Padi Tajuk Gulma
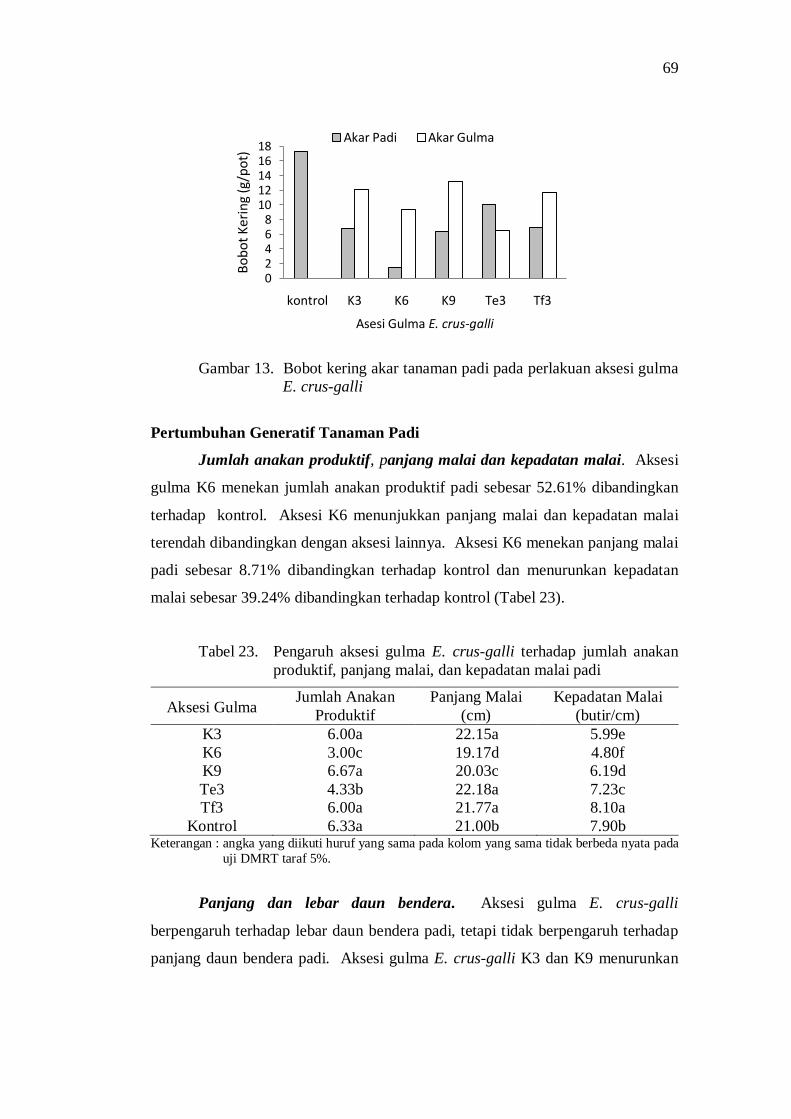
69
Gambar 13. Bobot kering akar tanaman padi pada perlakuan aksesi gulma E. crus-galli
Pertumbuhan Generatif Tanaman Padi
Jumlah anakan produktif, panjang malai dan kepadatan malai. Aksesi
gulma K6 menekan jumlah anakan produktif padi sebesar 52.61% dibandingkan
terhadap kontrol. Aksesi K6 menunjukkan panjang malai dan kepadatan malai
terendah dibandingkan dengan aksesi lainnya. Aksesi K6 menekan panjang malai
padi sebesar 8.71% dibandingkan terhadap kontrol dan menurunkan kepadatan
malai sebesar 39.24% dibandingkan terhadap kontrol (Tabel 23).
Tabel 23. Pengaruh aksesi gulma E. crus-galli terhadap jumlah anakan
produktif, panjang malai, dan kepadatan malai padi
Aksesi Gulma Jumlah Anakan
Produktif Panjang Malai
(cm) Kepadatan Malai
(butir/cm) K3 6.00a 22.15a 5.99e K6 3.00c 19.17d 4.80f K9 6.67a 20.03c 6.19d Te3 4.33b 22.18a 7.23c Tf3 6.00a 21.77a 8.10a
Kontrol 6.33a 21.00b 7.90b Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada
uji DMRT taraf 5%.
Panjang dan lebar daun bendera. Aksesi gulma E. crus-galli
berpengaruh terhadap lebar daun bendera padi, tetapi tidak berpengaruh terhadap
panjang daun bendera padi. Aksesi gulma E. crus-galli K3 dan K9 menurunkan
02468
1012141618
kontrol K3 K6 K9 Te3 Tf3
Bobo
t Ker
ing
(g/p
ot)
Asesi Gulma E. crus-galli
Akar Padi Akar Gulma
70
lebar daun bendera, sedangkan aksesi lainnya menunjukkan lebar daun bendera
yang tidak berbeda nyata dibandingkan terhadap kontrol (Tabel 24).
Tabel 24. Pengaruh aksesi gulma E. crus-galli terhadap panjang dan
lebar daun bendera tanaman padi
Aksesi Gulma Panjang Daun Bendera
(cm) Lebar Daun Bendera
(cm) K3 30.25a 1.09c K6 27.68a 1.15bc K9 27.88a 1.10c Te3 25.93a 1.13bc Tf3 30.94a 1.24a
Kontrol 27.94a 1.18ab Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada
uji lanjut DMRT taraf 5%.
Produksi gabah dan mutu hasil. Aksesi gulma E. crus-galli berpengaruh
terhadap bobot gabah kering total, bobot gabah isi, dan bobot gabah hampa.
Aksesi K6 menurunkan bobot gabah total, gabah isi dan gabah hampa padi
berturut-turut sebesar 78.69%, 81.91% dan 66.27% dibandingkan terhadap
kontrol. Aksesi gulma E. crus-galli tidak berpengaruh terhadap bobot 100 butir
gabah padi (Tabel 25).
Tabel 25. Pengaruh aksesi gulma E. crus-galli terhadap bobot gabah
total, bobot gabah isi, bobot gabah hampa, dan bobot 100 butir
Aksesi Bobot Gabah (g/pot) Bobot 100 Butir
(g) Total Isi Hampa K3 4.88d 3.70d 1.18d 1.96a K6 2.61f 1.76e 0.85e 1.58a K9 6.47c 5.25c 1.22c 2.03a Te3 7.84b 6.48b 1.37b 1.87a Tf3 4.40e 3.67d 0.73f 1.63a
Kontrol 12.25a 9.73a 2.52a 1.63a Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada
uji lanjut DMRT taraf 5%.
Pembahasan
Aksesi gulma E. crus-galli asal Jawa Barat memiliki potensi yang berbeda
dalam menurunkan pertumbuhan dan produksi tanaman padi di dalam pot pada
percobaan rumah kaca. Hasil percobaan menunjukkan bahwa aksesi gulma E.
crus-galli menyebabkan etiolasi pada tanaman padi sehingga tinggi tanaman padi

71
meningkat jika dibandingkan dengan tanaman padi tanpa gulma (Gambar 9).
Aksesi gulma E. crus-galli menyebabkan penurunan jumlah daun dan ukuran
daun tanaman padi (Tabel 21) yang diikuti dengan penurunan jumlah anakan
(Gambar 10) dan bobot tajuk tanaman padi (Gambar 12). Aksesi gulma E. crus-
galli juga menyebabkan penurunan panjang akar (Gambar 11) dan bobot akar
tanaman padi (Gambar 13). Aksesi gulma E. crus-galli menurunkan jumlah
anakan produktif, panjang malai, dan kepadatan malai (Tabel 23), serta
menurunkan produksi gabah (Tabel 25).
Perbedaan penurunan pertumbuhan dan produksi padi oleh aksesi gulma
diduga terkait dengan potensi alelopati tiap aksesi gulma E. crus-galli dan searah
dengan penghambatan plumula dan radikula. Aksesi K6 dengan tingkat
penghambatan plumula tinggi dan tingkat penghambatan radikula sedang
menyebabkan penurunan jumlah daun, panjang dan lebar daun, panjang akar,
bobot tajuk dan bobot akar yang lebih besar dibandingkan dengan aksesi lainnya.
Aksesi K3 yang memiliki potensi penghambatan plumula tinggi dan
penghambatan radikula rendah menyebabkan penurunan bobot tajuk yang hampir
sama dengan aksesi K6, tetapi penurunan bobot akar lebih rendah dibandingkan
dengan aksesi K6. Aksesi Tf3 dengan potensi penghambatan plumula rendah dan
penghambatan radikula sedang serta aksesi K9 dan Te3 dengan tingkat
penghambatan plumula sedang dan tingkat penghambatan radikula rendah
menunjukkan penurunan pertumbuhan dan produksi padi yang lebih rendah
dibandingkan dengan aksesi K6.
Potensi penurunan pertumbuhan dan produksi tanaman padi lebih
disebabkan oleh sifat genetik dari aksesi gulma E. crus-galli dibandingkan dengan
asal aksesi. Dalam kasus aksesi K3, K6, dan Tf3, aksesi tersebut secara genetik
berbeda dan lokasi asal aksesi berbeda. Ketiga aksesi tersebut memiliki potensi
penghambatan plumula dan radikula yang berbeda dan menyebabkan penurunan
pertumbuhan dan produksi padi juga berbeda. Aksesi K9 dan Te3 walaupun asal
lokasi berbeda, tetapi secara genetik sama. Aksesi K9 dan Te3 memiliki potensi
penghambatan plumula dan radikula yang sama sehingga menyebabkan
penurunan pertumbuhan dan produksi padi yang tidak berbeda.

72
Penurunan pertumbuhan dan produksi tanaman padi selain disebabkan
oleh potensi alelopati aksesi gulma E. crus-galli diduga juga terkait dengan
karakter morfologinya. Aksesi K6 dengan karakter panjang daun yang panjang
(tidak berbeda dengan aksesi lainnya), daun sempit (<0.7 cm), sudut daun kecil
(<16°), dan umur berbunga lambat diduga memiliki daya kompetisi di atas tanah
yang kuat. Daun tidak saling menaungi, sehingga proses fotosintesis lebih baik.
Selain itu, dengan masa vegetatif yang lebih panjang, maka masa kompetisi
dengan tanaman padi lebih lama. Apabila dikaitkan dengan lokasi Karawang
yang intensif dalam praktek budidaya tanaman padi, diduga aksesi ini memiliki
kemampuan adaptasi dan plastisitas fenotipik pada kondisi lingkungan yang
berbeda, sehingga memiliki kemampuan yang kuat dalam menurunkan
pertumbuhan dan produksi tanaman padi.
Kesimpulan
Aksesi gulma E. crus-galli menunjukkan potensi yang berbeda dalam
menurunkan pertumbuhan dan hasil tanaman padi. Perbedaan penurunan
pertumbuhan dan produksi padi disebabkan oleh perbedaan potensi alelopati
aksesi gulma E. crus-galli yang disebabkan perbedaan faktor genetik.
Aksesi K6 (Karawang) merupakan aksesi yang paling potensial dalam
menurunkan pertumbuhan dan produksi padi. Aksesi K6 yang memiliki potensi
penghambatan plumula tinggi dan penghambatan radikula sedang menyebabkan
penurunan jumlah daun, panjang dan lebar daun, jumlah anakan, panjang akar,
bobot kering tajuk dan akar, jumlah anakan produktif, panjang dan kepadatan
malai, bobot gabah total dan gabah isi yang lebih tinggi dibandingkan dengan
aksesi lainnya.
Potensi aksesi K6 dalam menurunkan pertumbuhan dan produksi tanaman
padi sawah juga terkait dengan karakter morfologi aksesi K6 yaitu panjang daun
panjang (>60 cm), lebar daun sempit (<0.7 cm), sudut daun kecil (<16°), dan
umur berbunga lambat (>16 MST).

PENGARUH AKSESI DAN KEPADATAN POPULASI GULMA Echinochloa crus-galli TERHADAP PERTUMBUHAN
DAN PRODUKSI PADI
ABSTRAK
Penelitian bertujuan untuk mempelajari pengaruh aksesi dan tingkat populasi gulma E. crus-galli terhadap pertumbuhan dan produksi tanaman padi sawah. Percobaan dilakukan dengan menggunakan rancangan petak terbagi dalam rancangan acak kelompok dengan tiga ulangan. Aksesi gulma E. crus-galli sebagai petak utama terdiri atas tiga aksesi yaitu aksesi Pangalengan (Tf3, dataran tinggi, 1500 m dpl), aksesi Sukabumi (Tc1, dataran sedang, 500 m dpl), dan aksesi Karawang (K6, dataran rendah , 37 m dpl). Populasi E. crus-galli sebagai anak petak terdiri atas empat taraf, yaitu: 0, 4, 8, dan 12 gulma E. crus-galli/m2. Hasil penelitian menunjukkan bahwa perbedaan aksesi gulma E. crus-galli menyebabkan perbedaan pertumbuhan dan produksi tanaman padi. Aksesi gulma E. crus-galli berpengaruh terhadap pertumbuhan dan produksi tanaman padi hibrida. Memperkuat hasil percobaan rumah kaca, aksesi asal Karawang (K6) memiliki derajat kompetisi yang lebih kuat dibandingkan dengan aksesi Sukabumi dan Pangalengan terhadap pertumbuhan tanaman padi hibrida. Kepadatan populasi gulma E. crus-galli berpengaruh terhadap pertumbuhan dan produksi tanaman padi hibrida. Semakin tinggi populasi gulma E. crus-galli, pengaruh kompetisi terhadap tanaman padi semakin besar. Interaksi antara aksesi dan gulma E. crus-galli berpengaruh terhadap tinggi tanaman, bobot kering akar dan tajuk, kepadatan malai dan bobot 1 000 butir tanaman padi hibrida. Gulma E. crus-galli harus dikendalikan pada saat populasi gulma mencapai 1.9 gulma/m2 untuk menyelamatkan kehilangan hasil sebesar 10%. Penggunaan benih padi yang bebas dari benih gulma E.crus-galli sangat dianjurkan untuk mencegah penyebaran aksesi gulma E. crus-galli. Kata kunci : aksesi Echinocloa crus-galli, population, growth, production, hybrid rice.

74
THE EFFECT OF ACCESSION AND POPULATION DENSITY OF Echinochloa crus-galli WEED ON GROWTH AND RICE
PRODUCTION
ABSTRACT
The research objective was to study the effect of accession and populations density of E. crus-galli weed on the growth and production of rice plants. The research was conducted in a split plot design with three replications. E. crus-galli accession as the main plots consisted of three accessions i.e. accession of Pangalengan (Tf3, high altitude, 1500 m asl), Sukabumi (Tc1, medium altitude, 500 m asl), and Karawang (K6, lowland altitude, 37 m asl). Population density of E. crus-galli as a subplot consisted of four levels, i.e. : 0, 4, 8, and 12 E. crus-galli/m2. The results showed that accession of E. crus-galli weed affected growth and production of rice plants. Strengthen the results of greenhouse experiments, accession from Karawang (K6) had much stronger competition degree than the Sukabumi and Pangalengan accession on the growth of hybrid rice plants. Weed population densities of E. crus-galli had significantly effect on growth and production of hybrid rice. The influence of competition on rice growth was higher when the weed population density increased. The interaction between accessions and E. crus-galli weed effect on plant height, dry weight of root and shoot, panicle density, and 1000 grain weight of rice hybrids. E. crus-galli weed must be controlled at 1.9 weed/m2 to save 10% yield loss. The use of rice seed free of E.crus-galli seed is highly recommended to prevent the spread of E. crus-galli weed accession.
Keywords : Echinocloa crus-galli accession, population, growth, production, hybrid rice.

75
Pendahuluan
Gulma menurunkan pertumbuhan dan produksi tanaman dengan
penurunan mencapai 30–60%, bahkan bila tidak dilakukan upaya pengendalian
dapat menyebabkan gagal panen (Singh et al. 1996). Kehadiran gulma pada
pertanaman padi sawah juga menyebabkan peningkatan biaya pengendalian
sehingga menurunkan pendapatan petani (Tungate et al. 2007).
Gulma E. crus-galli merupakan gulma pada tanaman padi sawah yang
memiliki adaptasi luas, dijumpai di dataran rendah maupun dataran tinggi sampai
ketinggian 2 500 m dpl, pertumbuhan cepat, produksi biji tinggi, dormansi benih
lama, dan penyebaran sangat pesat (Caton et al. 2004). Pada kondisi tanah kering
E. crus-galli tumbuh pendek dan memiliki jumlah anakan dan biji yang sedikit,
sedangkan pada kondisi tanah lembab dengan kandungan nitrogen tinggi E. crus-
galli tumbuh sangat baik dan serempak (Waterhouse 1994).
Perbedaan aksesi gulma E. crus-galli asal Jawa Barat menunjukkan
perbedaan dalam menurunkan pertumbuhan dan hasil produksi tanaman padi.
Aksesi K6 asal Karawang pada percobaan di rumah kaca menunjukkan potensi
penurunan pertumbuhan dan hasil produksi tanaman padi yang paling besar
dibandingkan dengan aksesi lainnya. Penurunan pertumbuhan dan hasil produksi
tanaman padi tersebut diduga terkait dengan potensi alelopati aksesi K6 yang
menunjukkan penghambatan plumula tinggi dan penghambatan radikula sedang.
Penurunan pertumbuhan dan hasil produksi tanaman padi di lapangan
dipengaruhi oleh kekuatan kompetisi dan adanya zat alelopati gulma E. crus-galli
yang dikeluarkan ke lingkungan tumbuh. Kekuatan kompetisi di lapangan
dipengaruhi oleh tingkat populasi gulma E. crus-galli. Menurut Chin (2001)
semakin tinggi tingkat populasi gulma, maka kompetisi terhadap tanaman padi
semakin meningkat. Populasi 25 gulma E. crus-galli/m2 menyebabkan
kehilangan hasil panen hingga 50%. Penelitian bertujuan untuk mempelajari
pengaruh aksesi dan tingkat populasi gulma E. crus-galli terhadap pertumbuhan
dan produksi tanaman padi sawah.

76
Bahan dan Metode
Penelitian dilaksanakan di lahan sawah Desa Parakan, Kecamatan Ciomas,
Kabupaten Bogor (250 m dpl) dengan jenis tanah latosol mulai bulan Desember
2010 hingga bulan Mei 2011. Bahan tanaman yang digunakan antara lain benih
padi hibrida varietas SL 8 SHS, biji gulma E. crus-galli (aksesi Karawang (K6),
Sukabumi (Tc1), dan Pangalengan (Tf3). Peralatan yang digunakan antara lain
GPS, oven, neraca dan peralatan budidaya tanaman.
Percobaan dilakukan dengan menggunakan rancangan petak terbagi (split
plot design) dalam rancangan acak kelompok. Percobaan terdiri atas dua faktor
dengan tiga ulangan. Aksesi gulma E. crus-galli sebagai petak utama terdiri atas
tiga aksesi yaitu aksesi Pangalengan (Tf3, dataran tinggi, 1500 m dpl), aksesi
Sukabumi (Tc1, dataran sedang, 500 m dpl), dan aksesi Karawang (K6, dataran
rendah, 37 m dpl). Populasi gulma E. crus-galli sebagai anak petak terdiri atas
empat taraf, yaitu: 0, 4, 8, dan 12 gulma E. crus-galli/m2
Gambar 14. Perlakuan populasi gulma E. crus-galli/m
. Setiap satuan percobaan
berupa petak berukuran 4 m x 5 m. Jarak antar petak 0.25 cm dan jarak antar
ulangan 0.5 m.
2
pada pertanaman padi
0 E. crus-galli/m2 4 E. crus-galli/m2
12 E. crus-galli/m2
8 E. crus-galli/m2
Keterangan: : tanaman padi : E. crus-galli

77
Bibit padi hibrida berumur 21 hari setelah semai (HSS) ditanam pada
petak percobaan dengan jarak tanam 25 cm x 25 cm. Bibit E. crus-galli berumur
14 HSS ditanam pada petak percobaan sesuai dengan perlakuan k
Pertumbuhan Vegetatif Tanaman Padi
epadatan
populasi. Pemupukan tanaman padi hibrida dilakukan dengan dosis 280 kg
Urea/ha, 100 kg SP-18/ha dan 100 kg KCl/ha. Pupuk Urea diberikan tiga kali
yaitu 100 kg/ha pada saat tanam, 90 kg/ha pada saat 4 MST, dan 90 kg/ha pada
saat 8 MST. Pupuk SP-18 diberikan seluruhnya pada saat tanam. Pupuk KCl
diberikan 80 kg/ha pada saat tanam dan 20 kg/ha pada saat 8 MST. Pemupukan
dilakukan dengan cara sebar. Pemeliharaan tanaman yang dilakukan antara lain
penyiangan gulma selain gulma E. crus-galli pada saat 3 MST dan 6 MST. Panen
padi hibrida dilakukan pada saat tanaman padi berumur 115 HSS atau 96 HST.
Peubah yang diamati antara lain tinggi tanaman, jumlah daun, panjang dan
lebar daun, jumlah anakan, indeks luas daun, bobot kering tajuk dan akar, saat
75% populasi berbunga, jumlah anakan produktif, produksi gabah, persentase
gabah isi dan hampa, bobot gabah 1000 butir, serapan hara tajuk. Peubah yang
diamati pada gulma E. crus-galli antara lain tinggi gulma, jumlah daun, jumlah
anakan, bobot kering tajuk dan akar, panjang malai, kepadatan malai, bobot biji
per malai, bobot biji per 1000 butir. Data hasil pengamatan dianalisis
menggunakan analisis ragam dengan uji lanjut DMRT pada taraf 5%.
Hasil dan Pembahasan
Tinggi tanaman. Tinggi tanaman padi dipengaruhi oleh interaksi antara
aksesi dan populasi gulma E. crus-galli mulai 3 MST hingga 6 MST. Pada saat 6
MST, aksesi gulma E. crus-galli asal Sukabumi menunjukkan tinggi tanaman
yang semakin meningkat dengan semakin meningkatnya populasi. Aksesi gulma
E. crus-galli asal Pangalengan dan Karawang menunjukkan tinggi tanaman yang
semakin menurun dengan semakin meningkatnya populasi gulma E. crus-galli.
Penurunan tinggi tanaman padi pada asesi Karawang lebih tajam dibandingkan
dengan penurunan pada asesi Pangalengan (Gambar 15). Hal ini menunjukkan
bahwa aksesi Karawang lebih kuat daya kompetisinya dibandingkan dengan
aksesi Pangalengan dan aksesi Sukabumi.
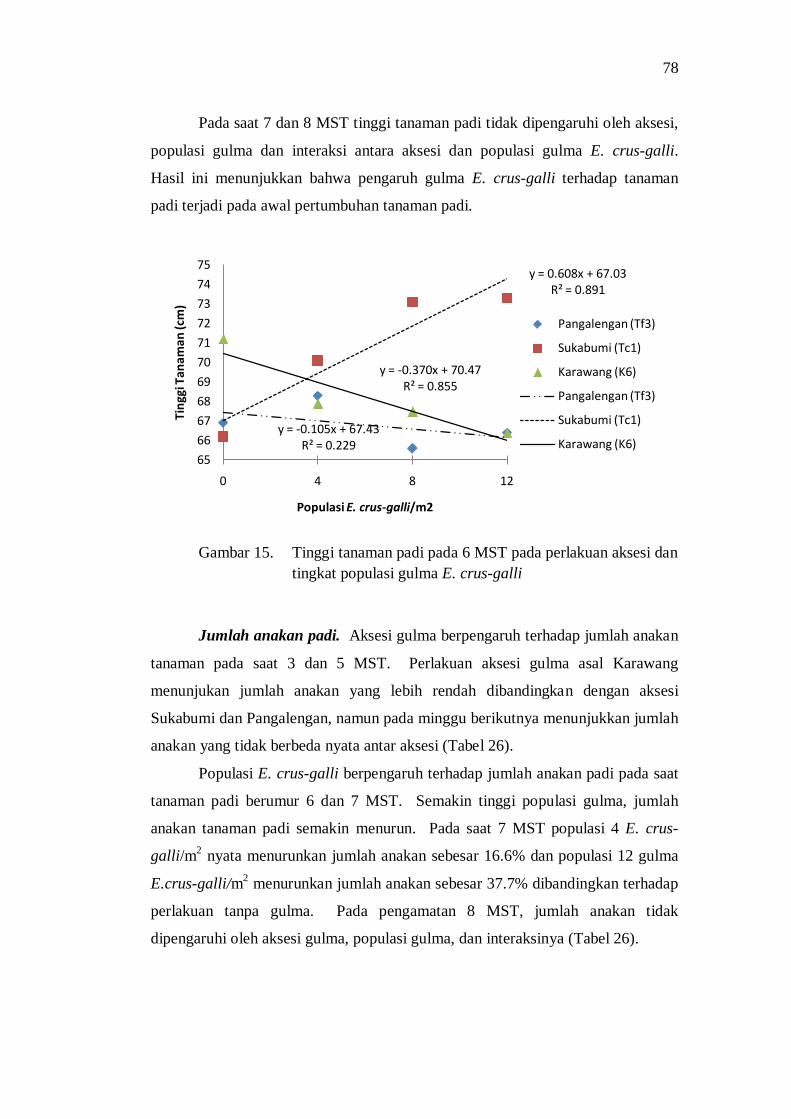
78
Pada saat 7 dan 8 MST tinggi tanaman padi tidak dipengaruhi oleh aksesi,
populasi gulma dan interaksi antara aksesi dan populasi gulma E. crus-galli.
Hasil ini menunjukkan bahwa pengaruh gulma E. crus-galli terhadap tanaman
padi terjadi pada awal pertumbuhan tanaman padi.
Gambar 15. Tinggi tanaman padi pada 6 MST pada perlakuan aksesi dan tingkat populasi gulma E. crus-galli
Jumlah anakan padi. Aksesi gulma berpengaruh terhadap jumlah anakan
tanaman pada saat 3 dan 5 MST. Perlakuan aksesi gulma asal Karawang
menunjukan jumlah anakan yang lebih rendah dibandingkan dengan aksesi
Sukabumi dan Pangalengan, namun pada minggu berikutnya menunjukkan jumlah
anakan yang tidak berbeda nyata antar aksesi (Tabel 26).
Populasi E. crus-galli berpengaruh terhadap jumlah anakan padi pada saat
tanaman padi berumur 6 dan 7 MST. Semakin tinggi populasi gulma, jumlah
anakan tanaman padi semakin menurun. Pada saat 7 MST populasi 4 E. crus-
galli/m2 nyata menurunkan jumlah anakan sebesar 16.6% dan populasi 12 gulma
E.crus-galli/m2
menurunkan jumlah anakan sebesar 37.7% dibandingkan terhadap
perlakuan tanpa gulma. Pada pengamatan 8 MST, jumlah anakan tidak
dipengaruhi oleh aksesi gulma, populasi gulma, dan interaksinya (Tabel 26).
y = -0.105x + 67.43R² = 0.229
y = 0.608x + 67.03R² = 0.891
y = -0.370x + 70.47R² = 0.855
65
66
67
68
69
70
71
72
73
74
75
0 4 8 12
Ting
gi T
anam
an (c
m)
Populasi E. crus-galli/m2
Pangalengan (Tf3)
Sukabumi (Tc1)
Karawang (K6)
Pangalengan (Tf3)
Sukabumi (Tc1)
Karawang (K6)
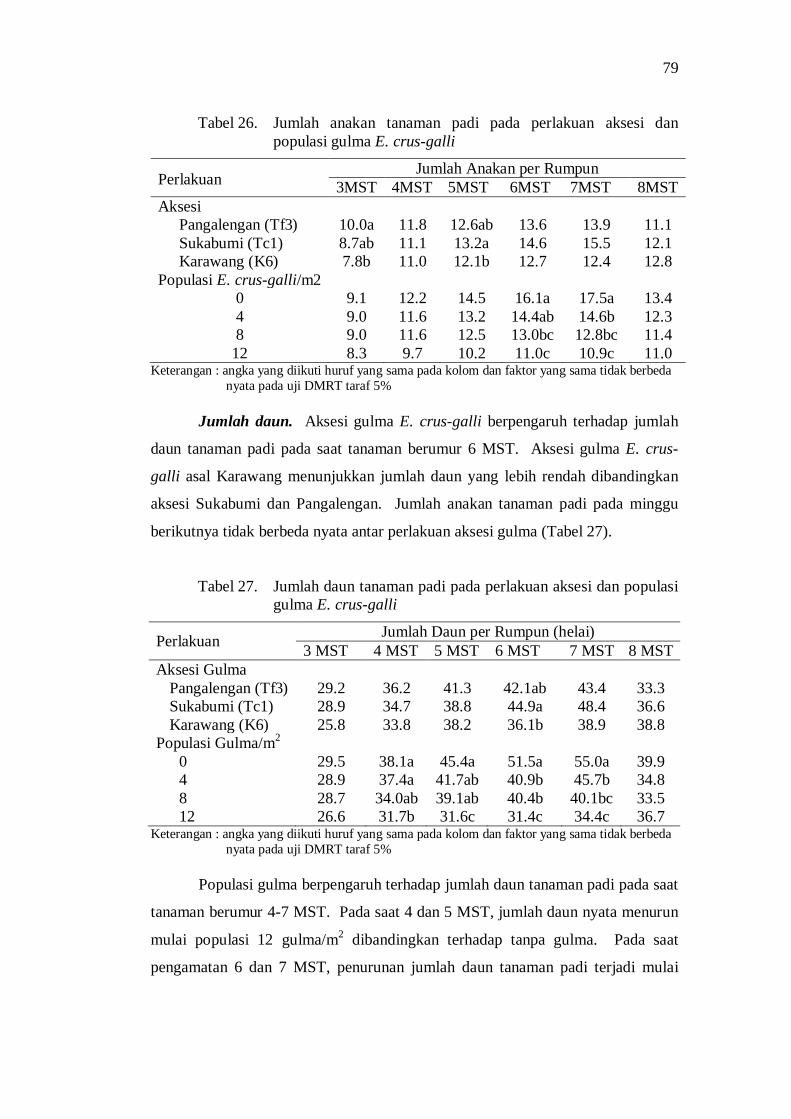
79
Tabel 26. Jumlah anakan tanaman padi pada perlakuan aksesi dan populasi gulma E. crus-galli
Perlakuan Jumlah Anakan per Rumpun
3MST 4MST 5MST 6MST 7MST 8MST Aksesi
Pangalengan (Tf3) 10.0a 11.8 12.6ab 13.6 13.9 11.1 Sukabumi (Tc1) 8.7ab 11.1 13.2a 14.6 15.5 12.1 Karawang (K6) 7.8b 11.0 12.1b 12.7 12.4 12.8
Populasi E. crus-galli/m2 0 9.1 12.2 14.5 16.1a 17.5a 13.4 4 9.0 11.6 13.2 14.4ab 14.6b 12.3 8 9.0 11.6 12.5 13.0bc 12.8bc 11.4
12 8.3 9.7 10.2 11.0c 10.9c 11.0 Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda
nyata pada uji DMRT taraf 5% Jumlah daun. Aksesi gulma E. crus-galli berpengaruh terhadap jumlah
daun tanaman padi pada saat tanaman berumur 6 MST. Aksesi gulma E. crus-
galli asal Karawang menunjukkan jumlah daun yang lebih rendah dibandingkan
aksesi Sukabumi dan Pangalengan. Jumlah anakan tanaman padi pada minggu
berikutnya tidak berbeda nyata antar perlakuan aksesi gulma (Tabel 27).
Tabel 27. Jumlah daun tanaman padi pada perlakuan aksesi dan populasi gulma E. crus-galli
Perlakuan Jumlah Daun per Rumpun (helai) 3 MST 4 MST 5 MST 6 MST 7 MST 8 MST
Aksesi Gulma Pangalengan (Tf3) 29.2 36.2 41.3 42.1ab 43.4 33.3 Sukabumi (Tc1) 28.9 34.7 38.8 44.9a 48.4 36.6 Karawang (K6) 25.8 33.8 38.2 36.1b 38.9 38.8
Populasi Gulma/m 2 0 29.5 38.1a 45.4a 51.5a 55.0a 39.9
4 28.9 37.4a 41.7ab 40.9b 45.7b 34.8 8 28.7 34.0ab 39.1ab 40.4b 40.1bc 33.5 12 26.6 31.7b 31.6c 31.4c 34.4c 36.7
Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda nyata pada uji DMRT taraf 5%
Populasi gulma berpengaruh terhadap jumlah daun tanaman padi pada saat
tanaman berumur 4-7 MST. Pada saat 4 dan 5 MST, jumlah daun nyata menurun
mulai populasi 12 gulma/m2 dibandingkan terhadap tanpa gulma. Pada saat
pengamatan 6 dan 7 MST, penurunan jumlah daun tanaman padi terjadi mulai
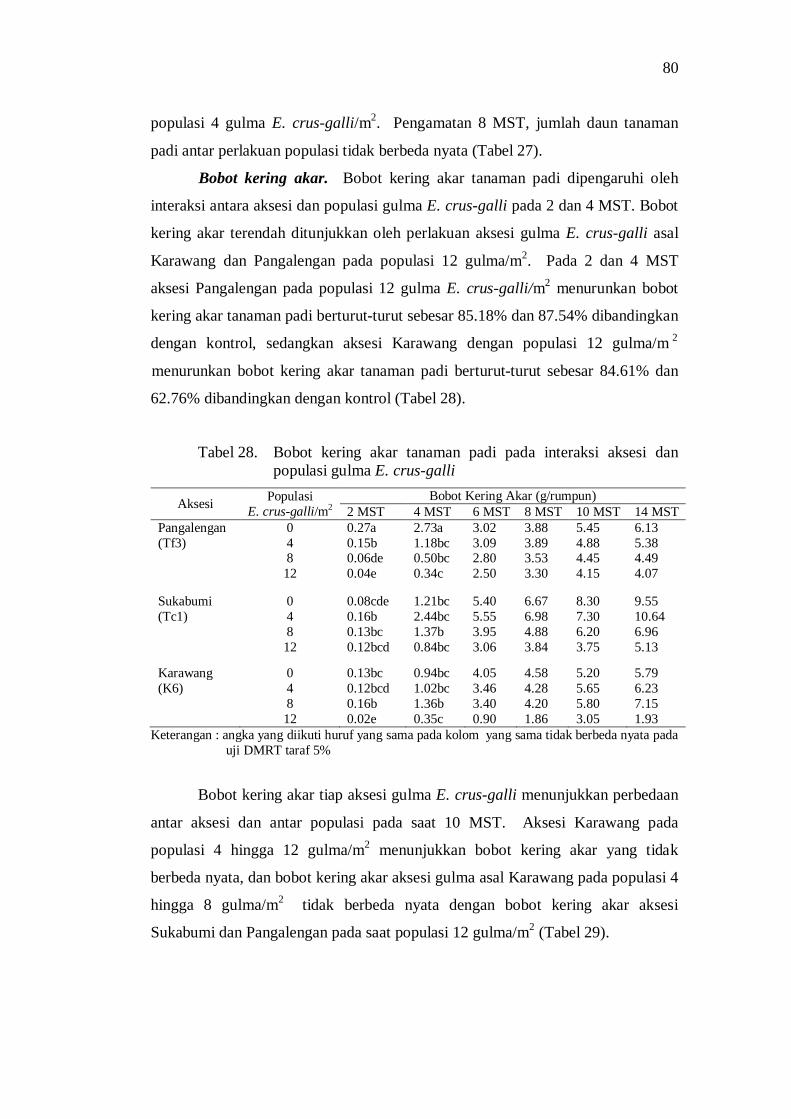
80
populasi 4 gulma E. crus-galli/m2
Bobot kering akar. Bobot kering akar tanaman padi dipengaruhi oleh
interaksi antara aksesi dan populasi gulma E. crus-galli pada 2 dan 4 MST. Bobot
kering akar terendah ditunjukkan oleh perlakuan aksesi gulma E. crus-galli asal
Karawang dan Pangalengan pada populasi 12 gulma/m
. Pengamatan 8 MST, jumlah daun tanaman
padi antar perlakuan populasi tidak berbeda nyata (Tabel 27).
2. Pada 2 dan 4 MST
aksesi Pangalengan pada populasi 12 gulma E. crus-galli/m2 menurunkan bobot
kering akar tanaman padi berturut-turut sebesar 85.18% dan 87.54% dibandingkan
dengan kontrol, sedangkan aksesi Karawang dengan populasi 12 gulma/m 2
Aksesi
menurunkan bobot kering akar tanaman padi berturut-turut sebesar 84.61% dan
62.76% dibandingkan dengan kontrol (Tabel 28).
Tabel 28. Bobot kering akar tanaman padi pada interaksi aksesi dan populasi gulma E. crus-galli
Populasi E. crus-galli/m
Bobot Kering Akar (g/rumpun) 2 2 MST 4 MST 6 MST 8 MST 10 MST 14 MST
Pangalengan 0 0.27a 2.73a 3.02 3.88 5.45 6.13 (Tf3) 4 0.15b 1.18bc 3.09 3.89 4.88 5.38
8 0.06de 0.50bc 2.80 3.53 4.45 4.49
12 0.04e 0.34c 2.50 3.30 4.15 4.07
Sukabumi 0 0.08cde 1.21bc 5.40 6.67 8.30 9.55 (Tc1) 4 0.16b 2.44bc 5.55 6.98 7.30 10.64
8 0.13bc 1.37b 3.95 4.88 6.20 6.96
12 0.12bcd 0.84bc 3.06 3.84 3.75 5.13
Karawang 0 0.13bc 0.94bc 4.05 4.58 5.20 5.79 (K6) 4 0.12bcd 1.02bc 3.46 4.28 5.65 6.23
8 0.16b 1.36b 3.40 4.20 5.80 7.15
12 0.02e 0.35c 0.90 1.86 3.05 1.93 Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada
uji DMRT taraf 5%
Bobot kering akar tiap aksesi gulma E. crus-galli menunjukkan perbedaan
antar aksesi dan antar populasi pada saat 10 MST. Aksesi Karawang pada
populasi 4 hingga 12 gulma/m2 menunjukkan bobot kering akar yang tidak
berbeda nyata, dan bobot kering akar aksesi gulma asal Karawang pada populasi 4
hingga 8 gulma/m2 tidak berbeda nyata dengan bobot kering akar aksesi
Sukabumi dan Pangalengan pada saat populasi 12 gulma/m2 (Tabel 29).
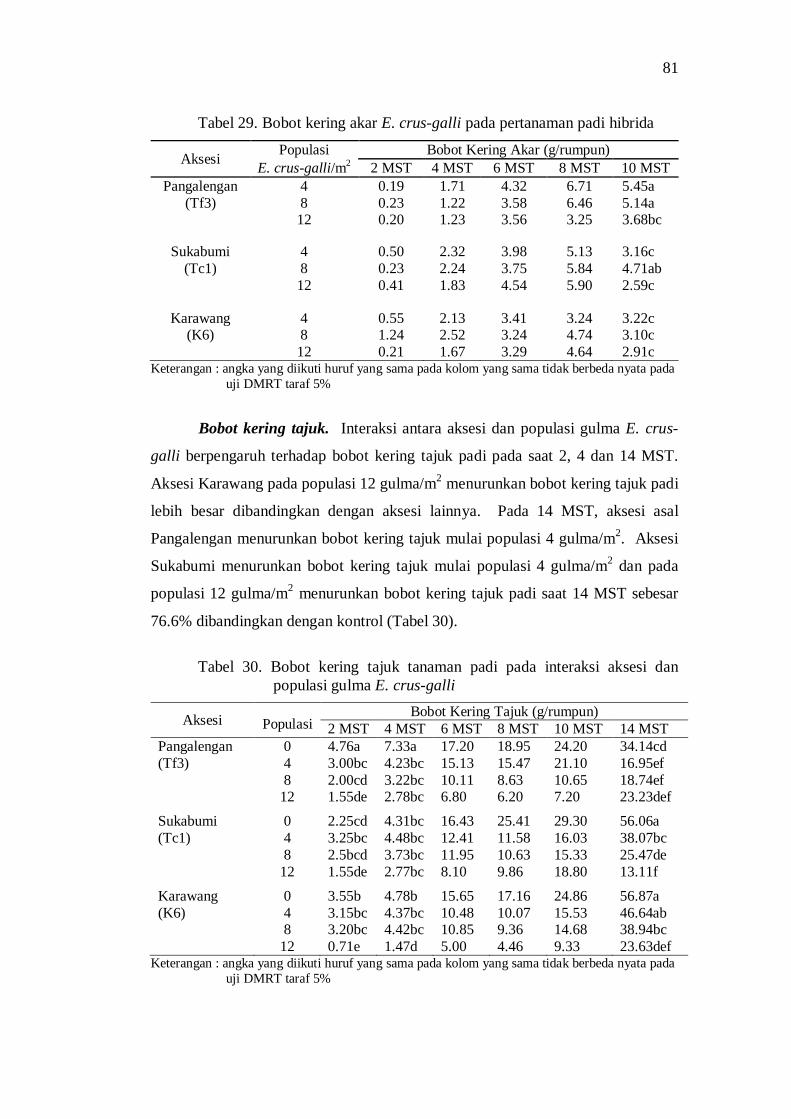
81
Tabel 29. Bobot kering akar E. crus-galli pada pertanaman padi hibrida
Aksesi Populasi Bobot Kering Akar (g/rumpun) E. crus-galli/m 2 MST 2 4 MST 6 MST 8 MST 10 MST
Pangalengan 4 0.19 1.71 4.32 6.71 5.45a (Tf3) 8 0.23 1.22 3.58 6.46 5.14a
12 0.20 1.23 3.56 3.25 3.68bc
Sukabumi 4 0.50 2.32 3.98 5.13 3.16c (Tc1) 8 0.23 2.24 3.75 5.84 4.71ab
12 0.41 1.83 4.54 5.90 2.59c
Karawang 4 0.55 2.13 3.41 3.24 3.22c (K6) 8 1.24 2.52 3.24 4.74 3.10c
12 0.21 1.67 3.29 4.64 2.91c Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada
uji DMRT taraf 5%
Bobot kering tajuk. Interaksi antara aksesi dan populasi gulma E. crus-
galli berpengaruh terhadap bobot kering tajuk padi pada saat 2, 4 dan 14 MST.
Aksesi Karawang pada populasi 12 gulma/m2 menurunkan bobot kering tajuk padi
lebih besar dibandingkan dengan aksesi lainnya. Pada 14 MST, aksesi asal
Pangalengan menurunkan bobot kering tajuk mulai populasi 4 gulma/m2. Aksesi
Sukabumi menurunkan bobot kering tajuk mulai populasi 4 gulma/m2 dan pada
populasi 12 gulma/m2
Tabel 30. Bobot kering tajuk tanaman padi pada interaksi aksesi dan populasi gulma E. crus-galli
menurunkan bobot kering tajuk padi saat 14 MST sebesar
76.6% dibandingkan dengan kontrol (Tabel 30).
Aksesi Populasi Bobot Kering Tajuk (g/rumpun)
2 MST 4 MST 6 MST 8 MST 10 MST 14 MST Pangalengan 0 4.76a 7.33a 17.20 18.95 24.20 34.14cd (Tf3) 4 3.00bc 4.23bc 15.13 15.47 21.10 16.95ef
8 2.00cd 3.22bc 10.11 8.63 10.65 18.74ef
12 1.55de 2.78bc 6.80 6.20 7.20 23.23def
Sukabumi 0 2.25cd 4.31bc 16.43 25.41 29.30 56.06a (Tc1) 4 3.25bc 4.48bc 12.41 11.58 16.03 38.07bc
8 2.5bcd 3.73bc 11.95 10.63 15.33 25.47de
12 1.55de 2.77bc 8.10 9.86 18.80 13.11f
Karawang 0 3.55b 4.78b 15.65 17.16 24.86 56.87a (K6) 4 3.15bc 4.37bc 10.48 10.07 15.53 46.64ab
8 3.20bc 4.42bc 10.85 9.36 14.68 38.94bc
12 0.71e 1.47d 5.00 4.46 9.33 23.63def Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada
uji DMRT taraf 5%
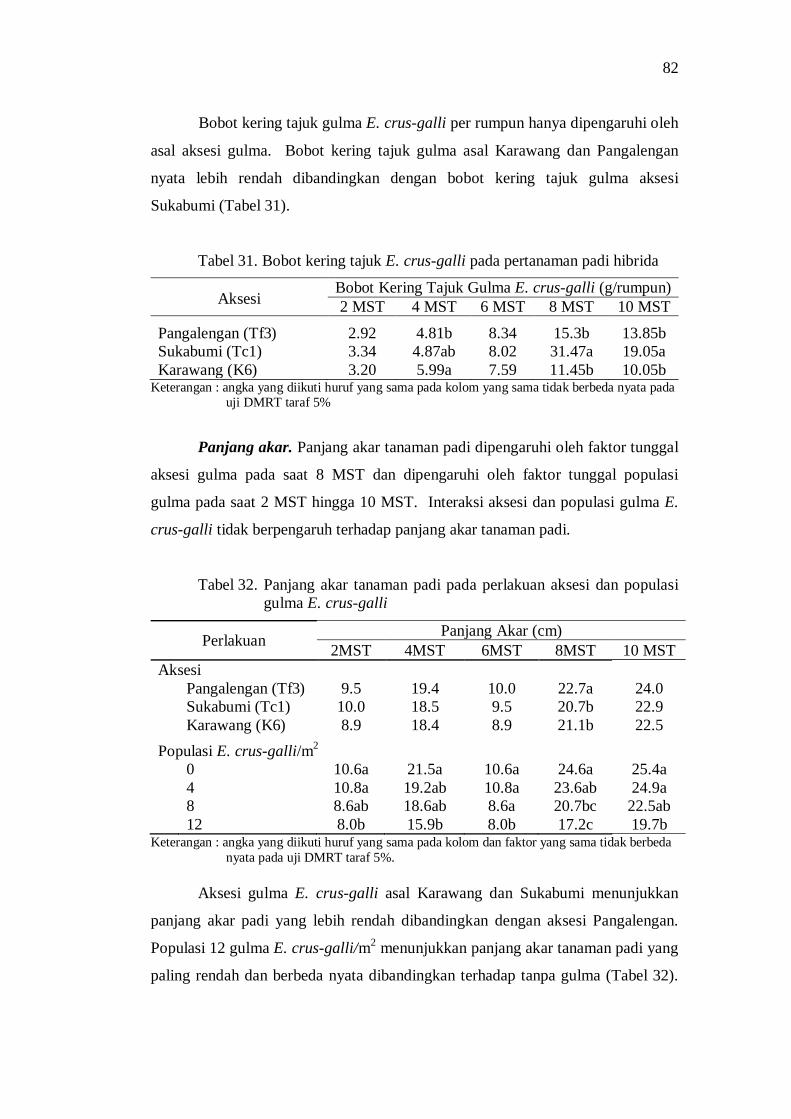
82
Bobot kering tajuk gulma E. crus-galli per rumpun hanya dipengaruhi oleh
asal aksesi gulma. Bobot kering tajuk gulma asal Karawang dan Pangalengan
nyata lebih rendah dibandingkan dengan bobot kering tajuk gulma aksesi
Sukabumi (Tabel 31).
Tabel 31. Bobot kering tajuk E. crus-galli pada pertanaman padi hibrida
Aksesi Bobot Kering Tajuk Gulma E. crus-galli (g/rumpun) 2 MST 4 MST 6 MST 8 MST 10 MST
Pangalengan (Tf3) 2.92 4.81b 8.34 15.3b 13.85b Sukabumi (Tc1) 3.34 4.87ab 8.02 31.47a 19.05a Karawang (K6) 3.20 5.99a 7.59 11.45b 10.05b
Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%
Panjang akar. Panjang akar tanaman padi dipengaruhi oleh faktor tunggal
aksesi gulma pada saat 8 MST dan dipengaruhi oleh faktor tunggal populasi
gulma pada saat 2 MST hingga 10 MST. Interaksi aksesi dan populasi gulma E.
crus-galli tidak berpengaruh terhadap panjang akar tanaman padi.
Tabel 32. Panjang akar tanaman padi pada perlakuan aksesi dan populasi gulma E. crus-galli
Perlakuan Panjang Akar (cm) 2MST 4MST 6MST 8MST 10 MST
Aksesi
Pangalengan (Tf3) 9.5 19.4 10.0 22.7a 24.0
Sukabumi (Tc1) 10.0 18.5 9.5 20.7b 22.9
Karawang (K6) 8.9 18.4 8.9 21.1b 22.5
Populasi E. crus-galli/m
2
0 10.6a 21.5a 10.6a 24.6a 25.4a
4 10.8a 19.2ab 10.8a 23.6ab 24.9a
8 8.6ab 18.6ab 8.6a 20.7bc 22.5ab
12 8.0b 15.9b 8.0b 17.2c 19.7b Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda
nyata pada uji DMRT taraf 5%.
Aksesi gulma E. crus-galli asal Karawang dan Sukabumi menunjukkan
panjang akar padi yang lebih rendah dibandingkan dengan aksesi Pangalengan.
Populasi 12 gulma E. crus-galli/m2 menunjukkan panjang akar tanaman padi yang
paling rendah dan berbeda nyata dibandingkan terhadap tanpa gulma (Tabel 32).

83
Hasil ini menunjukkan bahwa kompetisi gulma E. crus-galli di bawah tanah
(perakaran) terhadap tanaman padi berlangsung dari awal tanam dan pada 10 MST
pengaruh kompetisi masih dapat dilihat dengan penurunan panjang akar tanaman
padi. Penurunan panjang akar ini diduga dapat mempengaruhi tanaman padi
dalam serapan hara di dalam tanah.
Indeks luas daun (ILD). Indeks luas daun tanaman padi hanya
dipengaruhi oleh populasi gulma E. crus-galli. Perlakuan populasi gulma E. crus-
galli mulai 4 gulma/m2 nyata menurunkan ILD tanaman padi hibrida
dibandingkan dengan kontrol. Peningkatan populasi gulma menjadi 8 dan 12
gulma/m2 menunjukkan ILD yang tidak berbeda nyata dengan populasi 4
gulma/m2
Aksesi gulma E. crus-galli asal Karawang menunjukkan ILD gulma yang
lebih besar dibandingkan dengan aksesi asal Sukabumi dan cenderung lebih besar
dibandingkan dengan aksesi asal Pangalengan (Tabel 33). Hasil ini menguatkan
dugaan bahwa aksesi gulma E. crus-galli asal Karawang memiliki derajat
kompetisi yang lebih besar dibandingkan dengan aksesi Pangalengan dan
Sukabumi dalam penangkapan cahaya.
Populasi gulma E. crus-galli berpengaruh terhadap ILD gulma. Semakin
tinggi populasi gulma E. crus-galli, maka ILD gulma semakin tinggi. Pada
populasi 12 gulma/m
(Tabel 33).
2
Tabel 33. Indeks luas daun tanaman padi dan gulma E. crus-galli pada
saat 8 MST dari berbagai perlakuan
, gulma E. crus-galli menunjukkan ILD paling tinggi, yakni
sebesar 2.64 (Tabel 33).
Populasi E. crus-galli/m ILD Padi 2 ILD Gulma Aksesi Gulma
Pangalengan (Tf3) 2.65 1.88ab Sukabumi (Tc1) 2.74 1.52b Karawang (K6) 2.73 2.19a
Populasi E. crus-galli
0 3.20a - 4 2.53b 1.16c 8 2.37b 1.78b 12 2.73b 2.64a
Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda nyata pada uji DMRT taraf 5%
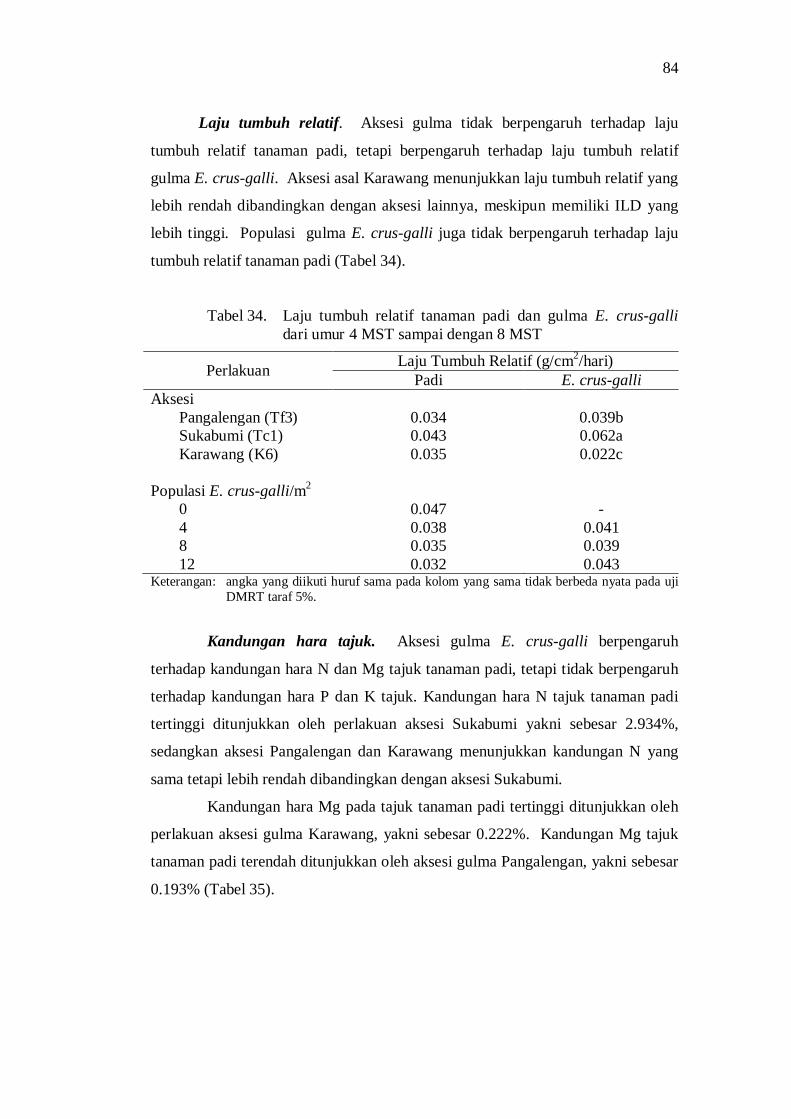
84
Laju tumbuh relatif. Aksesi gulma tidak berpengaruh terhadap laju
tumbuh relatif tanaman padi, tetapi berpengaruh terhadap laju tumbuh relatif
gulma E. crus-galli. Aksesi asal Karawang menunjukkan laju tumbuh relatif yang
lebih rendah dibandingkan dengan aksesi lainnya, meskipun memiliki ILD yang
lebih tinggi. Populasi gulma E. crus-galli juga tidak berpengaruh terhadap laju
tumbuh relatif tanaman padi (Tabel 34).
Tabel 34. Laju tumbuh relatif tanaman padi dan gulma E. crus-galli dari umur 4 MST sampai dengan 8 MST
Perlakuan Laju Tumbuh Relatif (g/cm2/hari) Padi E. crus-galli
Aksesi
Pangalengan (Tf3) 0.034 0.039b
Sukabumi (Tc1) 0.043 0.062a
Karawang (K6) 0.035 0.022c
Populasi E. crus-galli/m 2
0 0.047 -
4 0.038 0.041
8 0.035 0.039
12 0.032 0.043 Keterangan: angka yang diikuti huruf sama pada kolom yang sama tidak berbeda nyata pada uji
DMRT taraf 5%.
Kandungan hara tajuk. Aksesi gulma E. crus-galli berpengaruh
terhadap kandungan hara N dan Mg tajuk tanaman padi, tetapi tidak berpengaruh
terhadap kandungan hara P dan K tajuk. Kandungan hara N tajuk tanaman padi
tertinggi ditunjukkan oleh perlakuan aksesi Sukabumi yakni sebesar 2.934%,
sedangkan aksesi Pangalengan dan Karawang menunjukkan kandungan N yang
sama tetapi lebih rendah dibandingkan dengan aksesi Sukabumi.
Kandungan hara Mg pada tajuk tanaman padi tertinggi ditunjukkan oleh
perlakuan aksesi gulma Karawang, yakni sebesar 0.222%. Kandungan Mg tajuk
tanaman padi terendah ditunjukkan oleh aksesi gulma Pangalengan, yakni sebesar
0.193% (Tabel 35).
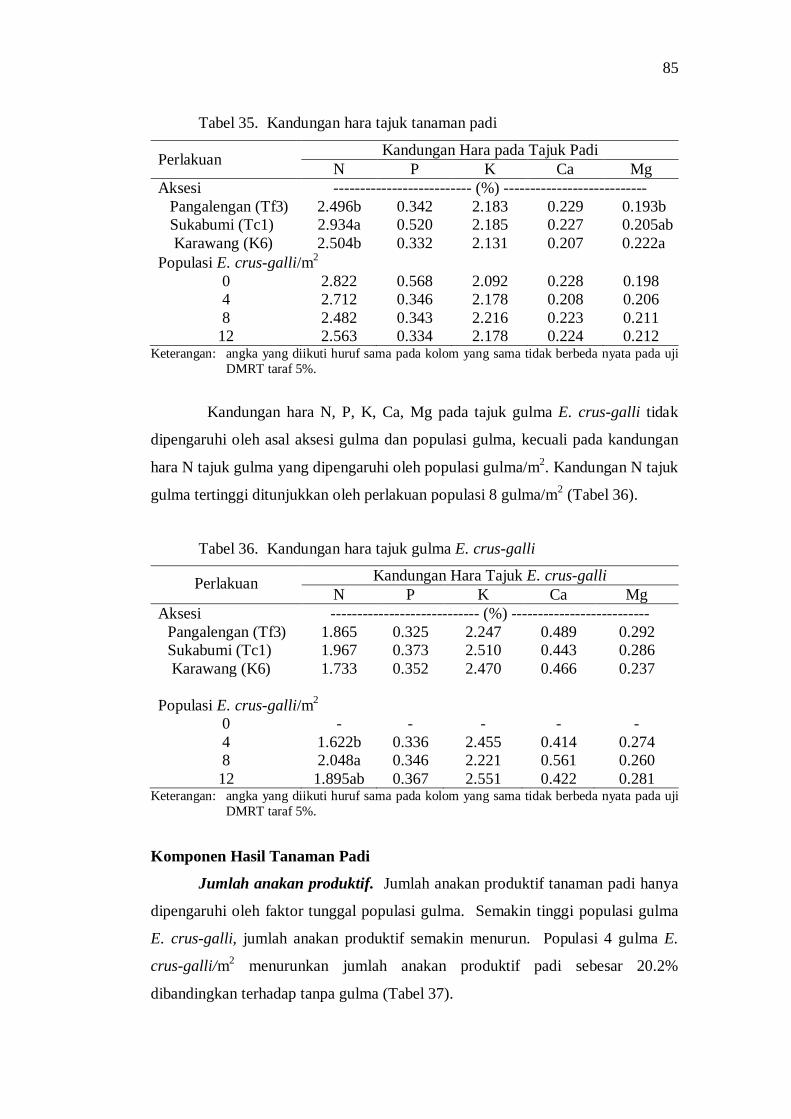
85
Tabel 35. Kandungan hara tajuk tanaman padi
Perlakuan Kandungan Hara pada Tajuk Padi N P K Ca Mg
Aksesi -------------------------- (%) --------------------------- Pangalengan (Tf3) 2.496b 0.342 2.183 0.229 0.193b Sukabumi (Tc1) 2.934a 0.520 2.185 0.227 0.205ab Karawang (K6) 2.504b 0.332 2.131 0.207 0.222a
Populasi E. crus-galli/m2
0 2.822 0.568 2.092 0.228 0.198 4 2.712 0.346 2.178 0.208 0.206 8 2.482 0.343 2.216 0.223 0.211
12 2.563 0.334 2.178 0.224 0.212 Keterangan: angka yang diikuti huruf sama pada kolom yang sama tidak berbeda nyata pada uji
DMRT taraf 5%. Kandungan hara N, P, K, Ca, Mg pada tajuk gulma E. crus-galli tidak
dipengaruhi oleh asal aksesi gulma dan populasi gulma, kecuali pada kandungan
hara N tajuk gulma yang dipengaruhi oleh populasi gulma/m2. Kandungan N tajuk
gulma tertinggi ditunjukkan oleh perlakuan populasi 8 gulma/m2
Perlakuan
(Tabel 36).
Tabel 36. Kandungan hara tajuk gulma E. crus-galli
Kandungan Hara Tajuk E. crus-galli N P K Ca Mg
Aksesi ---------------------------- (%) -------------------------- Pangalengan (Tf3) 1.865 0.325 2.247 0.489 0.292 Sukabumi (Tc1) 1.967 0.373 2.510 0.443 0.286 Karawang (K6) 1.733 0.352 2.470 0.466 0.237
Populasi E. crus-galli/m2
0 - - - - - 4 1.622b 0.336 2.455 0.414 0.274 8 2.048a 0.346 2.221 0.561 0.260 12 1.895ab 0.367 2.551 0.422 0.281
Keterangan: angka yang diikuti huruf sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Komponen Hasil Tanaman Padi
Jumlah anakan produktif. Jumlah anakan produktif tanaman padi hanya
dipengaruhi oleh faktor tunggal populasi gulma. Semakin tinggi populasi gulma
E. crus-galli, jumlah anakan produktif semakin menurun. Populasi 4 gulma E.
crus-galli/m2 menurunkan jumlah anakan produktif padi sebesar 20.2%
dibandingkan terhadap tanpa gulma (Tabel 37).
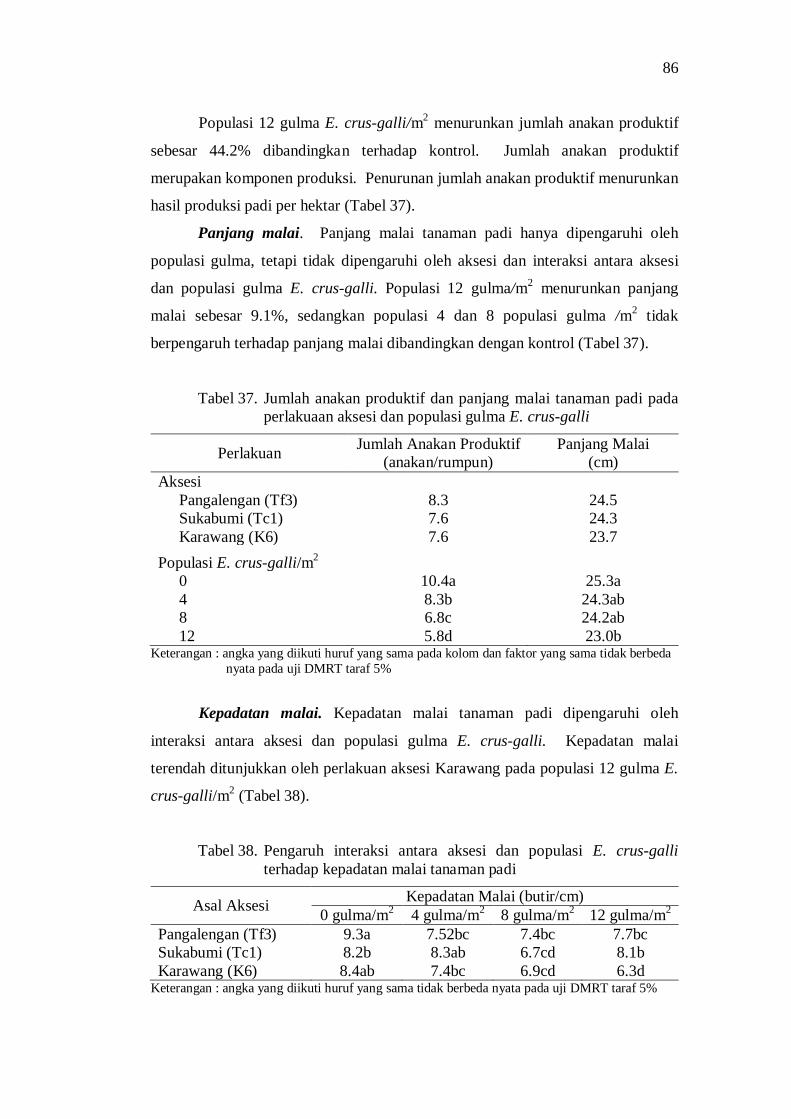
86
Populasi 12 gulma E. crus-galli/m2 menurunkan jumlah anakan produktif
sebesar 44.2% dibandingkan terhadap kontrol. Jumlah anakan produktif
merupakan komponen produksi. Penurunan jumlah anakan produktif menurunkan
hasil produksi padi per hektar (Tabel 37).
Panjang malai. Panjang malai tanaman padi hanya dipengaruhi oleh
populasi gulma, tetapi tidak dipengaruhi oleh aksesi dan interaksi antara aksesi
dan populasi gulma E. crus-galli. Populasi 12 gulma/m2 menurunkan panjang
malai sebesar 9.1%, sedangkan populasi 4 dan 8 populasi gulma /m2
Perlakuan
tidak
berpengaruh terhadap panjang malai dibandingkan dengan kontrol (Tabel 37).
Tabel 37. Jumlah anakan produktif dan panjang malai tanaman padi pada perlakuaan aksesi dan populasi gulma E. crus-galli
Jumlah Anakan Produktif (anakan/rumpun)
Panjang Malai (cm)
Aksesi Pangalengan (Tf3) 8.3 24.5
Sukabumi (Tc1) 7.6 24.3 Karawang (K6) 7.6 23.7 Populasi E. crus-galli/m
2 0 10.4a 25.3a 4 8.3b 24.3ab 8 6.8c 24.2ab 12 5.8d 23.0b
Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda nyata pada uji DMRT taraf 5%
Kepadatan malai. Kepadatan malai tanaman padi dipengaruhi oleh
interaksi antara aksesi dan populasi gulma E. crus-galli. Kepadatan malai
terendah ditunjukkan oleh perlakuan aksesi Karawang pada populasi 12 gulma E.
crus-galli/m2
Tabel 38. Pengaruh interaksi antara aksesi dan populasi E. crus-galli terhadap kepadatan malai tanaman padi
(Tabel 38).
Asal Aksesi Kepadatan Malai (butir/cm) 0 gulma/m 4 gulma/m2 8 gulma/m2 12 gulma/m2 2
Pangalengan (Tf3) 9.3a 7.52bc 7.4bc 7.7bc Sukabumi (Tc1) 8.2b 8.3ab 6.7cd 8.1b Karawang (K6) 8.4ab 7.4bc 6.9cd 6.3d
Keterangan : angka yang diikuti huruf yang sama tidak berbeda nyata pada uji DMRT taraf 5%

87
Produksi tanaman padi ditentukan oleh komponen panjang malai dan
kepadatan malai. Semakin panjang malai dan semakin tinggi kepadatan malainya,
maka hasil produksi tanaman padi akan semakin tinggi. Hasil ini menunjukkan
bahwa aksesi Karawang dengan populasi 12 gulma/m2 menyebabkan penurunan
hasil produksi terbesar.
Jumlah gabah dan persentase kehampaan per malai. Aksesi gulma E.
crus-galli berpengaruh terhadap jumlah gabah hampa per malai, tetapi tidak
berpengaruh terhadap jumlah gabah isi, jumlah gabah total, persentase
kehampaan. Namun terlihat bahwa aksesi Karawang menunjukkan jumlah gabah
total dan jumlah gabah isi yang cenderung lebih rendah dibandingkan dengan
perlakuan aksesi Pangalengan dan Sukabumi (Tabel 39).
Populasi gulma E. crus-galli berpengaruh terhadap jumlah gabah isi dan
jumlah gabah total/malai dan persentase kehampaan. Populasi 4 gulma/m2
menurunkan jumlah gabah total dan gabah isi berturut-turut sebesar 14.1% dan
18.6%. Jumlah gabah total dan gabah isi semakin menurun dengan semakin
meningkatnya populasi gulma. Perlakuan populasi 12 gulma E. crus-galli/m2
Tabel 39. Pengaruh aksesi dan populasi E. crus-galli terhadap jumlah gabah per malai dan persentase kehampaan
menurunkan jumlah gabah total sebesar 22.7% dan menurunkan jumlah gabah isi
sebesar 36.1% serta meningkatkan persentase kehampaan sebesar 54%
dibandingkan terhadap tanpa gulma (Tabel 39).
Perlakuan Jumlah Gabah per Malai (butir) Persen Hampa (%) Total Isi Hampa
Aksesi Pangalengan (Tf3) 196.6 130.9 65.7a 34.1
Sukabumi (Tc1) 190.4 136.9 53.5ab 28.3 Karawang (K6) 172.9 127.2 45.7b 26.7 Populasi E. crus-galli/m
2 0 219.1a 166.4a 52.7 23.9b 4 188.1b 135.4b 52.7 28.1b 8 170.0c 118.6c 51.4 30.1ab 12 169.3c 106.3c 63.0 36.8a
Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda nyata pada uji DMRT taraf 5%
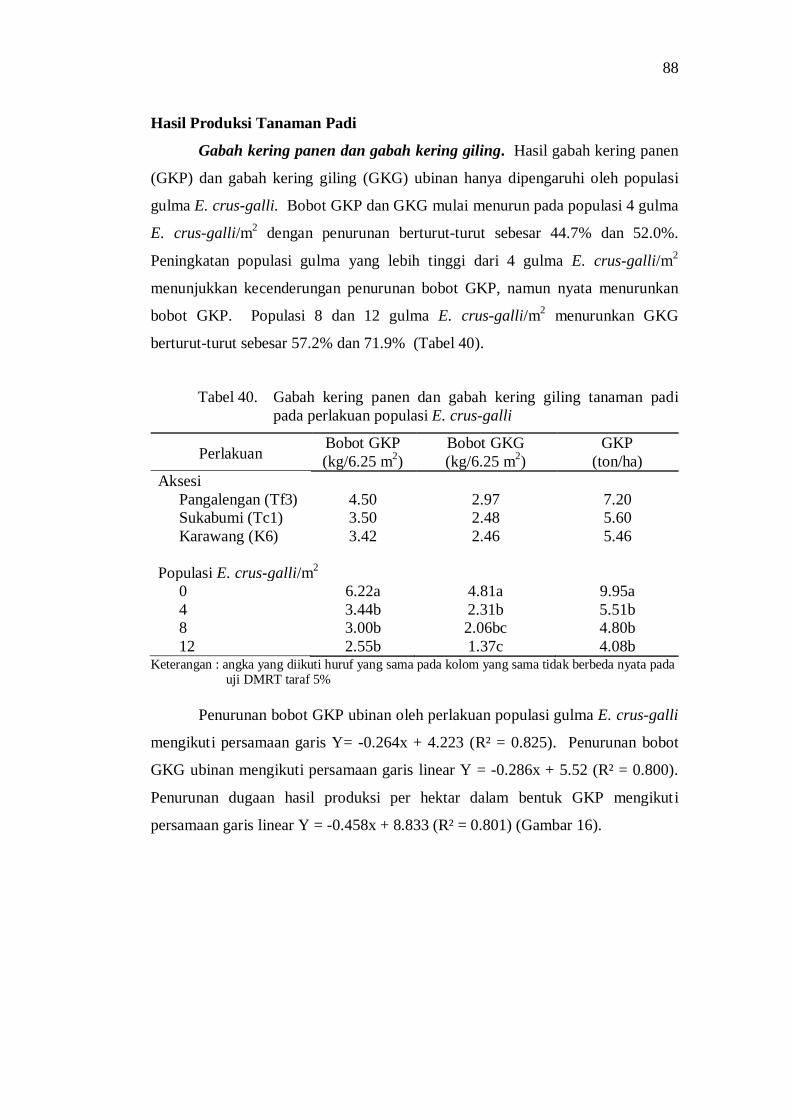
88
Hasil Produksi Tanaman Padi
Gabah kering panen dan gabah kering giling. Hasil gabah kering panen
(GKP) dan gabah kering giling (GKG) ubinan hanya dipengaruhi oleh populasi
gulma E. crus-galli. Bobot GKP dan GKG mulai menurun pada populasi 4 gulma
E. crus-galli/m2 dengan penurunan berturut-turut sebesar 44.7% dan 52.0%.
Peningkatan populasi gulma yang lebih tinggi dari 4 gulma E. crus-galli/m2
menunjukkan kecenderungan penurunan bobot GKP, namun nyata menurunkan
bobot GKP. Populasi 8 dan 12 gulma E. crus-galli/m2
Tabel 40. Gabah kering panen dan gabah kering giling tanaman padi
pada perlakuan populasi E. crus-galli
menurunkan GKG
berturut-turut sebesar 57.2% dan 71.9% (Tabel 40).
Perlakuan Bobot GKP (kg/6.25 m2
Bobot GKG (kg/6.25 m) 2
GKP (ton/ha) )
Aksesi
Pangalengan (Tf3) 4.50 2.97 7.20 Sukabumi (Tc1) 3.50 2.48 5.60 Karawang (K6) 3.42 2.46 5.46
Populasi E. crus-galli/m
2 0 6.22a 4.81a 9.95a 4 3.44b 2.31b 5.51b 8 3.00b 2.06bc 4.80b 12 2.55b 1.37c 4.08b
Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%
Penurunan bobot GKP ubinan oleh perlakuan populasi gulma E. crus-galli
mengikuti persamaan garis Y= -0.264x + 4.223 (R² = 0.825). Penurunan bobot
GKG ubinan mengikuti persamaan garis linear Y = -0.286x + 5.52 (R² = 0.800).
Penurunan dugaan hasil produksi per hektar dalam bentuk GKP mengikuti
persamaan garis linear Y = -0.458x + 8.833 (R² = 0.801) (Gambar 16).
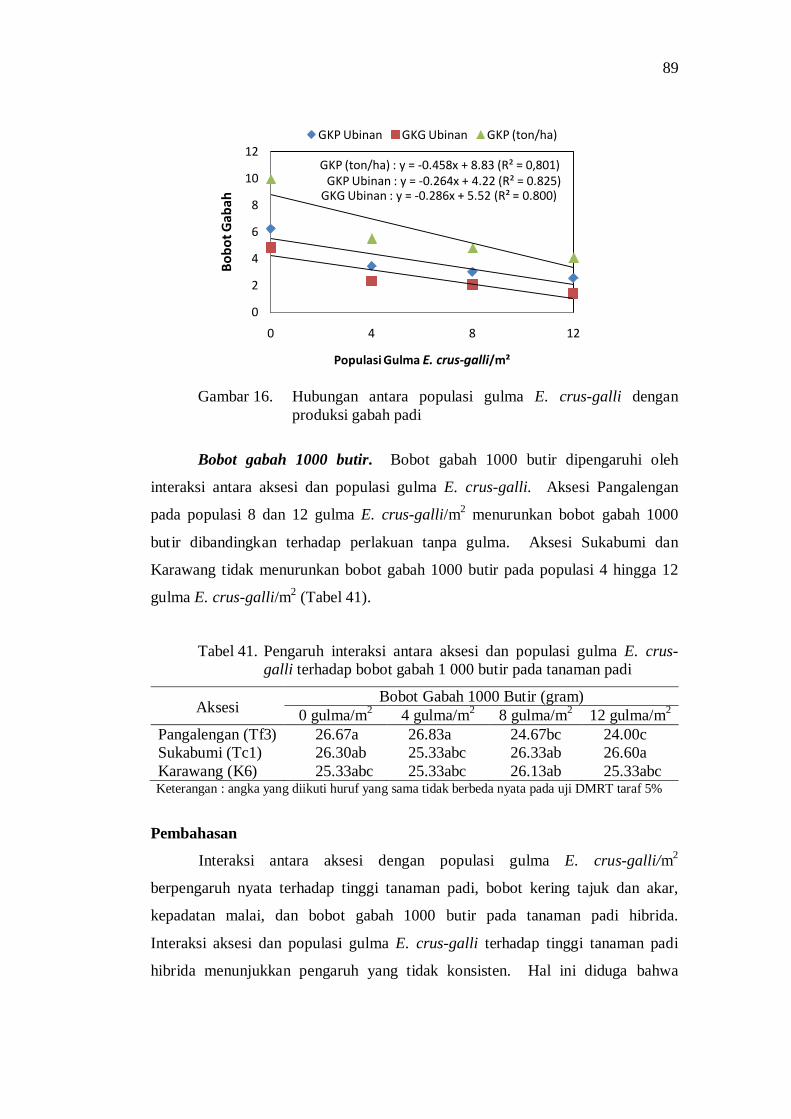
89
Gambar 16. Hubungan antara populasi gulma E. crus-galli dengan
produksi gabah padi
Bobot gabah 1000 butir. Bobot gabah 1000 butir dipengaruhi oleh
interaksi antara aksesi dan populasi gulma E. crus-galli. Aksesi Pangalengan
pada populasi 8 dan 12 gulma E. crus-galli/m2 menurunkan bobot gabah 1000
butir dibandingkan terhadap perlakuan tanpa gulma. Aksesi Sukabumi dan
Karawang tidak menurunkan bobot gabah 1000 butir pada populasi 4 hingga 12
gulma E. crus-galli/m2
Tabel 41. Pengaruh interaksi antara aksesi dan populasi gulma E. crus-galli terhadap bobot gabah 1 000 butir pada tanaman padi
(Tabel 41).
Aksesi Bobot Gabah 1000 Butir (gram)
0 gulma/m 4 gulma/m2 8 gulma/m2 12 gulma/m2 2 Pangalengan (Tf3) 26.67a 26.83a 24.67bc 24.00c Sukabumi (Tc1) 26.30ab 25.33abc 26.33ab 26.60a Karawang (K6) 25.33abc 25.33abc 26.13ab 25.33abc
Keterangan : angka yang diikuti huruf yang sama tidak berbeda nyata pada uji DMRT taraf 5%
Pembahasan
Interaksi antara aksesi dengan populasi gulma E. crus-galli/m2
berpengaruh nyata terhadap tinggi tanaman padi, bobot kering tajuk dan akar,
kepadatan malai, dan bobot gabah 1000 butir pada tanaman padi hibrida.
Interaksi aksesi dan populasi gulma E. crus-galli terhadap tinggi tanaman padi
hibrida menunjukkan pengaruh yang tidak konsisten. Hal ini diduga bahwa
GKG Ubinan : y = -0.286x + 5.52 (R² = 0.800)GKP Ubinan : y = -0.264x + 4.22 (R² = 0.825)
GKP (ton/ha) : y = -0.458x + 8.83 (R² = 0,801)
0
2
4
6
8
10
12
0 4 8 12
Bobo
t Gab
ah
Populasi Gulma E. crus-galli/m²
GKP Ubinan GKG Ubinan GKP (ton/ha)

90
tanaman padi mengalami etiolasi akibat pengaruh naungan gulma E. crus-galli.
Menurut Galinato et al. (1999) gulma E. crus-galli merupakan tumbuhan
golongan rumput (graminae) yang memiliki karakteristik pertumbuhan yang cepat
pada masa vegetatif. Menurut De Datta (1981) tanaman yang berasal dari jenis
dan habitat yang sama dapat terjadi kompetisi yang lebih besar karena memiliki
kebutuhan sumberdaya yang sama.
Aksesi gulma E. crus-galli berpengaruh terhadap pertumbuhan tanaman
padi hibrida yaitu pada peubah jumlah anakan, jumlah daun, panjang akar, ILD,
serapan hara, dan jumlah gabah hampa. Hasil ini sejalan dengan hasil penelitian
pendahuluan Guntoro et al. (2009) bahwa perbedaan aksesi gulma E. crus-galli
menyebabkan perbedaan pertumbuhan dan produksi tanaman padi khususnya
pada peubah tinggi, jumlah anakan dan kepadatan malai. Perbedaan pengaruh tiap
aksesi gulma E. crus-galli diduga karena adanya derajat kompetisi aksesi gulma
yang berbeda untuk memperoleh sarana tumbuh dan zat alelopati yang
dikeluarkan terhadap tanaman padi. Yamamoto et al. (1999) dan Xuan et al.
(2006) menyatakan bahwa eksudat akar E. crus-galli menyebabkan penurunan
perkecambahan dan pertumbuhan tanaman padi. Aksesi Karawang memiliki daya
kompetisi yang lebih kuat dibandingkan dengan aksesi Sukabumi dan
Pangalengan terhadap pertumbuhan tanaman padi hibrida, terutama dalam hal
menekan jumlah anakan pada pengamtan 3 dan 5 MST, jumlah daun pada 6 MST,
dan panjang akar pada 8 MST.
Aksesi gulma E. crus-galli asal Karawang memiliki derajat kompetisi
yang kuat terhadap pertumbuhan tanaman padi yang ditunjukkan dengan nilai ILD
yang lebih besar dibandingkan dengan aksesi Pangalengan dan Sukabumi,
menyebabkan penurunan tinggi tanaman padi yang lebih besar dibandingkan
dengan aksesi Pangalengan dan Sukabumi pada saat 3-5 MST, menyebabkan
penurunan panjang akar, bobot akar dan tajuk, jumlah anakan produktif serta
kepadatan malai yang lebih besar dibandingkan dengan aksesi lainnya. Penurunan
bobot akar dan tajuk diduga mempengaruhi proses pertumbuhan dan
perkembangan pada fase generatif, sebagai akibatnya jumlah anakan produktif
menurun. Penurunan jumlah anakan produktif diikuti dengan penurunan

91
kepadatan malai menyebabkan penurunan hasil produksi tanaman padi yang lebih
besar dibandingkan dengan aksesi Pangalengan dan Sukabumi.
Populasi gulma E. crus-galli nyata berpengaruh terhadap pertumbuhan dan
produksi tanaman padi hibrida, terutama pada peubah jumlah anakan, jumlah
daun, panjang akar, ILD, jumlah anakan produktif, panjang malai, jumlah gabah
per malai, gabah isi, persen biji hampa per malai, gabah kering panen dan gabah
kering giling. Menurut Ampong-Nyarko dan Datta (1991) salah satu faktor yang
menentukan tingkat kompetisi antara padi dan gulma adalah kepadatan gulma
pada pertanaman padi. Purba (2007) menambahkan bahwa semakin tinggi
kerapatan E. crus-galli per m2, maka penurunan hasil tanaman padi semakin
besar. Perlakuan populasi 4, 8 dan 12 gulma E. crus-galli per m2 nyata
menurunkan gabah kering panen dibandingkan dengan kontrol. Pengaruh
perlakuan populasi terhadap kehilangan gabah kering panen berkisar 44-59%.
Penurunan bobot GKP per hektar mengikuti persamaan garis linear Y = -
0.458x + 8.833 (R² = 0.801). Berdasarkan persamaan tersebut, penurunan
produksi sebesar 10% terjadi pada saat populasi gulma sebanyak 1.9 gulma/m2.
Untuk menyelamatkan kehilangan hasil sebesar 10%, maka gulma harus
dikendalikan ketika populasi gulma E. crus-galli di lapangan sudah mencapai 1.9
individu/m2
.
Kesimpulan dan Saran
Perbedaan aksesi gulma E. crus-galli menyebabkan perbedaan
pertumbuhan dan produksi tanaman padi. Aksesi mempengaruhi jumlah anakan,
jumlah daun, dan panjang akar tanaman padi. Memperkuat hasil percobaan rumah
kaca, aksesi asal Karawang (K6, ketinggian tempat 37 m dpl) memiliki derajat
kompetisi yang lebih kuat dibandingkan dengan aksesi Sukabumi dan
Pangalengan terhadap pertumbuhan tanaman padi hibrida yang ditunjukkan
dengan penurunan jumlah anakan tanaman padi pada 3 dan 5 MST, jumlah daun
pada 6 MST, dan panjang akar pada 8 MST, bobot kering akar dan tajuk, jumlah
anakan produktif, dan kepadatan malai.
Kepadatan populasi gulma E. crus-galli mempengaruhi pertumbuhan dan
produksi tanaman padi hibrida. Semakin tinggi populasi gulma E. crus-galli,

92
pengaruh kompetisi terhadap tanaman padi semakin besar. Interaksi antara aksesi
dan gulma E. crus-galli nyata mempengaruhi peubah tinggi, bobot kering akar dan
tajuk, kepadatan malai dan bobot 1 000 butir tanaman padi hibrida.
Gulma E. crus-galli harus dikendalikan pada saat populasi gulma
mencapai 1.9 gulma/m2 untuk menyelamatkan kehilangan hasil sebesar 10%.
Penggunaan benih padi yang bebas dari biji gulma E.crus-galli sangat dianjurkan
untuk mencegah penyebaran aksesi gulma E. crus-galli.

PENDUGAAN DERAJAT KOMPETISI GULMA Echinochloa crus-galli (L.) Beauv. MELALUI
METODE REPLACEMENT SERIES
ABSTRAK
Salah satu cara gulma E. crus-galli mempengaruhi pertumbuhan dan produksi
tanaman padi adalah melalui kompetisi. Tujuan penelitian adalah untuk
menganalisis derajat kompetisi gulma Echinochloa crus-galli terhadap tanaman
padi. Percobaan dilakukan dengan metode replacement series yang disusun
dalam rancangan acak kelompok dengan tiga ulangan. Populasi tanaman yang
dicobakan yaitu 1 gulma E. crus-galli/pot (G1), 2 gulma/pot (G2), 3 gulma/pot
(G3), 4 gulma/pot (G4), 1 padi/pot (P1), 2 padi/pot (P2), 3 padi/pot (P3), 4
padi/pot (P4), 3 padi + 1 gulma/pot (P3+G1), 2 padi + 2 gulma/pot (P2+G2), dan
1 padi + 3 gulma/pot (P1+G3). Hasil penelitian menunjukkan bahwa kompetisi
intraspesifik terjadi baik pada tanaman padi maupun pada gulma E.crus-galli
ketika populasi meningkat. Kompetisi intraspesifik pada gulma E. crus-galli lebih
besar dibandingkan dengan kompetisi intraspesifik pada tanaman padi.
Berdasarkan nilai koefisien pendesakan, gulma E. crus-galli memiliki derajat
kompetisi yang lebih besar terhadap tanaman padi ketika populasi gulma lebih
tinggi daripada populasi tanaman padi. Pada populasi yang sama antara padi dan
gulma ataupun populasi padi lebih tinggi dari gulma E. crus-galli, tanaman padi
lebih kuat berkompetisi. Berdasarkan nilai agresivitas, gulma E. crus-galli lebih
kuat berkompetisi dibandingkan tanaman padi ketika populasi padi dan gulma
sama ataupun populasi gulma lebih tinggi daripada populasi tanaman padi.
Kata kunci : Echinochloa crus-galli, derajat kompetisi, metode replacement series,
koefisien pendesakan, agresivitas.

94
ESTIMATION OF COMPETITION DEGREES OF Echinocloa crus-galli (L.) Beauv. WEED THROUGH REPLACEMENT
SERIES METHOD
ABSTRACT
One way of Echinochloa crus-galli weed affect the growth and production of rice plants is through competition. The research objective was to analyze the degree of competition of E. crus-galli weed on rice plants. The experiments were conducted by the replacement series method that were arranged in randomized block design with three replications. Populations of plants tested were 1 E. crus-galli weed/pot (G1), 2 weed/pot (G2), 3 weed/pot (G3), 4 weed/pot (G4), 1 rice plant/pot (P1), 2 rice plants/pot (P2), 3 rice plants/pot (P3), 4 rice plants/pot (P4), 3 rice plants + 1 weed/pot (P3 + G1), 2 rice plants + 2 weeds/pot (P2 + G2), and 1 rice plant + 3 weeds/pot (P1 + G3). The results showed that intraspecific competition occurs both in rice and in E.crus-galli weeds when the population increased. Intraspecific competition in E. crus-galli weeds was greater than intraspecific competition in the rice plant. Based on crawding coefficient, E. crus-galli weeds had a greater degree of competition to rice plants when the population weed was higher than the rice plants population. At the same populations between rice and weeds or rice population was higher than the E. crus-galli weeds, rice plants had stronger compete to E. crus-galli. Based on the aggressiveness, E. crus-galli weed competed more strongly than rice plant when the rice population and weeds was equal or the population
weed was higher than the rice population.
Keywords : Echinochloa crus-galli, competition degree, replacement series, crawding coefficient, aggresiveness.

95
Pendahuluan
Gulma merupakan salah satu diantara pembatas biologi yang penting pada
produksi padi di Indonesia. Kehilangan hasil akibat gulma bervariasi dari 28 –
54% pada tanaman padi pindahtanam (transplanting) dan 28-89% pada tanaman
padi tabur benih langsung (direct seeded) (Becker et al. 2003; Johnson et al.
2004). Gulma Echinochloa crus-galli merupakan gulma dominan pada padi
sawah yang dapat menurunkan hasil produksi tanaman padi hingga 72% (Islam et
al. 2003), memiliki daya adaptasi yang luas (Galinato et al. 1999), termasuk
tumbuhan C4 yang efisien dalam fotosintesis dan memiliki tingkat kompetisi yang
tinggi (Baki dan Azmi 2003).
Salah satu cara gulma E. crus-galli untuk mempengaruhi pertumbuhan dan
hasil produksi tanaman padi adalah melalui kompetisi. Kompetisi didefinisikan
sebagai hubungan negatif antara dua individu baik sama jenis ataupun berbeda
jenis yang diakibatkan oleh pemakaian secara bersama sumberdaya yang dalam
kondisi terbatas (Sastroutomo, 1990). Kompetisi terjadi apabila tanaman padi dan
gulma E. crus-galli hidup bersama pada tempat yang memiliki sumberdaya
terbatas.
Ada tiga bentuk kompetisi yang terjadi di antara spesies, yaitu kompetisi
yang mengakibatkan hasil sesungguhnya dari masing-masing spesies dalam
pertanaman campuran lebih rendah dari hasil yang diharapkan (mutual inhibition),
kompetisi yang mengakibatkan hasil dari masing-masing spesies dalam
pertanaman campuran lebih besar dari hasil yang diharapkan (mutual
cooperation), dan kompetisi yang mengakibatkan hasil sesungguhnya lebih
rendah dari hasil yang diharapkan untuk suatu spesies, dan sebaliknya lebih tinggi
dari hasil yang diharapkan untuk spesies yang lain (compensation).
Metode percobaan replacement series atau sering disebut metode seri
penggantian merupakan salah satu metode yang sering digunakan dalam kajian
kompetisi antara dua spesies yang hidup bersama (Partzsch et al. 2011). Metode
disusun dengan mengganti proporsi tanaman yang berkompetisi, tetapi total
individu dalam satuan luas lahan tetap. Tujuan penelitian ini adalah untuk
menganalisis kemampuan kompetisi gulma Echinochloa crus-galli terhadap
tanaman padi.

96
Bahan dan Metode
Penelitian dilakukan di rumah kaca Kebun Percobaan IPB Cikabayan,
pada bulan April 2009 sampai dengan Agustus 2009. Bahan tanaman yang
digunakan yaitu benih padi varietas IR-64 dan gulma E. crus-galli aksesi
Karawang (K6). Peralatan yang digunakan antara lain pot plastik berukuran 30-
40 cm (diameter-tinggi), bak semai, meteran, timbangan, dan oven.
Percobaan dilakukan dengan menggunakan rancangan acak kelompok
dengan satu faktor yaitu populasi tanaman per pot dengan tiga ulangan. Populasi
tanaman per pot terdiri atas sebelas taraf yaitu 1 gulma E. crus-galli/pot (G1),
2 gulma/pot (G2), 3 gulma/pot (G3), 4 gulma/pot (G4), 1 padi/pot (P1), 2 padi/pot
(P2), 3 padi/pot (P3), 4 padi/pot (P4), 3 padi + 1 gulma/pot (P3+G1), 2 padi +
2 gulma/pot (P2+G2), dan 1 padi + 3 gulma/pot (P1+G3). Satuan percobaan
berupa pot dengan diameter 30 cm dan tinggi 40 cm.
(G1) (G2) (G3) (G4)
(P1) (P2) (P3) (P4)
(P1+G3) (P2+G2) (P3+G1)
Gambar 17. Skema penanaman padi dan gulma E. crus-galli di pot pada percobaan replacement series
Media tanam adalah jenis tanah latosol yang berasal dari Kebun Percobaan
Sawah Baru Dramaga. Tanah dikeringanginkan dan dihaluskan, selanjutnya
dimasukkan ke dalam pot sebanyak 9 kg/ pot. Biji gulma E. crus-galli direndam
dalam larutan GA3 500 ppm selama 4 jam sebelum penyemaian pada bak semai.
Bibit gulma dan bibit padi berumur 14 hari setelah semai ditanam pada pot
percobaan sebanyak satu bibit per lubang dengan populasi sesuai perlakuan.

97
Pemupukan dilakukan 3 kali yaitu pada saat tanam, 4 minggu setelah tanam
(MST), dan 6 MST dengan dosis masing-masing 1.35 g urea/pot, 0.9 g SP-18/pot,
dan 0.9 g KCl/pot. Penyiraman dilakukan 2 hari sekali dengan ketinggian sekitar
5 cm. Penyiangan gulma selain E. crus-galli dilakukan secara manual.
Peubah yang diamati antara lain peubah pertumbuhan vegetatif yang
diamati pada saat 9 MST dan peubah pertumbuhan generatif yang diamati pada
saat panen. Peubah pertumbuhan vegetatif yang diamati antara lain tinggi
tanaman, jumlah anakan, jumlah daun, panjang duan dan lebar daun. Peubah
pertumbuhan generatif yang diamati antara lain panjang, lebar, dan luas daun
bendera, bobot biomassa tajuk dan akar, panjang akar terpanjang, panjang malai,
jumlah biji per malai, jumlah biji per pot, bobot gabah per pot, bobot 100 butir
gabah padi, dan kandungan hara tajuk. Data hasil pengamatan dianalisis
menggunakan analisis ragam dengan uji lanjut DMRT (Duncan Multiple Range
Test) pada taraf 5%.
Hasil dan Pembahasan
Kompetisi di Atas Permukaan Tanah
Tinggi tanaman. Rata-rata tinggi tanaman padi pada berbagai populasi
tidak berbeda nyata baik pada pertanaman monokultur maupun campuran. Tinggi
gulma E. crus-galli juga tidak berbeda nyata antara pertanaman monokultur dan
campuran (Tabel 42). Gulma E. crus-galli menunjukkan tinggi tanaman yang
lebih tinggi daripada tanaman padi, sehingga diduga memiliki kemampuan yang
lebih tinggi dalam kompetisi cahaya dibandingkan dengan tanaman padi.
Menurut Anten dan Hirose (1998), tanaman yang tinggi memiliki kemampuan
menangkap cahaya yang lebih besar dibandingkan dengan tanaman yang lebih
pendek.
Jumlah anakan. Jumlah anakan tanaman padi per pot tidak berbeda nyata
antar populasi tanaman, baik pada pertanaman monokultur maupun pada
pertanaman campuran, sedangkan gulma E. crus-galli pada populasi yang berbeda
menunjukkan jumlah anakan per pot yang berbeda, baik pada pertanaman
monokultur maupun pada pertanaman campuran. Pada pertanaman monokultur,
jumlah anakan gulma E. crus-galli nyata menurun mulai populasi 4 gulma/pot.

98
Pada pertanaman campuran, jumlah anakan gulma E. crus-galli tidak berbeda
nyata dengan perlakuan monokultur pada populasi 4 gulma/pot (Tabel 42). Hasil
ini menunjukkan bahwa kompetisi intraspesifik pada gulma E. crus-galli lebih
kuat dibandingkan dengan kompetisi interspesifik.
Tabel 42. Pengaruh populasi tanaman terhadap tinggi tanaman dan jumlah anakan padi dan gulma E. crus-galli pada 9 MST
Populasi per Pot Tinggi
Tanaman Padi (cm)
Tinggi Gulma (cm)
Jumlah Anakan
Padi/tanaman
Jumlah Anakan Gulma
1 padi 91.83 - 4.00 - 2 padi 89.10 - 3.00 - 3 padi 85.92 - 3.00 - 4 padi 83.54 - 2.00 -
3 padi + 1 gulma 84.62 119.03 2.33 1.3bc 2 padi + 2 gulma 88.37 114.85 2.50 1.8bc 1 padi + 3 gulma 89.43 111.57 2.33 0.8c
1 gulma - 128.17 - 4.3a 2 gulma - 116.57 - 3.2ab 3 gulma - 120.59 - 2.3abc 4 gulma - 110.71 - 1.6bc
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Jumlah daun. Pada pertanaman monokultur, jumlah daun tanaman padi
menurun mulai populasi 4 gulma/pot dibandingkan dengan populasi 1 padi/pot.
Pada pertanaman campuran, jumlah daun tanaman padi menurun pada populasi 3
padi + 1 gulma dan populasi 1 padi + 3 gulma, sedangkan pada populasi 2 padi +
2 gulma menunjukkan jumlah daun tanaman padi yang tidak berbeda nyata
dengan populasi 1 padi/pot.
Jumlah daun gulma E. crus-galli pada pertanaman monokultur mulai
menurun ketika populasi 2 gulma/pot dan peningkatan populasi lebih dari 2
gulma/pot menunjukkan jumlah daun gulma yang tidak berbeda nyata dengan
jumlah daun pada populasi 2 gulma/pot. Jumlah daun gulma E. crus-galli lebih
sedikit dibandingkan dengan jumlah daun tanaman padi (Tabel 43).
Panjang daun dan lebar daun. Panjang daun tanaman padi dan panjang
daun gulma E. crus-galli tidak berbeda nyata baik pada pertanaman monokultur
maupun pada pertanaman campuran. Rata-rata panjang daun gulma E. crus-galli
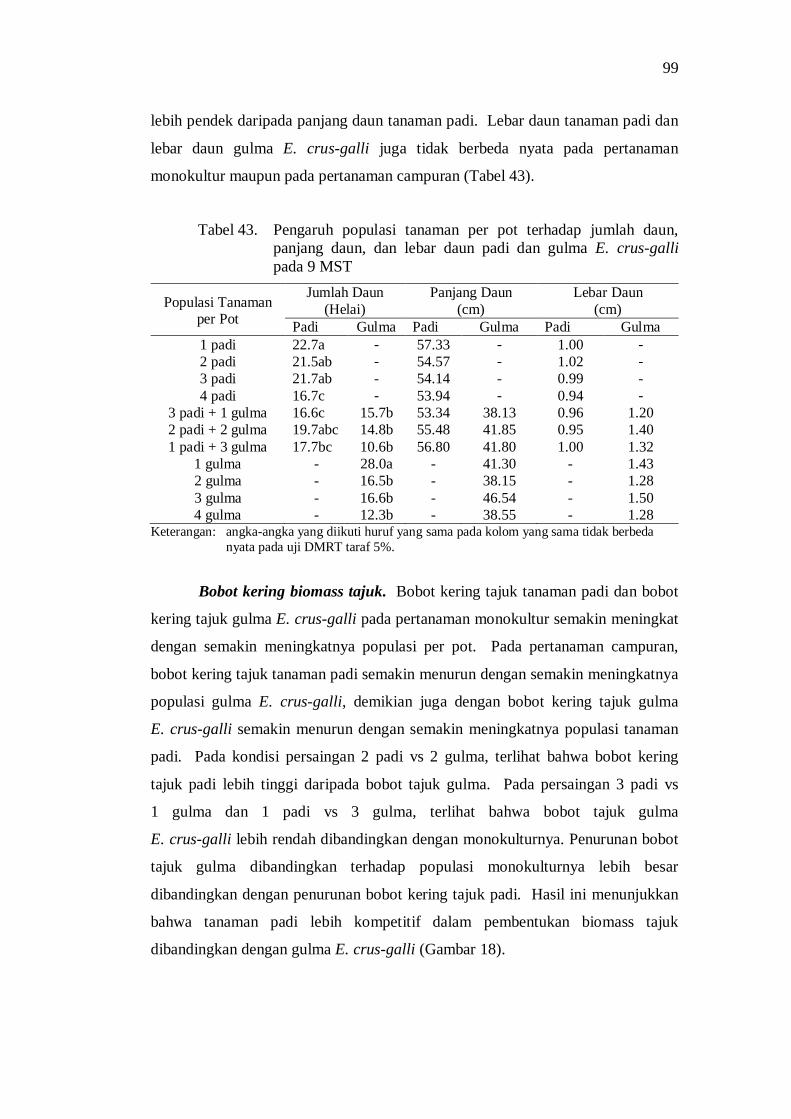
99
lebih pendek daripada panjang daun tanaman padi. Lebar daun tanaman padi dan
lebar daun gulma E. crus-galli juga tidak berbeda nyata pada pertanaman
monokultur maupun pada pertanaman campuran (Tabel 43).
Tabel 43. Pengaruh populasi tanaman per pot terhadap jumlah daun, panjang daun, dan lebar daun padi dan gulma E. crus-galli pada 9 MST
Populasi Tanaman per Pot
Jumlah Daun (Helai)
Panjang Daun (cm)
Lebar Daun (cm)
Padi Gulma Padi Gulma Padi Gulma 1 padi 22.7a - 57.33 - 1.00 - 2 padi 21.5ab - 54.57 - 1.02 - 3 padi 21.7ab - 54.14 - 0.99 - 4 padi 16.7c - 53.94 - 0.94 -
3 padi + 1 gulma 16.6c 15.7b 53.34 38.13 0.96 1.20 2 padi + 2 gulma 19.7abc 14.8b 55.48 41.85 0.95 1.40 1 padi + 3 gulma 17.7bc 10.6b 56.80 41.80 1.00 1.32
1 gulma - 28.0a - 41.30 - 1.43 2 gulma - 16.5b - 38.15 - 1.28 3 gulma - 16.6b - 46.54 - 1.50 4 gulma - 12.3b - 38.55 - 1.28
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Bobot kering biomass tajuk. Bobot kering tajuk tanaman padi dan bobot
kering tajuk gulma E. crus-galli pada pertanaman monokultur semakin meningkat
dengan semakin meningkatnya populasi per pot. Pada pertanaman campuran,
bobot kering tajuk tanaman padi semakin menurun dengan semakin meningkatnya
populasi gulma E. crus-galli, demikian juga dengan bobot kering tajuk gulma
E. crus-galli semakin menurun dengan semakin meningkatnya populasi tanaman
padi. Pada kondisi persaingan 2 padi vs 2 gulma, terlihat bahwa bobot kering
tajuk padi lebih tinggi daripada bobot tajuk gulma. Pada persaingan 3 padi vs
1 gulma dan 1 padi vs 3 gulma, terlihat bahwa bobot tajuk gulma
E. crus-galli lebih rendah dibandingkan dengan monokulturnya. Penurunan bobot
tajuk gulma dibandingkan terhadap populasi monokulturnya lebih besar
dibandingkan dengan penurunan bobot kering tajuk padi. Hasil ini menunjukkan
bahwa tanaman padi lebih kompetitif dalam pembentukan biomass tajuk
dibandingkan dengan gulma E. crus-galli (Gambar 18).
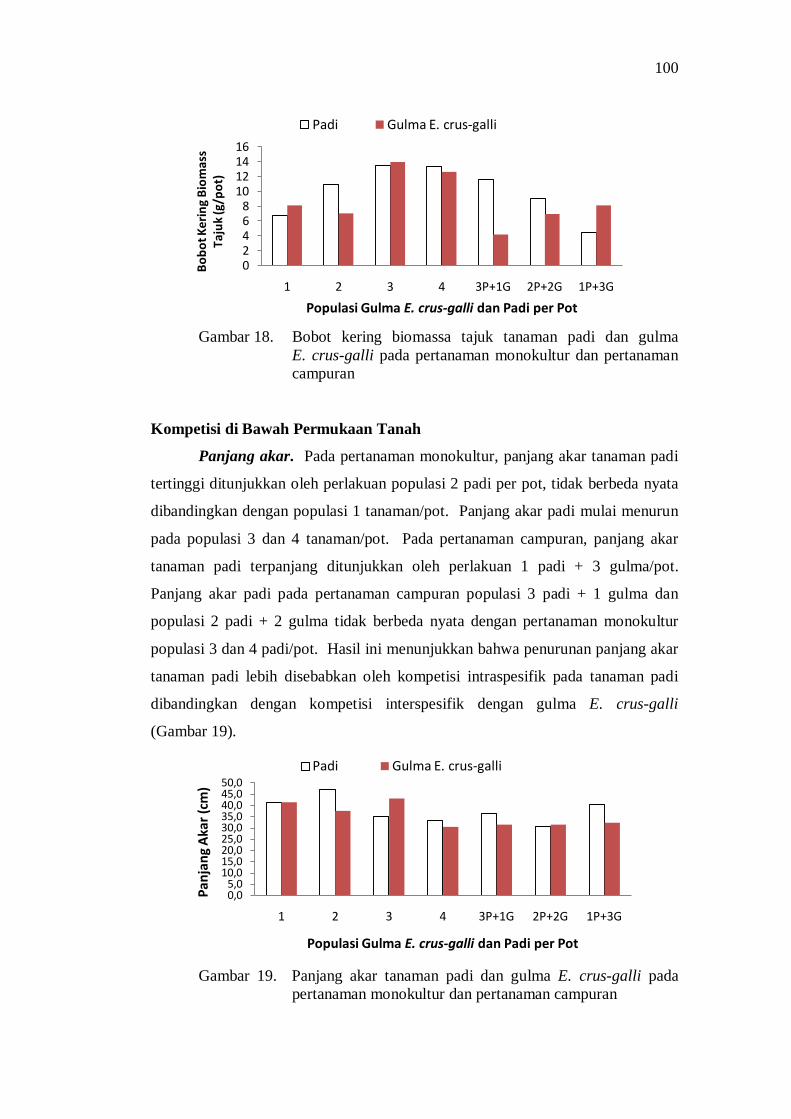
100
Gambar 18. Bobot kering biomassa tajuk tanaman padi dan gulma
E. crus-galli pada pertanaman monokultur dan pertanaman campuran
Kompetisi di Bawah Permukaan Tanah
Panjang akar. Pada pertanaman monokultur, panjang akar tanaman padi
tertinggi ditunjukkan oleh perlakuan populasi 2 padi per pot, tidak berbeda nyata
dibandingkan dengan populasi 1 tanaman/pot. Panjang akar padi mulai menurun
pada populasi 3 dan 4 tanaman/pot. Pada pertanaman campuran, panjang akar
tanaman padi terpanjang ditunjukkan oleh perlakuan 1 padi + 3 gulma/pot.
Panjang akar padi pada pertanaman campuran populasi 3 padi + 1 gulma dan
populasi 2 padi + 2 gulma tidak berbeda nyata dengan pertanaman monokultur
populasi 3 dan 4 padi/pot. Hasil ini menunjukkan bahwa penurunan panjang akar
tanaman padi lebih disebabkan oleh kompetisi intraspesifik pada tanaman padi
dibandingkan dengan kompetisi interspesifik dengan gulma E. crus-galli
(Gambar 19).
Gambar 19. Panjang akar tanaman padi dan gulma E. crus-galli pada
pertanaman monokultur dan pertanaman campuran
02468
10121416
1 2 3 4 3P+1G 2P+2G 1P+3G
Bobo
t Ker
ing
Biom
ass
Taju
k (g
/pot
)
Populasi Gulma E. crus-galli dan Padi per Pot
Padi Gulma E. crus-galli
0,05,0
10,015,020,025,030,035,040,045,050,0
1 2 3 4 3P+1G 2P+2G 1P+3G
Panj
ang
Aka
r (cm
)
Populasi Gulma E. crus-galli dan Padi per Pot
Padi Gulma E. crus-galli
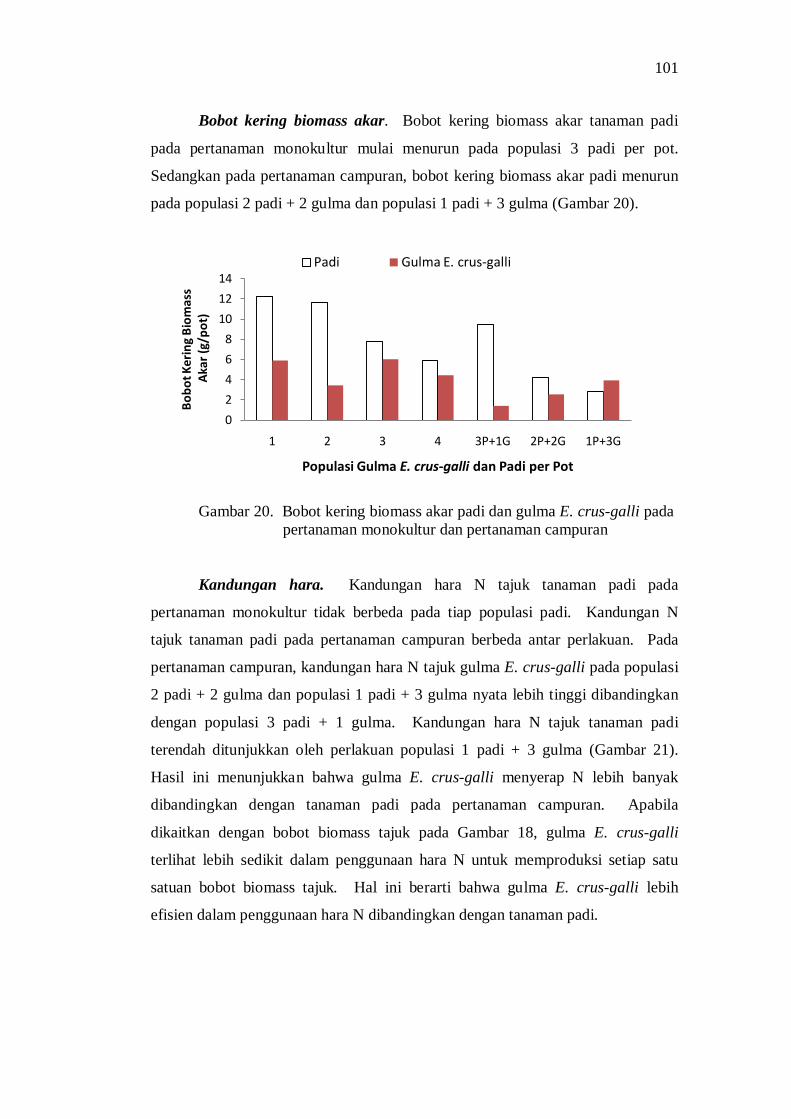
101
Bobot kering biomass akar. Bobot kering biomass akar tanaman padi
pada pertanaman monokultur mulai menurun pada populasi 3 padi per pot.
Sedangkan pada pertanaman campuran, bobot kering biomass akar padi menurun
pada populasi 2 padi + 2 gulma dan populasi 1 padi + 3 gulma (Gambar 20).
Gambar 20. Bobot kering biomass akar padi dan gulma E. crus-galli pada pertanaman monokultur dan pertanaman campuran
Kandungan hara. Kandungan hara N tajuk tanaman padi pada
pertanaman monokultur tidak berbeda pada tiap populasi padi. Kandungan N
tajuk tanaman padi pada pertanaman campuran berbeda antar perlakuan. Pada
pertanaman campuran, kandungan hara N tajuk gulma E. crus-galli pada populasi
2 padi + 2 gulma dan populasi 1 padi + 3 gulma nyata lebih tinggi dibandingkan
dengan populasi 3 padi + 1 gulma. Kandungan hara N tajuk tanaman padi
terendah ditunjukkan oleh perlakuan populasi 1 padi + 3 gulma (Gambar 21).
Hasil ini menunjukkan bahwa gulma E. crus-galli menyerap N lebih banyak
dibandingkan dengan tanaman padi pada pertanaman campuran. Apabila
dikaitkan dengan bobot biomass tajuk pada Gambar 18, gulma E. crus-galli
terlihat lebih sedikit dalam penggunaan hara N untuk memproduksi setiap satu
satuan bobot biomass tajuk. Hal ini berarti bahwa gulma E. crus-galli lebih
efisien dalam penggunaan hara N dibandingkan dengan tanaman padi.
0
2
4
6
8
10
12
14
1 2 3 4 3P+1G 2P+2G 1P+3G
Bobo
t Ker
ing
Biom
ass
Aka
r (g/
pot)
Populasi Gulma E. crus-galli dan Padi per Pot
Padi Gulma E. crus-galli
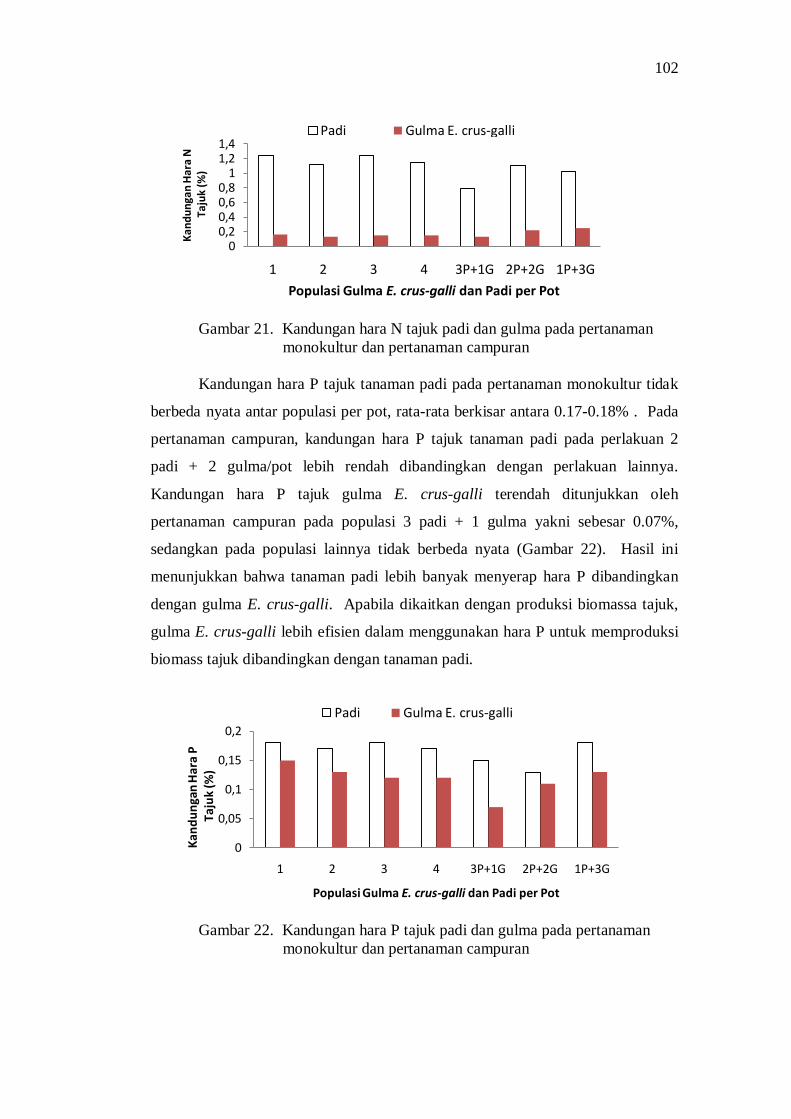
102
Gambar 21. Kandungan hara N tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran
Kandungan hara P tajuk tanaman padi pada pertanaman monokultur tidak
berbeda nyata antar populasi per pot, rata-rata berkisar antara 0.17-0.18% . Pada
pertanaman campuran, kandungan hara P tajuk tanaman padi pada perlakuan 2
padi + 2 gulma/pot lebih rendah dibandingkan dengan perlakuan lainnya.
Kandungan hara P tajuk gulma E. crus-galli terendah ditunjukkan oleh
pertanaman campuran pada populasi 3 padi + 1 gulma yakni sebesar 0.07%,
sedangkan pada populasi lainnya tidak berbeda nyata (Gambar 22). Hasil ini
menunjukkan bahwa tanaman padi lebih banyak menyerap hara P dibandingkan
dengan gulma E. crus-galli. Apabila dikaitkan dengan produksi biomassa tajuk,
gulma E. crus-galli lebih efisien dalam menggunakan hara P untuk memproduksi
biomass tajuk dibandingkan dengan tanaman padi.
Gambar 22. Kandungan hara P tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran
00,20,40,60,8
11,21,4
1 2 3 4 3P+1G 2P+2G 1P+3G
Kand
unga
n H
ara
NTa
juk
(%)
Populasi Gulma E. crus-galli dan Padi per Pot
Padi Gulma E. crus-galli
0
0,05
0,1
0,15
0,2
1 2 3 4 3P+1G 2P+2G 1P+3G
Kand
unga
n H
ara
P Ta
juk
(%)
Populasi Gulma E. crus-galli dan Padi per Pot
Padi Gulma E. crus-galli
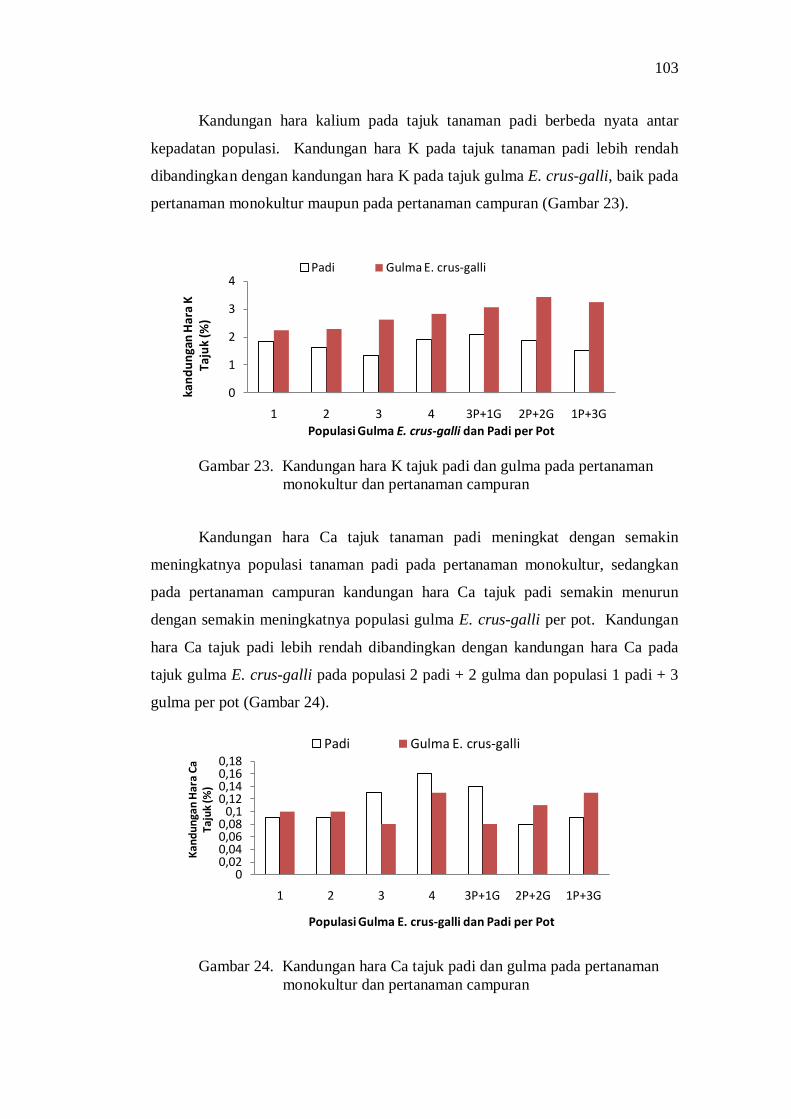
103
Kandungan hara kalium pada tajuk tanaman padi berbeda nyata antar
kepadatan populasi. Kandungan hara K pada tajuk tanaman padi lebih rendah
dibandingkan dengan kandungan hara K pada tajuk gulma E. crus-galli, baik pada
pertanaman monokultur maupun pada pertanaman campuran (Gambar 23).
Gambar 23. Kandungan hara K tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran
Kandungan hara Ca tajuk tanaman padi meningkat dengan semakin
meningkatnya populasi tanaman padi pada pertanaman monokultur, sedangkan
pada pertanaman campuran kandungan hara Ca tajuk padi semakin menurun
dengan semakin meningkatnya populasi gulma E. crus-galli per pot. Kandungan
hara Ca tajuk padi lebih rendah dibandingkan dengan kandungan hara Ca pada
tajuk gulma E. crus-galli pada populasi 2 padi + 2 gulma dan populasi 1 padi + 3
gulma per pot (Gambar 24).
Gambar 24. Kandungan hara Ca tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran
0
1
2
3
4
1 2 3 4 3P+1G 2P+2G 1P+3G
kand
unga
n H
ara
KTa
juk
(%)
Populasi Gulma E. crus-galli dan Padi per Pot
Padi Gulma E. crus-galli
00,020,040,060,08
0,10,120,140,160,18
1 2 3 4 3P+1G 2P+2G 1P+3G
Kand
unga
n H
ara
Ca
Taju
k (%
)
Populasi Gulma E. crus-galli dan Padi per Pot
Padi Gulma E. crus-galli

104
Pada pertanaman monokultur, kandungan hara Mg pada tajuk tanaman
padi tidak berbeda antar populasi tanaman per pot. Kandungan hara Mg pada
tajuk gulma semakin menurun dengan semakin meningkatnya populasi gulma.
Pada pertanaman campuran, kandungan hara Mg tajuk tanaman padi meningkat
dengan semakin meningkatnya populasi gulma E. crus-galli, demikian juga
dengan kandungan Mg pada tajuk gulma yang semakin meningkat dengan
semakin meningkatnya populasi gulma per pot (Gambar 25).
Gambar 25. Kandungan hara Mg tajuk padi dan gulma pada pertanaman monokultur dan pertanaman campuran
Pengaruh Kompetisi terhadap Produksi Biomass Total
Bobot biomass total tanaman padi tidak berbeda nyata dengan semakin
meningkatnya populasi tanaman padi pada pertanaman monokultur. Pada
pertanaman campuran, populasi gulma E. crus-galli per pot menyebabkan
penurunan bobot kering biomass total tanaman padi. Bobot kering biomass gulma
E. crus-galli pada pertanaman monokultur juga semakin meningkat dengan
meningkatnya populasi gulma. Pada pertanaman campuran, bobot gulma E. crus-
galli semakin menurun dengan semakin meningkatnya populasi tanaman padi per
pot. Pada kondisi persaingan 2 padi + 2 gulma, terlihat bahwa bobot biomass total
tanaman padi lebih tinggi dibandingkan dengan gulma E. crus-galli (Gambar 26).
00,05
0,10,15
0,20,25
0,3
1 2 3 4 3P+1G 2P+2G 1P+3G
Kand
unga
n H
ara
Mg
Taju
k (%
)
Populasi Gulma E. crus-galli dan Padi
Padi Gulma E. crus-galli
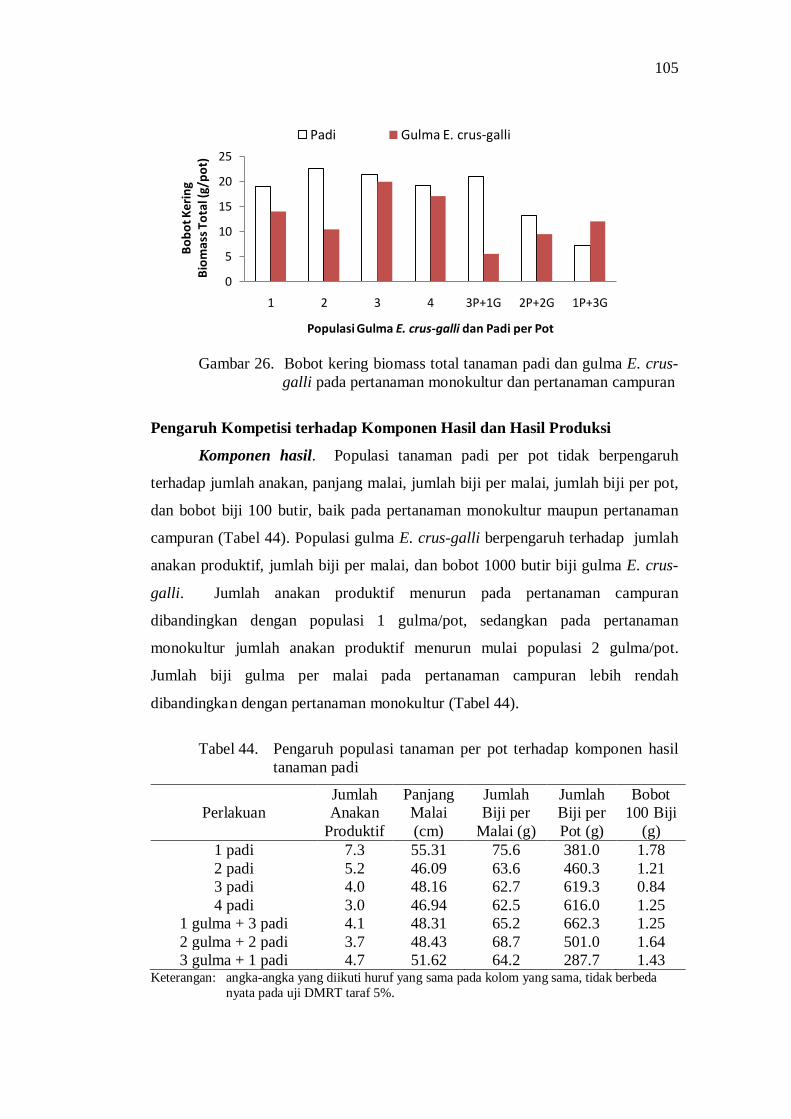
105
Gambar 26. Bobot kering biomass total tanaman padi dan gulma E. crus-galli pada pertanaman monokultur dan pertanaman campuran
Pengaruh Kompetisi terhadap Komponen Hasil dan Hasil Produksi
Komponen hasil. Populasi tanaman padi per pot tidak berpengaruh
terhadap jumlah anakan, panjang malai, jumlah biji per malai, jumlah biji per pot,
dan bobot biji 100 butir, baik pada pertanaman monokultur maupun pertanaman
campuran (Tabel 44). Populasi gulma E. crus-galli berpengaruh terhadap jumlah
anakan produktif, jumlah biji per malai, dan bobot 1000 butir biji gulma E. crus-
galli. Jumlah anakan produktif menurun pada pertanaman campuran
dibandingkan dengan populasi 1 gulma/pot, sedangkan pada pertanaman
monokultur jumlah anakan produktif menurun mulai populasi 2 gulma/pot.
Jumlah biji gulma per malai pada pertanaman campuran lebih rendah
dibandingkan dengan pertanaman monokultur (Tabel 44).
Tabel 44. Pengaruh populasi tanaman per pot terhadap komponen hasil
tanaman padi
Perlakuan Jumlah Anakan
Produktif
Panjang Malai (cm)
Jumlah Biji per
Malai (g)
Jumlah Biji per Pot (g)
Bobot 100 Biji
(g) 1 padi 7.3 55.31 75.6 381.0 1.78 2 padi 5.2 46.09 63.6 460.3 1.21 3 padi 4.0 48.16 62.7 619.3 0.84 4 padi 3.0 46.94 62.5 616.0 1.25
1 gulma + 3 padi 4.1 48.31 65.2 662.3 1.25 2 gulma + 2 padi 3.7 48.43 68.7 501.0 1.64 3 gulma + 1 padi 4.7 51.62 64.2 287.7 1.43
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama, tidak berbeda nyata pada uji DMRT taraf 5%.
0
5
10
15
20
25
1 2 3 4 3P+1G 2P+2G 1P+3G
Bobo
t Ker
ing
Biom
ass T
otal
(g/p
ot)
Populasi Gulma E. crus-galli dan Padi per Pot
Padi Gulma E. crus-galli
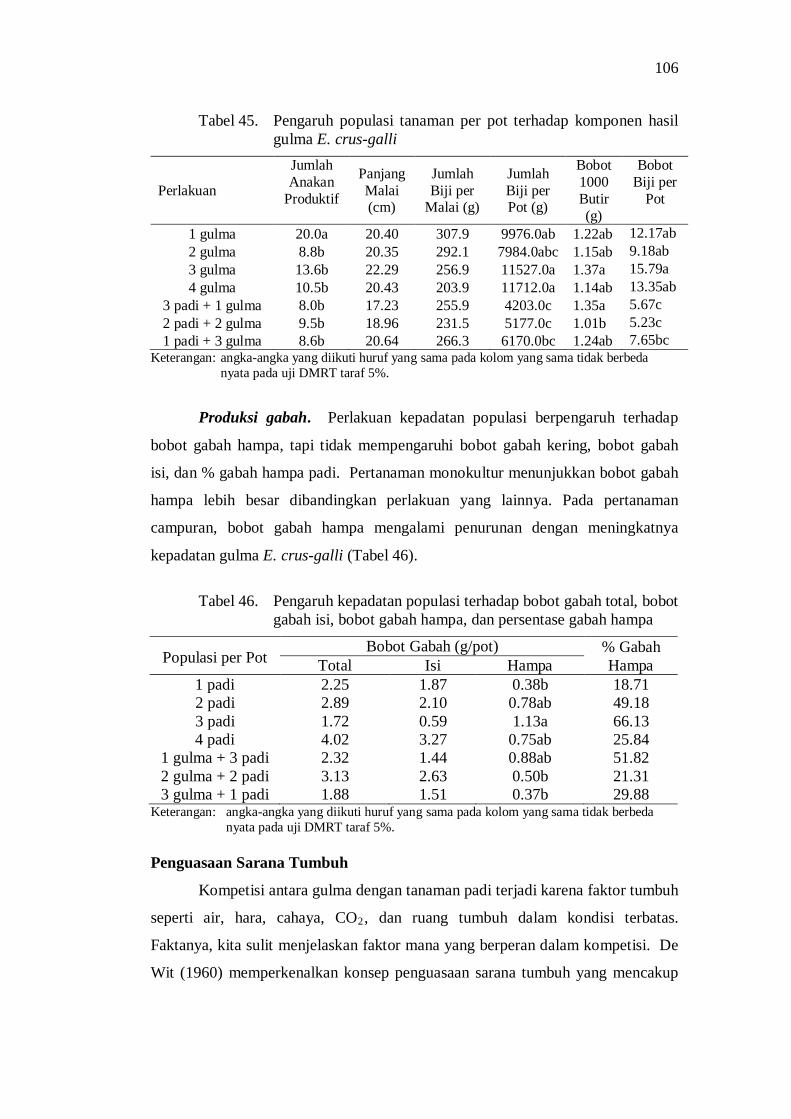
106
Tabel 45. Pengaruh populasi tanaman per pot terhadap komponen hasil gulma E. crus-galli
Perlakuan Jumlah Anakan
Produktif Panjang Malai (cm)
Jumlah Biji per
Malai (g)
Jumlah Biji per Pot (g)
Bobot 1000 Butir (g)
Bobot Biji per
Pot
1 gulma 20.0a 20.40 307.9 9976.0ab 1.22ab 12.17ab 2 gulma 8.8b 20.35 292.1 7984.0abc 1.15ab 9.18ab 3 gulma 13.6b 22.29 256.9 11527.0a 1.37a 15.79a 4 gulma 10.5b 20.43 203.9 11712.0a 1.14ab 13.35ab
3 padi + 1 gulma 8.0b 17.23 255.9 4203.0c 1.35a 5.67c 2 padi + 2 gulma 9.5b 18.96 231.5 5177.0c 1.01b 5.23c 1 padi + 3 gulma 8.6b 20.64 266.3 6170.0bc 1.24ab 7.65bc
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Produksi gabah. Perlakuan kepadatan populasi berpengaruh terhadap
bobot gabah hampa, tapi tidak mempengaruhi bobot gabah kering, bobot gabah
isi, dan % gabah hampa padi. Pertanaman monokultur menunjukkan bobot gabah
hampa lebih besar dibandingkan perlakuan yang lainnya. Pada pertanaman
campuran, bobot gabah hampa mengalami penurunan dengan meningkatnya
kepadatan gulma E. crus-galli (Tabel 46).
Tabel 46. Pengaruh kepadatan populasi terhadap bobot gabah total, bobot
gabah isi, bobot gabah hampa, dan persentase gabah hampa
Populasi per Pot Bobot Gabah (g/pot) % Gabah
Hampa Total Isi Hampa 1 padi 2.25 1.87 0.38b 18.71 2 padi 2.89 2.10 0.78ab 49.18 3 padi 1.72 0.59 1.13a 66.13 4 padi 4.02 3.27 0.75ab 25.84
1 gulma + 3 padi 2.32 1.44 0.88ab 51.82 2 gulma + 2 padi 3.13 2.63 0.50b 21.31 3 gulma + 1 padi 1.88 1.51 0.37b 29.88
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Penguasaan Sarana Tumbuh
Kompetisi antara gulma dengan tanaman padi terjadi karena faktor tumbuh
seperti air, hara, cahaya, CO2, dan ruang tumbuh dalam kondisi terbatas.
Faktanya, kita sulit menjelaskan faktor mana yang berperan dalam kompetisi. De
Wit (1960) memperkenalkan konsep penguasaan sarana tumbuh yang mencakup

107
semua faktor yang terlibat dalam kompetisi, yang dirumuskan dengan persamaan
hiperbolik Y = ( .Ymax
Perhitungan penguasaan sarana tumbuh didasarkan pada jumlah biji per
pot. Pada pertanaman monokultur, produksi biji tanaman padi mengikuti
persamaan garis linear y = 1.387x + 1.286, dimana y = (1/Y), dan x = (1/X),
Y=hasil nyata atau jumlah biji padi yang dihasilkan pada tiap populasi, X =
populasi tanaman padi per pot. Nilai populasi X dimasukkan ke dalam persamaan
garis linear tersebut, sehingga diperoleh nilai Y
. Persamaan linear resiprokal dari persamaan tersebut
yaitu , dimana Y = hasil nyata, d = densitas tumbuhan,
Ymax = hasil maksimal, dan b = kemampuan penguasaan sarana tumbuh.
Penguasaan sarana tumbuh dirumuskan dengan PST = (Y/Ymax) x 100%.
dugaan jumlah biji yang dihasilkan
oleh tanaman padi pada pertanaman monokultur. Berdasarkan persamaan linear
tersebut, diperoleh nilai Ymax
sebesar 777.6 dan nilai b sebesar 0.94, serta nilai Y
dugaan pda tiap-tiap populasi tanaman padi per pot (Tabel 47).
Tabel 47. Hasil dugaan produksi biji tanaman padi pada pertanaman monokultur
Populasi Padi (X) Jumlah Biji Padi (Y) Ydugaan
1 381 374 2 460 505 3 619 572 4 616 612
Produksi biji gulma E. crus-galli per pot pada pertanaman monokultur
mengikuti persamaan garis linear y = 0.178x + 0.901, dengan perhitungan yang
sama dengan tanaman padi diperoleh nilai Ymax
sebesar 11 098.8 dan nilai b
sebesar 5.06, serta nilai Y dugaan pada tiap-tiap populasi gulma per pot (Tabel
48). Nilai b pada gulma E. crus-galli lebih besar dibandingkan dengan nilai b
pada tanaman padi. Hasil ini menunjukkan bahwa gulma E. crus-galli lebih
menguasai sarana tumbuh dibandingkan dengan tanaman padi.
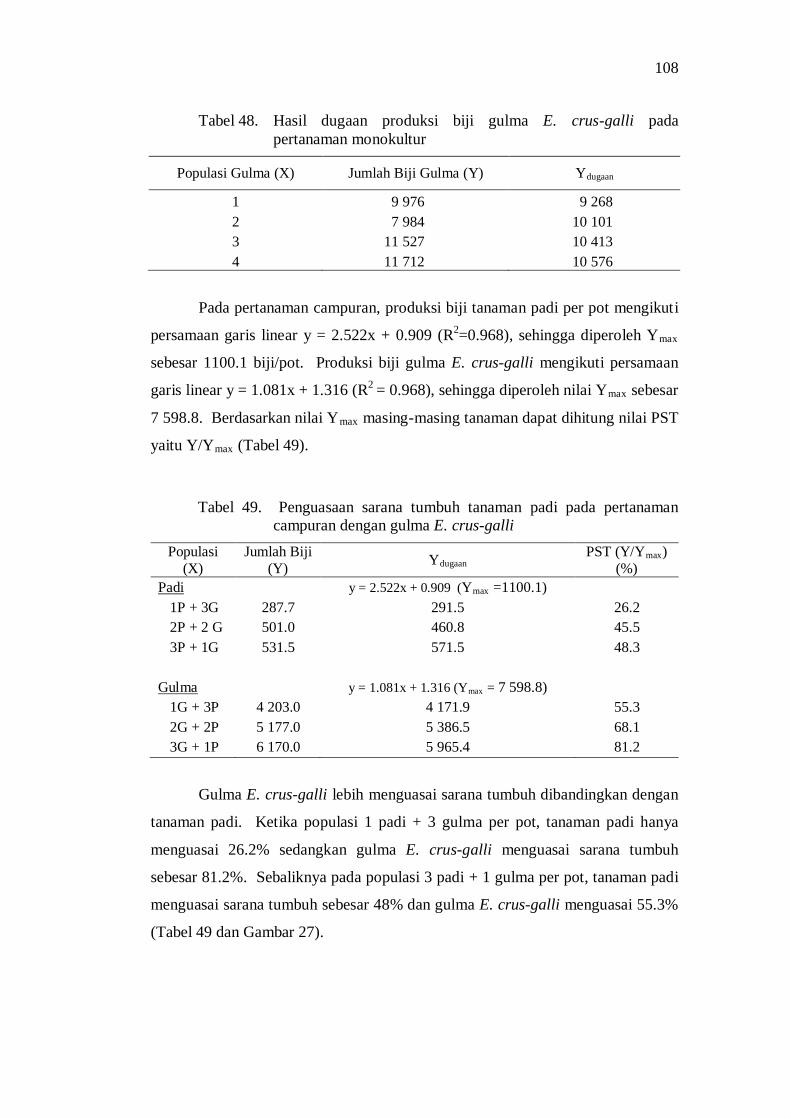
108
Tabel 48. Hasil dugaan produksi biji gulma E. crus-galli pada pertanaman monokultur
Populasi Gulma (X) Jumlah Biji Gulma (Y) Ydugaan
1 9 976 9 268 2 7 984 10 101 3 11 527 10 413 4 11 712 10 576
Pada pertanaman campuran, produksi biji tanaman padi per pot mengikuti
persamaan garis linear y = 2.522x + 0.909 (R2=0.968), sehingga diperoleh Ymax
sebesar 1100.1 biji/pot. Produksi biji gulma E. crus-galli mengikuti persamaan
garis linear y = 1.081x + 1.316 (R2 = 0.968), sehingga diperoleh nilai Ymax sebesar
7 598.8. Berdasarkan nilai Ymax masing-masing tanaman dapat dihitung nilai PST
yaitu Y/Ymax
(Tabel 49).
Tabel 49. Penguasaan sarana tumbuh tanaman padi pada pertanaman campuran dengan gulma E. crus-galli
Populasi (X)
Jumlah Biji (Y) Y PST (Y/Y
dugaan max(%)
)
Padi y = 2.522x + 0.909 (Ymax =1100.1) 1P + 3G 287.7 291.5 26.2 2P + 2 G 501.0 460.8 45.5 3P + 1G 531.5 571.5 48.3
Gulma y = 1.081x + 1.316 (Ymax = 7 598.8) 1G + 3P 4 203.0 4 171.9 55.3 2G + 2P 5 177.0 5 386.5 68.1 3G + 1P 6 170.0 5 965.4 81.2
Gulma E. crus-galli lebih menguasai sarana tumbuh dibandingkan dengan
tanaman padi. Ketika populasi 1 padi + 3 gulma per pot, tanaman padi hanya
menguasai 26.2% sedangkan gulma E. crus-galli menguasai sarana tumbuh
sebesar 81.2%. Sebaliknya pada populasi 3 padi + 1 gulma per pot, tanaman padi
menguasai sarana tumbuh sebesar 48% dan gulma E. crus-galli menguasai 55.3%
(Tabel 49 dan Gambar 27).
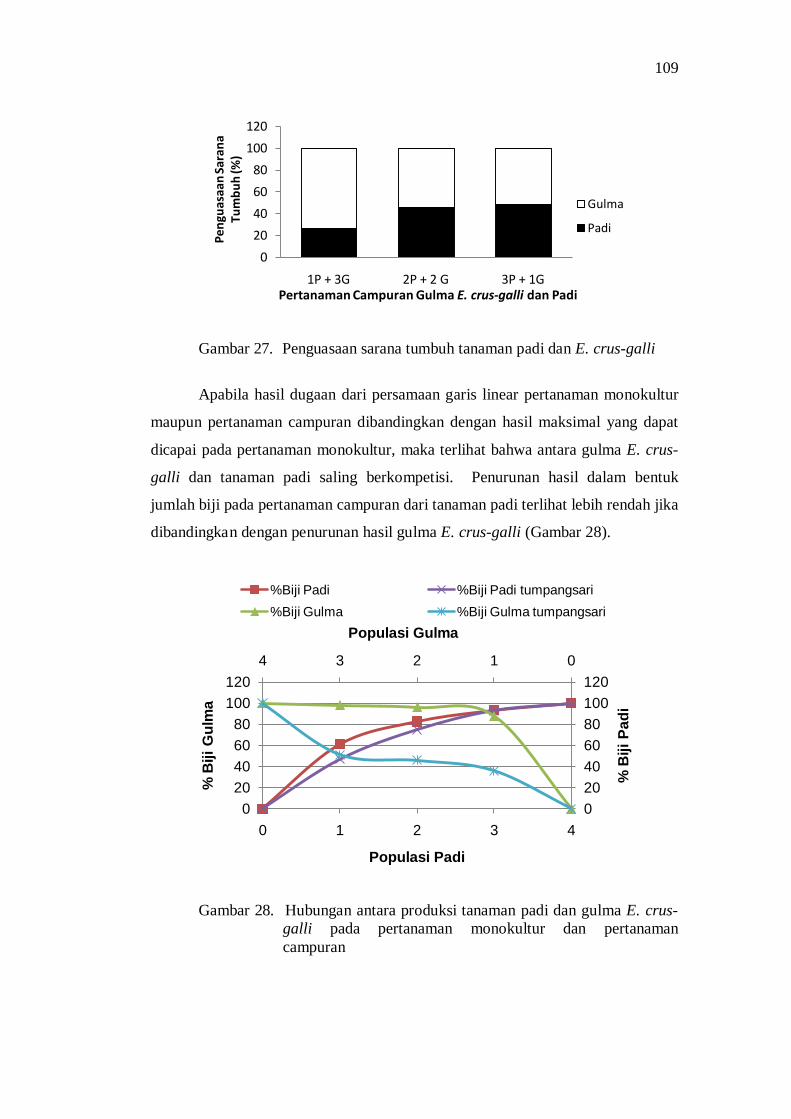
109
Gambar 27. Penguasaan sarana tumbuh tanaman padi dan E. crus-galli
Apabila hasil dugaan dari persamaan garis linear pertanaman monokultur
maupun pertanaman campuran dibandingkan dengan hasil maksimal yang dapat
dicapai pada pertanaman monokultur, maka terlihat bahwa antara gulma E. crus-
galli dan tanaman padi saling berkompetisi. Penurunan hasil dalam bentuk
jumlah biji pada pertanaman campuran dari tanaman padi terlihat lebih rendah jika
dibandingkan dengan penurunan hasil gulma E. crus-galli (Gambar 28).
Gambar 28. Hubungan antara produksi tanaman padi dan gulma E. crus-
galli pada pertanaman monokultur dan pertanaman campuran
0
20
40
60
80
100
120
1P + 3G 2P + 2 G 3P + 1G
Peng
uasa
an S
aran
a Tu
mbu
h (%
)
Pertanaman Campuran Gulma E. crus-galli dan Padi
Gulma
Padi
01234
020406080100120
020406080
100120
0 1 2 3 4
Populasi Gulma%
Biji
Pad
i
% B
iji G
ulm
a
Populasi Padi
%Biji Padi %Biji Padi tumpangsari%Biji Gulma %Biji Gulma tumpangsari

110
Koefisien Pendesakan
Berdasarkan jumlah biji per pot, populasi 3 padi + 1 gulma per pot dan
populasi 2 padi + 2 gulma menunjukkan koefisien pendesakan tanaman padi
terhadap gulma (KPp-e) lebih besar dibandingkan dengan koefisien pendesakan
gulma terhadap tanaman padi (KPe-p). Hal ini berarti bahwa pada populasi
tersebut tanaman padi lebih kuat dalam berkompetisi dibandingkan dengan gulma
E. crus-galli. Pada populasi 1 padi + 3 gulma per pot, KPe-p lebih besar
dibandingkan dengan KPp-e (Tabel 50) atau gulma E. crus-galli lebih kuat
berkompetisi dibandingkan dengan tanaman padi. Apabila dikaitkan dengan
persamaan garis dugaan produksi pada pertanaman campuran, maka kombinasi
populasi yang populasi tanaman padi lebih rendah dari populasi gulma akan
menghasilkan KPe-p lebih besar dibandingkan dengan KPp-e
, artinya tanaman padi
akan kalah berkompetisi terhadap gulma E. crus-galli.
Tabel 50. Nilai koefisien pendesakan (KP) pada pertanaman campuran padi dengan gulma E. crus-galli
Perlakuan KP KPp-e e-p
3 Padi + 1 Gulma 18.74 0.19 2 Padi + 2 Gulma 4.36 0.79 1 Padi + 3 Gulma 0.29 3.34
Nilai Agresivitas (A)
Agresivitas menunjukkan kemampuan kompetisi suatu spesies terhadap
spesies lainnya dalam interaksi antar spesies. Nilai agresivitas dihitung
berdasarkan bobot kering biomass total. Hasil percobaan menunjukkan bahwa
nilai agresivitas tanaman padi terhadap gulma E. crus-galli (Apadi-gulma) dan
agresivitas gulma E. crus-galli terhadap tanaman padi (Agulma-padi) tidak sama
dengan nol yang berarti bahwa masing-masing spesies memiliki kemampuan
kompetisi yang berbeda. Pada populasi 3 padi + 1 gulma per pot, nilai Apadi-gulma
bernilai positif (0.611), sedangkan nilai Agulma-padi bernilai negatif (-0.611). Hal
ini menunjukkan bahwa pada populasi 3 padi + 1 gulma, tanaman padi lebih
kompetitif dibandingkan dengan gulma E. crus-galli. Pada populasi 2 padi + 2
gulma dan populasi 1 padi + 3 gulma, nilai Apadi-gulma bernilai negatif yaitu

111
berturut-turut -0.268 dan -0.183, sedangkan nilai Agulma-padi
bernilai positif yaitu
0.268 dan 0.183. Hal ini berarti bahwa pada populasi 2 padi + 2 gulma dan
populasi 1 padi + 3 gulma, gulma E. crus-galli lebih kompetitif dibandingkan
dengan tanaman padi (Tabel 51).
Tabel 51. Nilai agresivitas tanaman padi dan gulma E. crus-galli pada pertanaman campuran
Perlakuan Aggresivitas A Apadi – gulma gulma-padi
3 padi + 1 gulma 0.611 -0.611 2 padi + 2 gulma -0.268 0.268 1 padi + 3 gulma -0.183 0.183
Pembahasan
Kompetisi intraspesifik terjadi baik pada tanaman padi maupun pada
gulma E. crus-galli apabila populasi semakin meningkat pada pertanaman
monokultur. Kompetisi intraspesifik pada tanaman padi ditunjukkan dengan
penurunan jumlah daun tanaman padi mulai populasi 4 tanaman padi/pot (Tabel
43), penurunan panjang akar mulai populasi 3 padi/pot (Gambar 19), penurunan
bobot kering biomass akar mulai populasi 3 padi/pot (Gambar 20). Kompetisi
intraspesifik pada gulma E. crus-galli ditunjukkan dengan penurunan jumlah daun
mulai populasi 2 gulma/pot dibandingkan dengan populasi 1 individu/pot (Tabel
43), dan penurunan kandungan hara Mg pada tajuk mulai populasi 3 gulma/pot
(Gambar 25). Hasil ini menunjukkan bahwa kompetisi intraspesifik pada gulma
E. crus-galli lebih kuat dibandingkan dengan kompetisi intraspesifik pada
tanaman padi.
Kompetisi interspesifik terjadi antara tanaman padi dengan gulma E. crus-
galli ketika gulma dan tanaman padi hidup bersama dalam kondisi sumberdaya
terbatas. Kompetisi antara gulma E. crus-galli dan tanaman padi terjadi baik di
atas permukaan tanah maupun di bawah permukaan tanah (rhizosfer).
Kompetisi interspesifik di atas permukaan tanah antara gulma E. crus-galli
dengan tanaman padi diantaranya ditunjukkan dengan penurunan bobot kering
biomass tajuk baik pada tanaman padi maupun pada gulma E. crus-galli
dibandingkan dengan pertanaman monokulturnya. Pada pertanaman campuran,

112
penurunan bobot biomass tajuk pada tanaman padi lebih besar dibandingkan
dengan penurunan bobot biomass gulma E. crus-galli. Gulma E. crus-galli
menunjukkan tinggi gulma yang lebih tinggi, panjang daun lebih pendek, dan
lebar daun yang lebih lebar daripada tanaman padi. Dengan karakter yang
demikian, gulma E. crus-galli memiliki kemampuan kompetisi yang lebih baik
dalam menangkap cahaya matahari dibandingkan dengan tanaman padi, sehingga
penurunan bobot kering biomass tajuk tanaman padi lebih besar dibandingkan
dengan gulma. Menurut Anten dan Hirose (1998) tanaman yang pertumbuhannya
besar akan menangkap cahaya yang lebih banyak sehingga memiliki kemampuan
kompetisi interspesifik yang lebih besar.
Kompetisi interspesifik di bawah permukaan tanah antara gulma E. crus-
galli dengan tanaman padi ditunjukkan dengan penurunan bobot kering biomass
akar baik pada tanaman padi maupun gulma E. crus-galli dibandingkan dengan
tanaman monokulturnya. Penurunan bobot kering biomass akar tanaman padi
lebih besar dibandingkan dengan bobot kering biomass akar gulma E. crus-galli.
Kompetisi bawah tanah juga terjadi dalam hal penggunaan hara. Gulma E. crus-
galli lebih efisien dalam penggunaan hara N dan P untuk memproduksi
biomassa, namun gulma E. crus-galli menyerap hara Mg dan K yang lebih banyak
dibandingkan dengan tanaman padi. Menurut Gibson et al. (1999), kompetisi di
bagian rhizosfer atau perakaran memainkan peranan penting dibandingkan dengan
kompetisi di bagian tajuk. Pada populasi tinggi, besarnya persaingan tanaman
akan meningkat (Liu et al. 2008). Tanaman yang memiliki ukuran lebih besar
akan mendapatkan proporsi sumberdaya yang lebih besar daripada tanaman yang
berukuran kecil (Yuan et al. 2004).
Kejadian kompetisi antara gulma E. crus-galli dan tanaman padi dapat
diketahui berdasarkan peubah-peubah kompetisi, diantaranya penguasaan sarana
tumbuh, koefisien pendesakan, dan agresivitas. Penghitungan peubah-peubah
tersebut didasarkan pada produksi biji per pot. Penghitungan penguasaan sarana
tumbuh berdasarkan produksi biji per pot menunjukkan adanya kejadian
kompetisi antara tanaman padi dengan gulma E. crus-galli. Gulma E. crus-galli
menguasai sarana tumbuh yang lebih besar dibandingkan dengan tanaman padi
pada pertanaman campuran (Tabel 49).

113
Berdasarkan nilai koefisien pendesakan terlihat bahwa gulma E. crus-galli
dan tanaman padi pada pertanaman campuran menunjukkan kompetisi yang
ditunjukkan dengan adanya penurunan produksi per pot jika dibandingkan dengan
pertanaman monokulturnya. Berdasarkan Gambar 28 terlihat bahwa penurunan
produksi biji per pot pada tanaman padi lebih rendah dibandingkan dengan gulma
E. crus-galli ketika populasi tanaman padi lebih banyak daripada populasi
gulmanya, sedangkan pada populasi gulma E. crus-galli yang lebih tinggi dari
tanaman padi maka penurunan produksi biji per pot pada tanaman padi lebih besar
dibandingkan dengan gulma E. crus-galli. Hasil ini menunjukkan bahwa gulma
E. crus-galli lebih kompetitif terhadap tanaman padi ketika populasi gulma E.
crus-galli lebih banyak dibandingkan dengan tanaman padi.
Perhitungan nilai agresivitas berdasarkan produksi biomass menunjukkan
bahwa nilai agresivitas gulma E. crus-galli positif pada saat populasi gulma dan
tanaman padi pada proporsi yang sama ataupun populasi gulma lebih banyak
dibandingkan dengan populasi tanaman padi. Nilai agresivitas positif
menunjukkan bahwa gulma E. crus-galli lebih dominan dalam kompetisi
dibandingkan dengan tanaman padi dalam produksi biomass (Tabel 51).
Kesimpulan dan Saran
Kompetisi intraspesifik terjadi baik pada tanaman padi maupun pada
gulma E.crus-galli dengan adanya peningkatan populasi per pot. Kompetisi
intraspesifik pada gulma E. crus-galli lebih besar dibandingkan dengan kompetisi
intraspesifik pada tanaman padi. Kompetisi interspesifik terjadi antara gulma E.
crus-galli dan tanaman padi ketika hidup bersama dalam pertanaman campuran.
Kekuatan kompetisi masing-masing tergantung pada kepadatan populasi gulma.
Kompetisi terjadi baik di atas permukaan tanah maupun di bawah permukaan
tanah.
Berdasarkan penguasaan sarana tumbuh, gulma E. crus-galli menguasai
sarana tumbuh lebih besar dibandingkan dengan tanaman padi pada pertanaman
campuran. Berdasarkan nilai koefisien pendesakan, gulma E. crus-galli memiliki
derajat kompetisi yang lebih besar terhadap tanaman padi ketika populasi gulma
lebih tinggi daripada populasi tanaman padi. Pada populasi yang seimbang antara

114
padi dan gulma ataupun populasi padi lebih tinggi dari gulma E. crus-galli,
tanaman padi lebih kuat berkompetisi. Berdasarkan nilai agresivitas, gulma E.
crus-galli lebih kuat berkompetisi dibandingkan tanaman padi ketika populasi
padi dan gulma seimbang ataupun populasi gulma lebih tinggi daripada populasi
tanaman padi.

KAJIAN FISIOLOGI KOMPETISI ANTARA TANAMAN PADI SAWAH DENGAN GULMA
Echinochloa crus-galli
ABSTRAK
Tiap varietas padi memiliki pertumbuhan dan produksi serta kemampuan
kompetisi yang berbeda terhadap gulma E. crus-galli. Penelitian bertujuan untuk
menganalisis fisiologi kompetisi beberapa varietas unggul padi sawah terhadap
gulma E. crus-galli. Penelitian dilakukan dengan rancangan split plot dengan tiga
ulangan. Varietas padi sebagai petak utama terdiri atas empat varietas yaitu
varietas Ciherang, Fatmawati, Inpari 6 JT 6 JT, dan hibrida SL 8 SHS. Populasi
gulma sebagai anak petak terdiri atas 0, 4, 8, dan 12 gulma E. crus-galli/m2
.
Satuan percobaan berupa petak berukuran 4 m x 5 m. Hasil percobaan
menunjukkan bahwa proses fisiologi tanaman padi sawah menurun akibat
kompetisi dengan gulma E. crus-galli. Kompetisi menyebabkan peningkatan leaf
area ratio (LAR) dan penurunan net assimilation rate (NAR), relative growth
rate (RGR) dan crop growth rate (RGR) pada tanaman padi. Varietas Fatmawati
menunjukkan varietas yang toleran, sedangkan varietas hibrida SL 8 SHS
menunjukkan varietas yang sensitif terhadap kompetisi gulma E. crus-galli.
Kompetisi gulma E. crus-galli terhadap tanaman padi sawah dipengaruhi oleh
kepadatan populasi gulma E. crus-galli. Semakin tinggi populasi gulma E. crus-
galli, pertumbuhan dan produksi tanaman padi semakin menurun.
Kata kunci : Echinochloa crus-galli, kompetisi, fisiologi, varietas padi, populasi gulma.

116
STUDY OF PHYSIOLOGICAL COMPETITION OF RICE TOWARD Echinochloa crus-
galli WEED
ABSTRACT
Each rice variety has the growth and production and the different ability of competition against E. crus-galli weeds. The objective of the research was to analyze the physiological competition of some high yielding varieties of paddy to E. crus-galli weed. The research was conducted with a split plot design with three replications. Rice varieties as main plots consisted of four varieties i.e. Ciherang, Fatmawati, Inpari 6 JT 6 JT, and SL 8 SHS hybrid varieties. Weed population as a subplot consisted of 0, 4, 8, and 12 E. crus-galli/m2
. The results indicate that the physiological process of rice plant decreased due to competition with weeds E. crus-galli. Competition increased leaf area ratio (LAR) and a decreased net assimilation rate (NAR), relative growth rate (RGR) and the crop growth rate (RGR). Fatmawati showed tolerant varieties, while SL 8 SHS showed sensitive varieties to weed competition of E. crus-galli. Competition of E. crus-galli weed to rice plants was affected by the population densities of E. crus-galli weed. The higher weed populations of E. crus-galli decreased growth and production of rice plants.
Keywords : Echinochloa crus-galli, competition, physiology, rice variety, weed population.

117
Pendahuluan
Luas pertanaman padi di Indonesia diperkirakan mencapai 12 juta hektar
yang tersebar di berbagai tipologi lahan, antara lain lahan sawah (5.1 juta ha),
lahan tandah hujan (2.1 juta ha), ladang (1.2 juta ha) dan lahan pasang surut.
Lebih dari 90% produksi beras nasional dihasilkan dari lahan sawah dan lebih dari
80% total areal pertanaman padi sawah telah ditanami varietas unggul (Badan
Pusat Statistik, 2010).
E. crus-galli merupakan gulma yang menjadi problem utama pada
budidaya tanaman padi sawah dan merupakan penyebab kehilangan hasil produksi
yang utama pada produksi padi sawah (Gealy et al. 2003; Haefele et al. 2004).
Penurunan produksi padi dapat mencapai 46-59% (Sultana, 2000; Chin, 2001;
Guntoro et al. 2009), 57-95% (Ahn dan Chung 2000), 97% (Islam dan Karim
2003). Tindall et al. (2003) melaporkan bahwa penurunan hasil produksi padi
terjadi karena penurunan jumlah anakan, jumlah malai, dan jumlah gabah per
malai.
Varietas unggul padi sawah merupakan kunci keberhasilan peningkatan
produksi padi di Indonesia. Saat ini penanaman varietas unggul padi meliputi
lebih dari 80% total areal pertanaman padi di Indonesia (Susanto et al., 2003).
Tiap varietas memiliki potensi produksi yang berbeda dan diduga memiliki
kemampuan mempertahankan pertumbuhan dan produksinya dari gangguan
gulma E. crus-galli di lapangan. Di wilayah Propinsi Jawa Barat, luas tanaman
padi verietas Ciherang merupakan varietas yang terluas ditanam yang mencapai
sekitar 318 ribu hektar. Informasi tentang kemampuan varietas padi dalam
berkompetisi terhadap gulma E. crus-galli masih sangat terbatas di Indonesia.
Penelitian bertujuan menganalisis fisiologi kompetisi beberapa varietas unggul
padi sawah dengan gulma E. crus-galli.
Bahan dan Metode
Penelitian dilakukan di lahan sawah Desa Parakan, Kecamatan Ciomas,
Kabupaten Bogor yang berada pada ketinggian 250 m dpl dengan jenis tanah
latosol mulai bulan Desember 2010 hingga bulan Mei 2011. Bahan tanaman yang

118
digunakan yaitu benih varietas Inpari 6 JT 6 JT, Fatmawati, Ciherang, dan SL 8
SHS. Gulma yang digunakan adalah aksesi asal Karawang (K6).
Percobaan dilakukan menggunakan rancangan petak terbagi (split-plot
design) dalam rancangan acak kelompok dengan tiga ulangan. Percobaan terdiri
atas 2 faktor yaitu varietas padi dan populasi gulma. Varietas padi sebagai petak
utama terdiri atas varietas Inpari 6 JT 6 JT (V1), Fatmawati (V2), Ciherang (V3),
dan hibrida SL 8 SHS (V4). Populasi gulma E. crus-galli sebagai anak petak
terdiri atas empat taraf yaitu 0, 4, 8, dan 12 bibit gulma/m2. Satuan percobaan
berupa petak berukuran 4 m x 5 m. Data dianalisis menggunakan analisis ragam
dengan uji lanjut Duncan’s Multiple Range Test (DMRT) pada taraf nyata 5%.
Pengolahan lahan dilakukan dua kali yaitu pembajakan pada saat 2 minggu
sebelum tanam dan penghalusan tanah dengan menggunakan cangkul pada saat 1
minggu sebelum tanam. Benih padi direndam 48 jam dan ditiriskan selama 24
jam selanjutnya disemai pada lahan semai basah berukuran 1.2 m x 5 m. Benih
E. cruss-galli direndam dengan air hangat selama 24 jam dan ditiriskan selama 24
jam, kemudian disemai pada bedengan berukuran 0.5 m x 5 m. Bibit padi
berumur 2 minggu setelah semai ditanam sebanyak 2 bibit per lubang tanam
dengan jarak tanam 25 cm x 25 cm. Bibit E. crus-galli berumur 2 minggu setelah
semai ditanam sesuai dengan perlakuan, yaitu populasi 0, 4, 8 dan 12 bibit
Pemupukan pertama dilakukan pada saat tanam dengan dosis 100 kg
Urea/ha, 100 kg SP-18/ha, dan 80 kg KCl/ha. Pemupukan kedua pada saat 4 MST
dengan dosis 90 kg Urea/ha. Pemupukan ketiga pada saat 8 MST dengan dosis 90
kg Urea/ha dan 20 kg KCl/ha. Pengendalian gulma selain gulma E. crus-galli
dilakukan secara manual pada saat 3 dan 6 MST. Pengairan dilakukan dengan
menjaga ketinggian air tetap 5 cm dari permukaan tanah sejak 2 MST sampai dua
minggu sebelum panen. Panen padi dilakukan pada umur 115 hari setelah semai
atau 96 hari setelah pindahtanam.
E. crus-galli per m2 (Gambar 29).

119
Gambar 29. Perlakuan populasi gulma E. crus-galli/m2
Peubah yang diamati pada tanaman padi antara lain tinggi tanaman, jumlah
anakan dan jumlah daun, bobot kering akar dan tajuk pada saat 4, 8, dan 14 MST
(saat panen), indeks luas daun, kandungan hara daun pada saat 8 MST, waktu
heading, jumlah anakan produktif, panjang malai, jumlah bulir per malai, bobot
gabah kering panen (GKP) dan gabah kering giling (GKG), bobot gabah 1 000
butir, dan mutu gabah. Pengamatan pada E. crus-galli antara lain tinggi gulma,
jumlah daun dan anakan, bobot kering akar dan tajuk, jumlah anakan produktif,
panjang malai, jumlah biji per malai, bobot biji 1000 butir, harvest indeks (HI).
Pengamatan peubah fisiologi antara lain
Leaf Area Ratio (LAR), Net Assimilation
Rate (NAR), Relative Growth Rate (RGR), dan Crop Growth Rate (CGR).
Keterangan : LA = luas daun total W1 = bobot kering total pada saat T1 W2 = bobot kering total pada saat T2 W = bobot kering total tanaman Ln = natural logarithm LA1 = Leaf Area pada saat T1 LA2 = Leaf Area pada saat T2
0 E. crus-galli/m2 4 E. crus-galli/m2
12 E. crus-galli/m2 8 E. crus-galli/m2
Keterangan: : tanaman padi : E. crus-galli
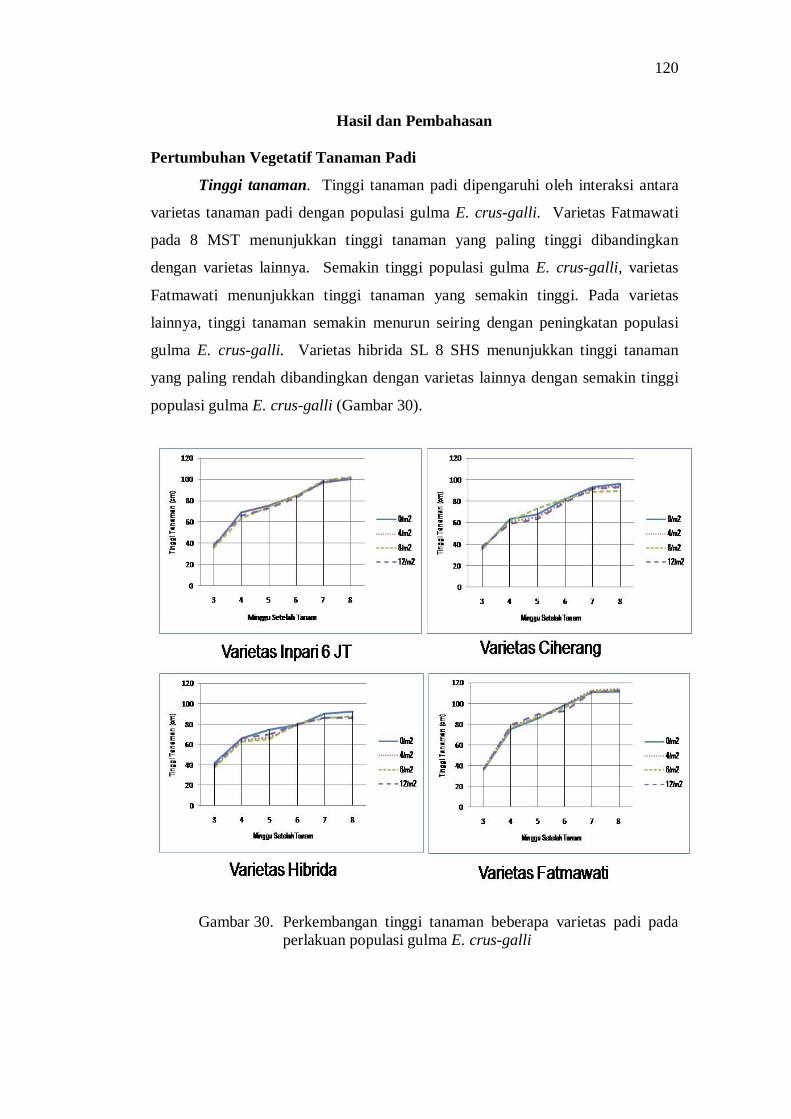
120
Hasil dan Pembahasan
Pertumbuhan Vegetatif Tanaman Padi
Tinggi tanaman. Tinggi tanaman padi dipengaruhi oleh interaksi antara
varietas tanaman padi dengan populasi gulma E. crus-galli. Varietas Fatmawati
pada 8 MST menunjukkan tinggi tanaman yang paling tinggi dibandingkan
dengan varietas lainnya. Semakin tinggi populasi gulma E. crus-galli, varietas
Fatmawati menunjukkan tinggi tanaman yang semakin tinggi. Pada varietas
lainnya, tinggi tanaman semakin menurun seiring dengan peningkatan populasi
gulma E. crus-galli. Varietas hibrida SL 8 SHS menunjukkan tinggi tanaman
yang paling rendah dibandingkan dengan varietas lainnya dengan semakin tinggi
populasi gulma E. crus-galli (Gambar 30).
Gambar 30. Perkembangan tinggi tanaman beberapa varietas padi pada
perlakuan populasi gulma E. crus-galli
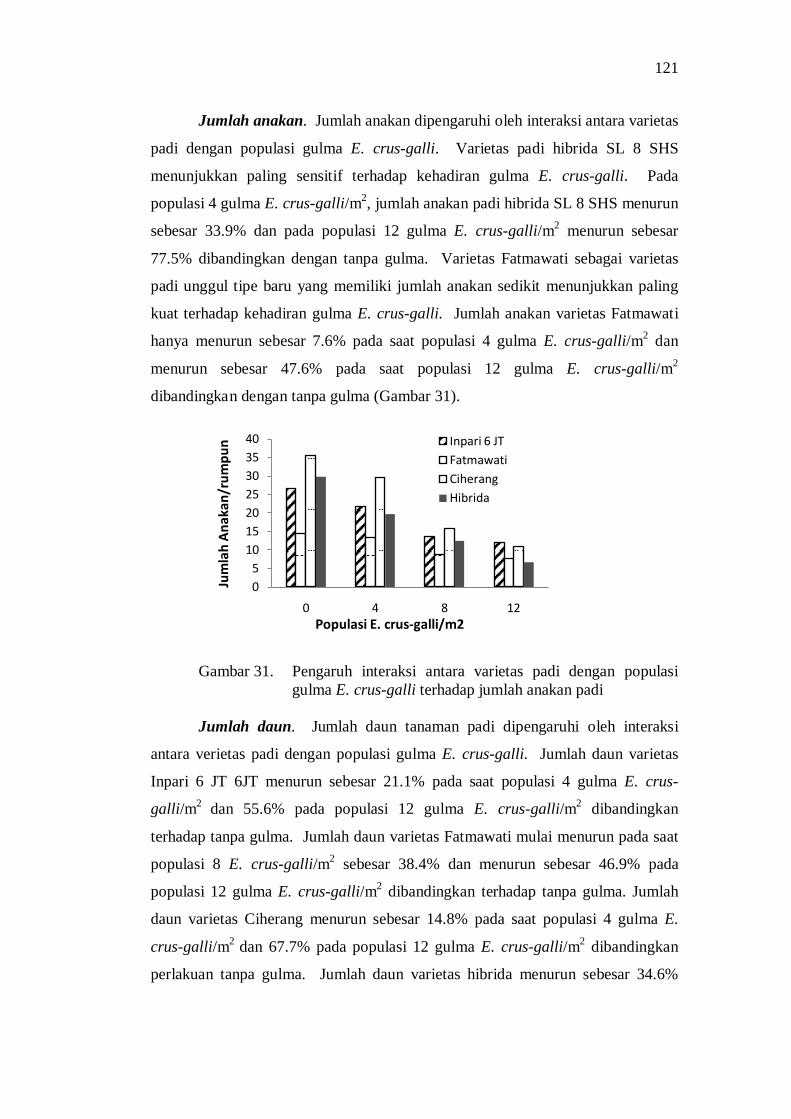
121
Jumlah anakan. Jumlah anakan dipengaruhi oleh interaksi antara varietas
padi dengan populasi gulma E. crus-galli. Varietas padi hibrida SL 8 SHS
menunjukkan paling sensitif terhadap kehadiran gulma E. crus-galli. Pada
populasi 4 gulma E. crus-galli/m2, jumlah anakan padi hibrida SL 8 SHS menurun
sebesar 33.9% dan pada populasi 12 gulma E. crus-galli/m2 menurun sebesar
77.5% dibandingkan dengan tanpa gulma. Varietas Fatmawati sebagai varietas
padi unggul tipe baru yang memiliki jumlah anakan sedikit menunjukkan paling
kuat terhadap kehadiran gulma E. crus-galli. Jumlah anakan varietas Fatmawati
hanya menurun sebesar 7.6% pada saat populasi 4 gulma E. crus-galli/m2 dan
menurun sebesar 47.6% pada saat populasi 12 gulma E. crus-galli/m2
dibandingkan dengan tanpa gulma (Gambar 31).
Gambar 31. Pengaruh interaksi antara varietas padi dengan populasi gulma E. crus-galli terhadap jumlah anakan padi
Jumlah daun. Jumlah daun tanaman padi dipengaruhi oleh interaksi
antara verietas padi dengan populasi gulma E. crus-galli. Jumlah daun varietas
Inpari 6 JT 6JT menurun sebesar 21.1% pada saat populasi 4 gulma E. crus-
galli/m2 dan 55.6% pada populasi 12 gulma E. crus-galli/m2 dibandingkan
terhadap tanpa gulma. Jumlah daun varietas Fatmawati mulai menurun pada saat
populasi 8 E. crus-galli/m2 sebesar 38.4% dan menurun sebesar 46.9% pada
populasi 12 gulma E. crus-galli/m2 dibandingkan terhadap tanpa gulma. Jumlah
daun varietas Ciherang menurun sebesar 14.8% pada saat populasi 4 gulma E.
crus-galli/m2 dan 67.7% pada populasi 12 gulma E. crus-galli/m2 dibandingkan
perlakuan tanpa gulma. Jumlah daun varietas hibrida menurun sebesar 34.6%
05
10152025303540
0 4 8 12
Jum
lah
Ana
kan/
rum
pun
Populasi E. crus-galli/m2
Inpari 6 JTFatmawatiCiherangHibrida
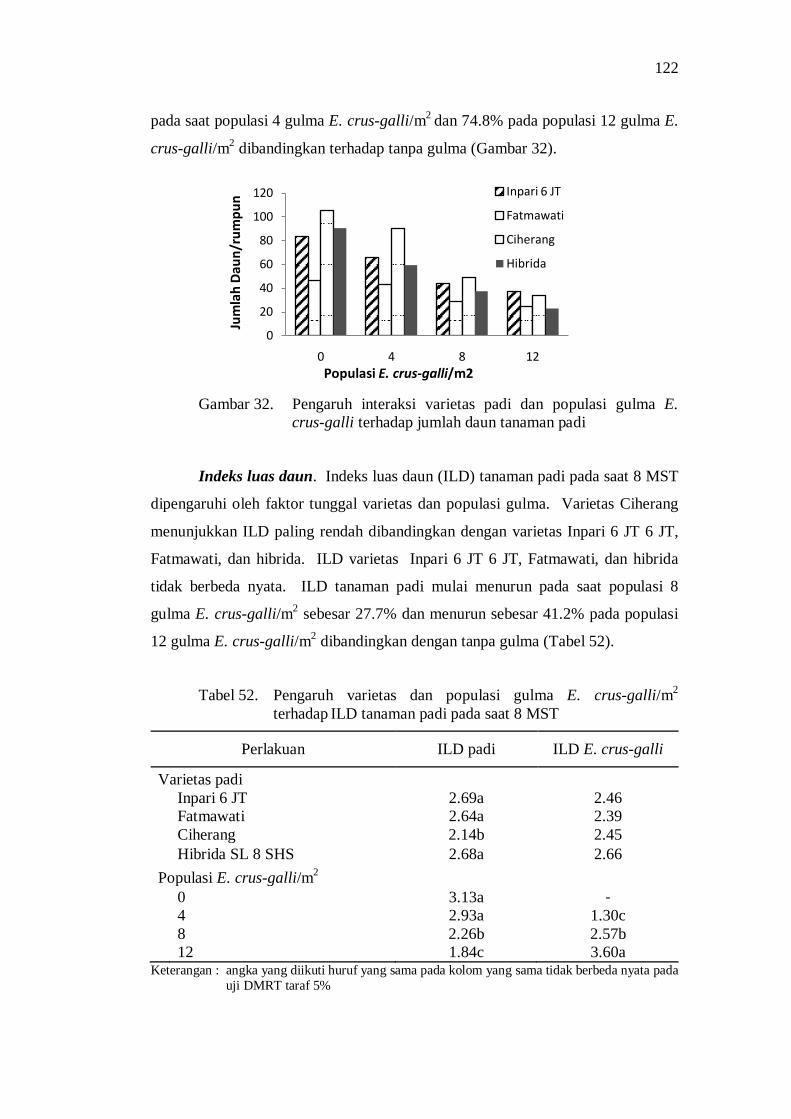
122
pada saat populasi 4 gulma E. crus-galli/m2 dan 74.8% pada populasi 12 gulma E.
crus-galli/m2 dibandingkan terhadap tanpa gulma (Gambar 32).
Gambar 32. Pengaruh interaksi varietas padi dan populasi gulma E. crus-galli terhadap jumlah daun tanaman padi
Indeks luas daun. Indeks luas daun (ILD) tanaman padi pada saat 8 MST
dipengaruhi oleh faktor tunggal varietas dan populasi gulma. Varietas Ciherang
menunjukkan ILD paling rendah dibandingkan dengan varietas Inpari 6 JT 6 JT,
Fatmawati, dan hibrida. ILD varietas Inpari 6 JT 6 JT, Fatmawati, dan hibrida
tidak berbeda nyata. ILD tanaman padi mulai menurun pada saat populasi 8
gulma E. crus-galli/m2 sebesar 27.7% dan menurun sebesar 41.2% pada populasi
12 gulma E. crus-galli/m2 dibandingkan dengan tanpa gulma (Tabel 52).
Tabel 52. Pengaruh varietas dan populasi gulma E. crus-galli/m2
terhadap ILD tanaman padi pada saat 8 MST
Perlakuan ILD padi ILD E. crus-galli
Varietas padi
Inpari 6 JT 2.69a 2.46
Fatmawati 2.64a 2.39
Ciherang 2.14b 2.45
Hibrida SL 8 SHS 2.68a 2.66 Populasi E. crus-galli/m2
0 3.13a -
4 2.93a 1.30c
8 2.26b 2.57b
12 1.84c 3.60a Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada
uji DMRT taraf 5%
0
20
40
60
80
100
120
0 4 8 12
Jum
lah
Dau
n/ru
mpu
n
Populasi E. crus-galli/m2
Inpari 6 JT
Fatmawati
Ciherang
Hibrida
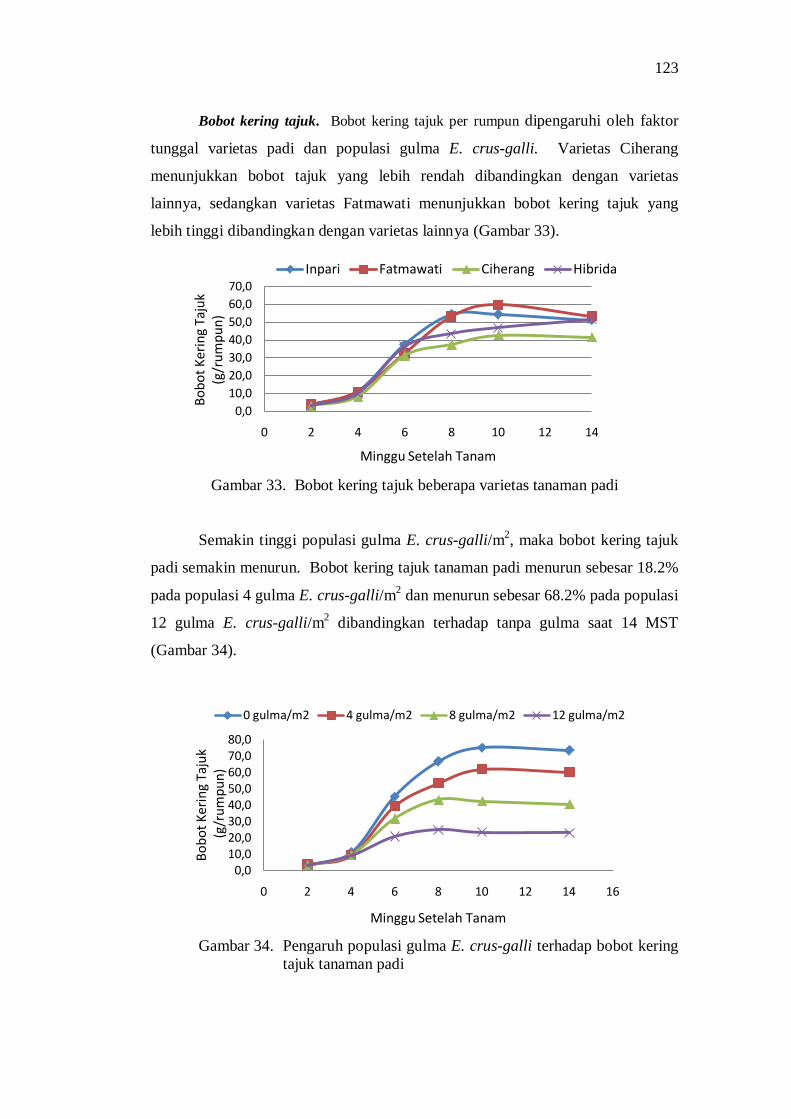
123
Bobot kering tajuk. Bobot kering tajuk per rumpun dipengaruhi oleh faktor
tunggal varietas padi dan populasi gulma E. crus-galli. Varietas Ciherang
menunjukkan bobot tajuk yang lebih rendah dibandingkan dengan varietas
lainnya, sedangkan varietas Fatmawati menunjukkan bobot kering tajuk yang
lebih tinggi dibandingkan dengan varietas lainnya (Gambar 33).
Gambar 33. Bobot kering tajuk beberapa varietas tanaman padi
Semakin tinggi populasi gulma E. crus-galli/m2, maka bobot kering tajuk
padi semakin menurun. Bobot kering tajuk tanaman padi menurun sebesar 18.2%
pada populasi 4 gulma E. crus-galli/m2 dan menurun sebesar 68.2% pada populasi
12 gulma E. crus-galli/m2 dibandingkan terhadap tanpa gulma saat 14 MST
(Gambar 34).
Gambar 34. Pengaruh populasi gulma E. crus-galli terhadap bobot kering
tajuk tanaman padi
0,010,020,030,040,050,060,070,0
0 2 4 6 8 10 12 14
Bobo
t Ker
ing
Taju
k (g
/rum
pun)
Minggu Setelah Tanam
Inpari Fatmawati Ciherang Hibrida
0,010,020,030,040,050,060,070,080,0
0 2 4 6 8 10 12 14 16
Bobo
t Ker
ing
Taju
k(g
/rum
pun)
Minggu Setelah Tanam
0 gulma/m2 4 gulma/m2 8 gulma/m2 12 gulma/m2
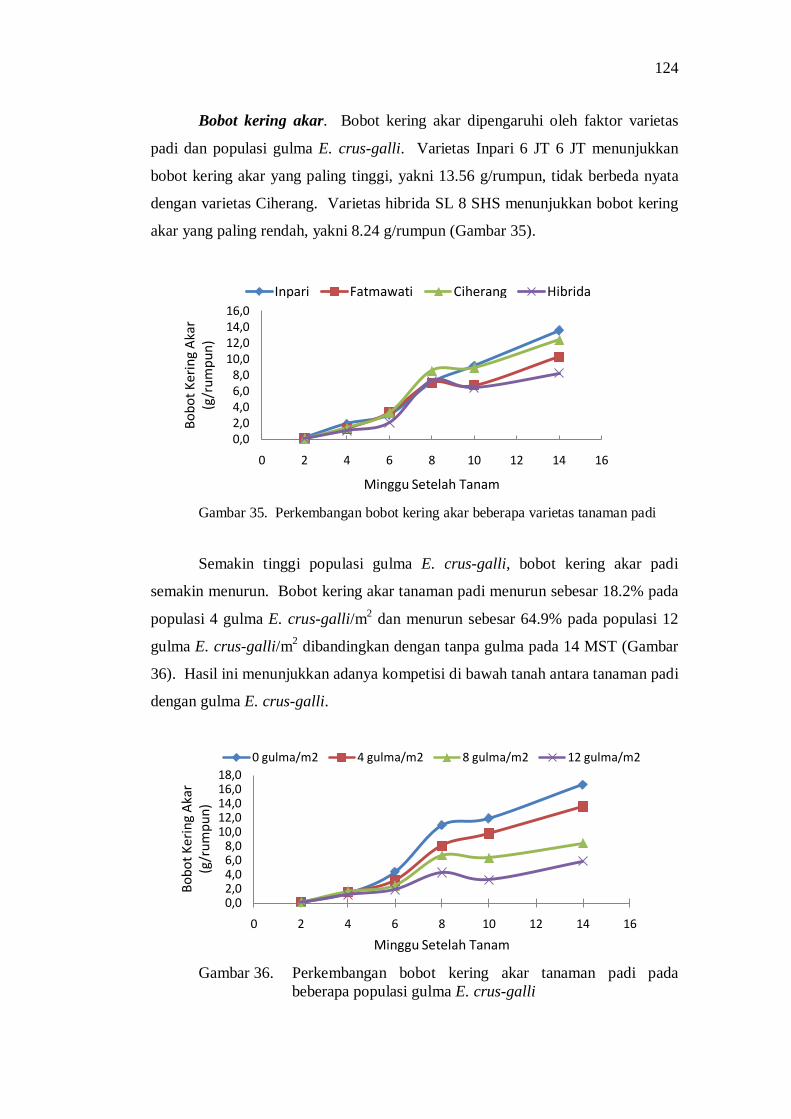
124
Bobot kering akar. Bobot kering akar dipengaruhi oleh faktor varietas
padi dan populasi gulma E. crus-galli. Varietas Inpari 6 JT 6 JT menunjukkan
bobot kering akar yang paling tinggi, yakni 13.56 g/rumpun, tidak berbeda nyata
dengan varietas Ciherang. Varietas hibrida SL 8 SHS menunjukkan bobot kering
akar yang paling rendah, yakni 8.24 g/rumpun (Gambar 35).
Gambar 35. Perkembangan bobot kering akar beberapa varietas tanaman padi
Semakin tinggi populasi gulma E. crus-galli, bobot kering akar padi
semakin menurun. Bobot kering akar tanaman padi menurun sebesar 18.2% pada
populasi 4 gulma E. crus-galli/m2 dan menurun sebesar 64.9% pada populasi 12
gulma E. crus-galli/m2 dibandingkan dengan tanpa gulma pada 14 MST (Gambar
36). Hasil ini menunjukkan adanya kompetisi di bawah tanah antara tanaman padi
dengan gulma E. crus-galli.
Gambar 36. Perkembangan bobot kering akar tanaman padi pada
beberapa populasi gulma E. crus-galli
0,02,04,06,08,0
10,012,014,016,0
0 2 4 6 8 10 12 14 16
Bobo
t Ker
ing
Aka
r (g
/rum
pun)
Minggu Setelah Tanam
Inpari Fatmawati Ciherang Hibrida
0,02,04,06,08,0
10,012,014,016,018,0
0 2 4 6 8 10 12 14 16
Bobo
t Ker
ing
Aka
r(g
/rum
pun)
Minggu Setelah Tanam
0 gulma/m2 4 gulma/m2 8 gulma/m2 12 gulma/m2
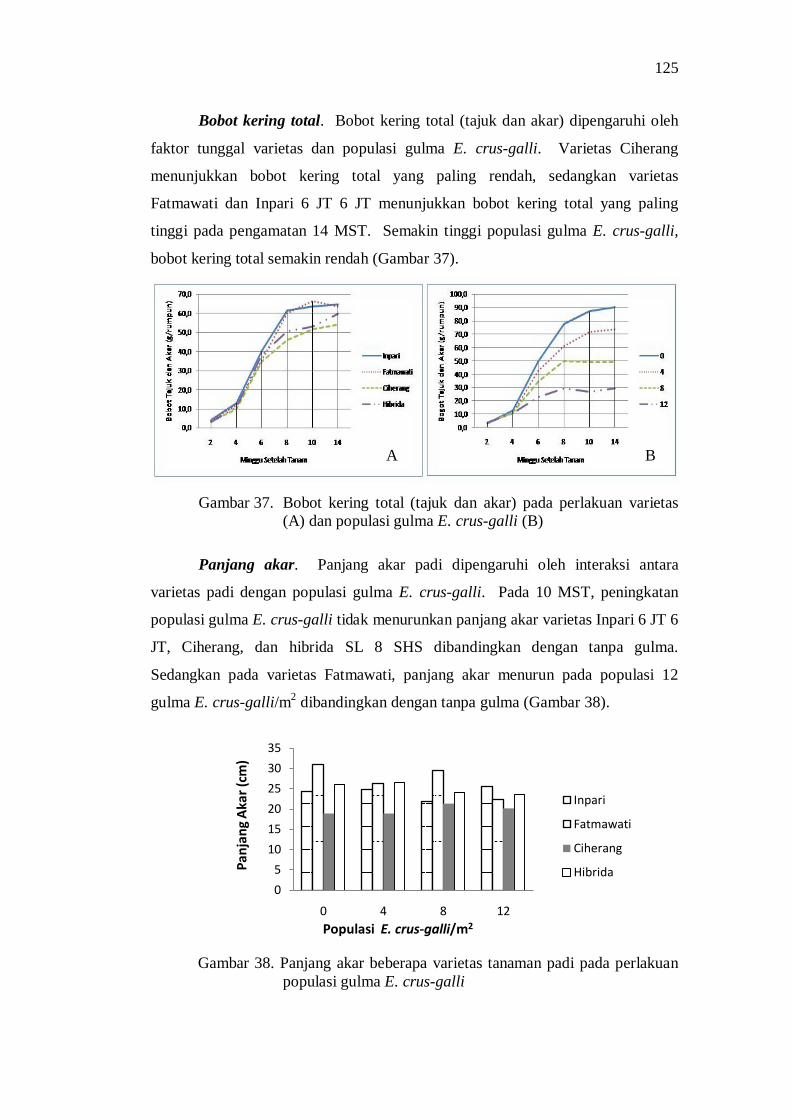
125
Bobot kering total. Bobot kering total (tajuk dan akar) dipengaruhi oleh
faktor tunggal varietas dan populasi gulma E. crus-galli. Varietas Ciherang
menunjukkan bobot kering total yang paling rendah, sedangkan varietas
Fatmawati dan Inpari 6 JT 6 JT menunjukkan bobot kering total yang paling
tinggi pada pengamatan 14 MST. Semakin tinggi populasi gulma E. crus-galli,
bobot kering total semakin rendah (Gambar 37).
Gambar 37. Bobot kering total (tajuk dan akar) pada perlakuan varietas
(A) dan populasi gulma E. crus-galli (B)
Panjang akar. Panjang akar padi dipengaruhi oleh interaksi antara
varietas padi dengan populasi gulma E. crus-galli. Pada 10 MST, peningkatan
populasi gulma E. crus-galli tidak menurunkan panjang akar varietas Inpari 6 JT 6
JT, Ciherang, dan hibrida SL 8 SHS dibandingkan dengan tanpa gulma.
Sedangkan pada varietas Fatmawati, panjang akar menurun pada populasi 12
gulma E. crus-galli/m2 dibandingkan dengan tanpa gulma (Gambar 38).
Gambar 38. Panjang akar beberapa varietas tanaman padi pada perlakuan populasi gulma E. crus-galli
0
5
10
15
20
25
30
35
0 4 8 12
Panj
ang
Aka
r (cm
)
Populasi E. crus-galli/m2
Inpari
Fatmawati
Ciherang
Hibrida
A B
126
Fisiologi Kompetisi antara Tanaman Padi dengan Gulma E. crus-galli
Kandungan hara daun. Kandungan hara P, K, dan Mg pada tajuk
tanaman padi hanya dipengaruhi oleh varietas padi. Varietas Fatmawati
menunjukkan kandungan hara P tajuk paling tinggi, yakni 3.07%, tetapi
kandungan K terendah, yakni 2.09%. Varietas Inpari 6 JT menunjukkan
kandungan hara P tajuk yang terendah, yakni 0.32%, dan kandungan hara K tajuk
tertinggi, yakni 2.42%. Varietas Ciherang menunjukkan serapan hara Mg yang
lebih tinggi dibandingkan dengan varietas lainnya, yakni 0.18% (Tabel 53).
Tabel 53. Kandungan hara daun beberapa varietas tanaman padi pada populasi gulma E. crus-galli yang berbeda
Perlakuan Kandungan Hara pada Daun Padi N P K Ca Mg
Varietas padi ------------------------ % ------------------------- Inpari 6 JT 3.07 0.32c 2.42a 0.27 0.14b Fatmawati 2.86 0.37a 2.09b 0.34 0.14b Ciherang 3.16 0.36ab 2.21ab 0.26 0.18a Hibrida SL 8 SHS 3.13 0.34bc 2.16ab 0.25 0.13b Populasi E. crus-galli/m2
0 3.10 0.36 2.28 0.26 0.14 4 3.09 0.35 2.27 0.29 0.16 8 3.06 0.35 2.15 0.29 0.16 12 2.97 0.34 2.17 0.28 0.15
Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda nyata pada uji DMRT taraf 5%
Efisiensi serapan hara. Efisiensi serapan hara menunjukkan banyaknya
bahan kering yang dapat diproduksi oleh setiap satuan hara yang diserap oleh
tanaman. Efisiensi serapan hara N, K, dan Ca tidak dipengaruhi oleh faktor
varietas, populasi gulma E. crus-galli maupun interaksi antara varietas dan
populasi gulma. Efisiensi serapan hara P dan Mg hanya dipengaruhi oleh faktor
tunggal varietas tanaman padi (Tabel 54).
Varietas Inpari 6 JT menunjukkan efisiensi serapan hara P yang lebih
tinggi dibandingkan dengan varietas Fatmawati dan Ciherang, namun tidak
berbeda nyata dengan varietas hibrida SL 8 SHS. Efisiensi serapan hara Mg
tertinggi ditunjukkan oleh varietas hibrida SL 8 SHS dan efisiensi serapan hara
terendah ditunjukkan oleh varietas Ciherang (Tabel 54).
127
Tabel 54. Efisiensi serapan hara pada beberapa varietas tanaman padi dan populasi gulma E. crus-galli
Perlakuan N P K Ca Mg
Varietas -----------------g biomass / g hara------------------
Inpari 6 JT 0.33 3.08a 0.42 4.06 7.01ab
Fatmawati 0.37 2.67b 0.51 3.40 7.03ab
Ciherang 0.32 2.73b 0.45 4.09 5.66b
Hibrida SL 8 SHS 0.33 2.90ab 0.47 4.16 8.24a Populasi E. crus-galli/m2
0 0.34 2.75 0.45 4.47 8.31
4 0.33 2.84 0.45 3.46 6.39
8 0.34 2.87 0.49 3.81 6.42
12 0.34 2.93 0.46 3.96 6.81 Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda nyata pada
uji DMRT taraf 5%.
Leaf Area Ratio. Leaf area ratio (LAR) merupakan perbandingan antara
luas daun dengan bobot kering tanaman total. LAR antar varietas tidak berbeda
nyata, namun terlihat bahwa varietas Fatmawati menunjukkan nilai LAR yang
cenderung lebih rendah dibandingkan dengan varietas lainnya, yakni sebesar
24.26 cm2/g (Tabel 55). Hal ini berarti bahwa varietas Fatmawati membutuhkan
luasan daun yang cenderung lebih rendah untuk memproduksi setiap satuan bahan
kering atau cenderung lebih efisien dibandingkan dengan varietas lainnya.
LAR dipengaruhi oleh tingkat populasi gulma E. crus-galli. Perlakuan
populasi 4 dan 8 gulma E. crus-galli/m2 menunjukkan LAR yang cenderung
meningkat dibandingkan dengan tanpa gulma, sedangkan populasi 12 E. crus-
galli/m2 menunjukkan nilai LAR yang nyata lebih tinggi atau meningkat sebesar
69.01% dibandingkan dengan tanpa gulma (Tabel 55). Hasil ini menunjukkan
bahwa semakin tinggi populasi gulma menyebabkan produksi bahan kering
semakin tidak efisien. Produksi setiap satuan bahan kering membutuhkan luasan
daun padi yang lebih luas dengan semakin meningkatnya populasi gulma.
Net Assimilation Rate. Net assimilation rate (NAR) atau laju asimilasi
bersih hanya dipengaruhi oleh populasi gulma E. crus-galli. Nilai NAR mulai
nyata menurun pada saat populasi 8 gulma E. crus-galli/m2 yakni menurun
sebesar 29.9% dibandingkan dengan tanpa gulma dan NAR menurun lebih besar
lagi pada populasi 12 gulma E. crus-galli/m2 yakni sebesar 61.6% dibandingkan
dengan tanpa gulma (Tabel 55). Hasil ini menunjukkan bahwa kompetisi gulma
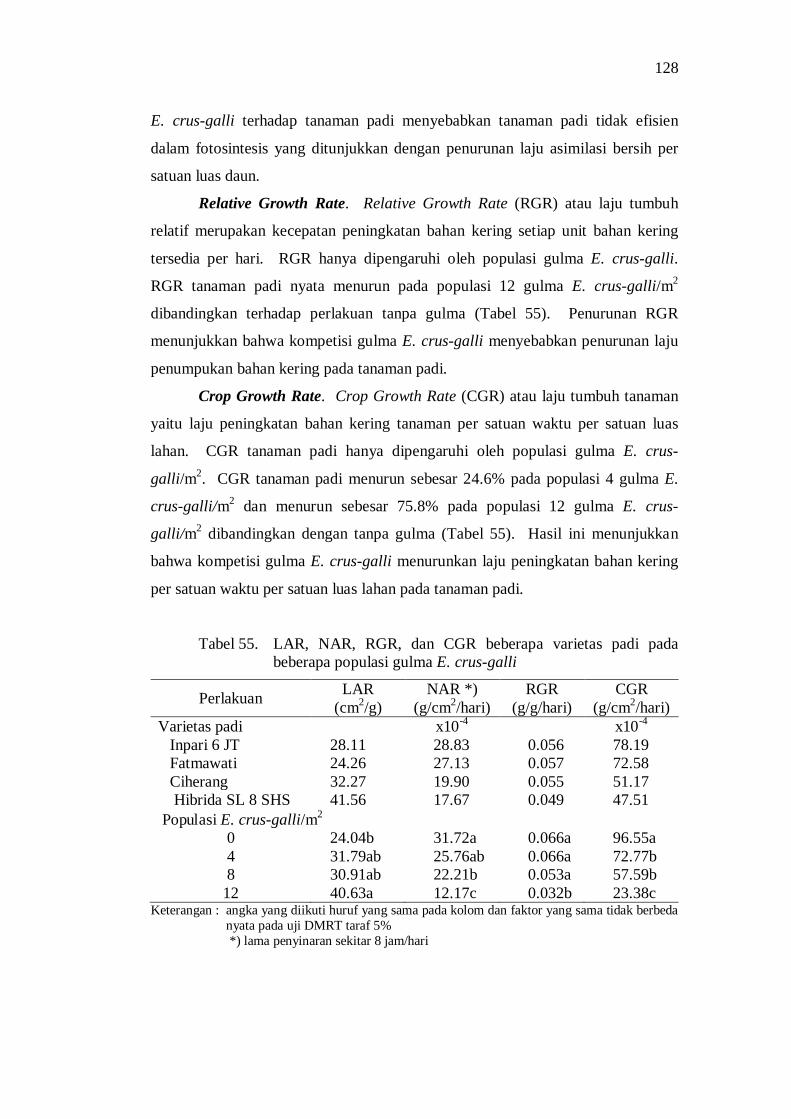
128
E. crus-galli terhadap tanaman padi menyebabkan tanaman padi tidak efisien
dalam fotosintesis yang ditunjukkan dengan penurunan laju asimilasi bersih per
satuan luas daun.
Relative Growth Rate. Relative Growth Rate (RGR) atau laju tumbuh
relatif merupakan kecepatan peningkatan bahan kering setiap unit bahan kering
tersedia per hari. RGR hanya dipengaruhi oleh populasi gulma E. crus-galli.
RGR tanaman padi nyata menurun pada populasi 12 gulma E. crus-galli/m2
dibandingkan terhadap perlakuan tanpa gulma (Tabel 55). Penurunan RGR
menunjukkan bahwa kompetisi gulma E. crus-galli menyebabkan penurunan laju
penumpukan bahan kering pada tanaman padi.
Crop Growth Rate. Crop Growth Rate (CGR) atau laju tumbuh tanaman
yaitu laju peningkatan bahan kering tanaman per satuan waktu per satuan luas
lahan. CGR tanaman padi hanya dipengaruhi oleh populasi gulma E. crus-
galli/m2. CGR tanaman padi menurun sebesar 24.6% pada populasi 4 gulma E.
crus-galli/m2 dan menurun sebesar 75.8% pada populasi 12 gulma E. crus-
galli/m2 dibandingkan dengan tanpa gulma (Tabel 55). Hasil ini menunjukkan
bahwa kompetisi gulma E. crus-galli menurunkan laju peningkatan bahan kering
per satuan waktu per satuan luas lahan pada tanaman padi.
Tabel 55. LAR, NAR, RGR, dan CGR beberapa varietas padi pada beberapa populasi gulma E. crus-galli
Perlakuan LAR (cm2/g)
NAR *) (g/cm2/hari)
RGR (g/g/hari)
CGR (g/cm2/hari)
Varietas padi
x10-4
x10-4 Inpari 6 JT 28.11 28.83 0.056 78.19 Fatmawati 24.26 27.13 0.057 72.58 Ciherang 32.27 19.90 0.055 51.17 Hibrida SL 8 SHS 41.56 17.67 0.049 47.51 Populasi E. crus-galli/m2
0 24.04b 31.72a 0.066a 96.55a 4 31.79ab 25.76ab 0.066a 72.77b 8 30.91ab 22.21b 0.053a 57.59b
12 40.63a 12.17c 0.032b 23.38c Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda
nyata pada uji DMRT taraf 5% *) lama penyinaran sekitar 8 jam/hari

129
Pertumbuhan Generatif Tanaman Padi
Waktu heading. Waktu heading tanaman padi hanya dipengaruhi oleh
varietas padi. Varietas Inpari 6 JT dan hibrida SL 8 SHS memiliki waktu
heading yang lebih panjang daripada varietas Fatmawati dan Ciherang. Varietas
Fatmawati memiliki waktu heading yang paling cepat, yaitu sekitar 62.9 hari
setelah tanam (Tabel 56).
Tabel 56. Waktu heading tanaman padi pada perlakuan varietas padi dan populasi gulma E. crus-galli
Perlakuan Waktu Heading (HST) Varietas padi
Inpari 6 JT 77.6a
Fatmawati 62.9c
Ciherang 68.9b
Hibrida SL 8 SHS 77.2a Populasi E. crus-galli/m2
0 71.2
4 71.4
8 72.0
12 72.0 Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda nyata
pada uji DMRT taraf 5%
Komponen Produksi Tanaman Padi
Jumlan anakan produktif. Jumlah anakan produktif dipengaruhi oleh
interaksi antara varietas padi dengan populasi gulma E. crus-galli. Varietas Inpari
6 JT memiliki jumlah anakan tertinggi pada kondisi tanpa gulma. Jumlah anakan
menurun pada populasi 4 dan 12 gulma/m2 berturut-turut sebesar 10.2% dan
65.6% dibandingkan terhadap tanpa gulma. Varietas Fatmawati pada kondisi
tanpa gulma memiliki jumlah anakan produktif yang lebih rendah dibandingkan
dengan varietas lainnya. Jumlah anakan produktif varietas Fatmawati menurun
pada populasi 4 hingga 12 gulma/m2 berturut-turut sebesar 22.2% hingga 53.1%.
Jumlah anakan produktif varietas Ciherang menurun pada populasi 4 hingga 12
gulma/m2 bertutur-turut sebesar 42.1% hingga 88.7%. Jumlah anakan produktif
varietas hibrida SL 8 SHS menurun sebesar 23.4% pada populasi 4 gulma/m2 dan
menurun sebesar 70.2% pada populasi 12 gulma/m2 (Gambar 39). Hasil tersebut
menunjukkan bahwa varietas Ciherang lebih sensitif terhadap kehadiran gulma E.
crus-galli dibandingkan dengan varietas lainnya. Varietas Fatmawati

130
menunjukkan kemampuan bersaing dengan gulma E. crus-galli yang lebih kuat
yang ditunjukkan dengan penurunan jumlah anakan produktif pada populasi 12
gulma/m2 yang lebih rendah dibandingkan dengan varietas lainnya.
Gambar 39. Pengaruh populasi gulma E. crus-galli terhadap jumlah
anakan produktif beberapa varietas tanaman padi
Panjang malai dan kepadatan malai. Panjang malai dan kepadatan malai
padi dipengaruhi oleh interaksi antara varietas dan populasi gulma E. crus-galli.
Panjang malai varietas Inpari 6 JT dan Ciherang mulai menurun pada populasi 12
gulma E. crus-galli/m2. Panjang malai varietas Fatmawati dan hibrida tidak
berbeda nyata pada semua perlakuan populasi gulma E. crus-galli (Tabel 57).
Tabel 57. Pengaruh interaksi varietas padi dan populasi gulma E. crus-galli terhadap panjang malai dan jumlah biji per malai
Varietas Populasi Gulma E. crus-galli/m2 0 4 8 12
Panjang Malai (cm) Inpari 6 JT 26.5b 25.7bcd 26.3bc 24.5d Fatmawati 29.3a 29.9a 29.6a 29.6a Ciherang 25.4bcd 24.8bcd 24.8bcd 22.2e Hibrida 25.7bcd 25.7bcd 24.5cd 25.0bcd Jumlah Biji per Malai (bulir) Inpari 6 JT 169.3bc 147.9cdef 159.5bcd 130.2fg Fatmawati 244.1a 236.0a 232.7a 242.8a Ciherang 149.9cdef 153.8bcde 122.0g 134.1efg Hibrida 161.8bc 175.9b 156.5bcde 138.6defg
Keterangan : angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%
Bobot 1000 butir gabah. Bobot 1000 butir gabah dipengaruhi oleh
varietas tanaman padi, tetapi tidak dipengaruhi oleh populasi gulma E. crus-galli
0,0
5,0
10,0
15,0
20,0
0 4 8 12
Jum
lah
Ana
kan
Prod
ukti
f
Populasi E. crus-galli/m²
InpariFatmawatiCiherangHibrida

131
maupun interaksi antara varietas dan populasi gulma. Varietas Fatmawati
menunjukkan bobot gabah 1000 butir tertinggi dibandingkan dengan varietas
lainnya, yakni 26.10 g, sedangkan varietas padi Ciherang menunjukkan bobot
1000 butir yang terendah, yakni 22.52 g (Tabel 58).
Tabel 58. Bobot 1 000 butir gabah pada beberapa varietas tanaman padi dan pada berbagai populasi gulma E. crus-galli
Perlakuan Bobot 1 000 butir (g) Varietas padi
Inpari 6 JT 25.49b
Fatmawati 26.10a
Ciherang 22.52c
Hibrida SL 8 SHS 25.15b Populasi E. crus-galli/m2
0 24.90
4 25.05
8 24.81
12 24.49 Keterangan : angka yang diikuti huruf yang sama pada kolom dan faktor yang sama tidak berbeda
nyata pada uji DMRT taraf 5%
Hasil Produksi Tanaman Padi
Produksi gabah. Setiap varietas padi menunjukkan penurunan produksi
dengan semakin meningkatnya populasi gulma E. crus-galli. Varietas Ciherang
menunjukkan produksi GKP dan GKG ubinan paling rendah, sedangkan varietas
Inpari 6 JT menunjukkan produksi GKP dan GKG tertinggi dibandingkan dengan
varietas lainnya. Penurunan produksi GKP per hektar varietas Ciherang,
Fatmawati, hibrida SL 8 SHS, dan Inpari 6 JT berturut-turut mengikuti persamaan
garis linear : y(ciherang) = -0.275x + 4.704, y(Fatmawati) = -0.202x + 6.405, y(hibrida) = -
0.476x + 8.280, dan y(Inpari 6 JT 6 JT) = -0.424x + 9.517, sedangkan produksi GKG
per hektar mengikuti persamaan garis linear : y(Ciherang) = -0.232x + 3.965,
y(Fatmawati) = -0.211x + 5.515, y(Hibrida) = -0.405x + 7.030, dan y(Inpari 6 JT 6JT) = -
0.358x + 8.030. Berdasarkan persamaan tersebut, terlihat bahwa varietas padi
hibrida lebih peka terhadap tingkat populasi gulma E. crus-galli, sedangkan
varietas Fatmawati lebih kuat dibandingkan dengan varietas lainnya.
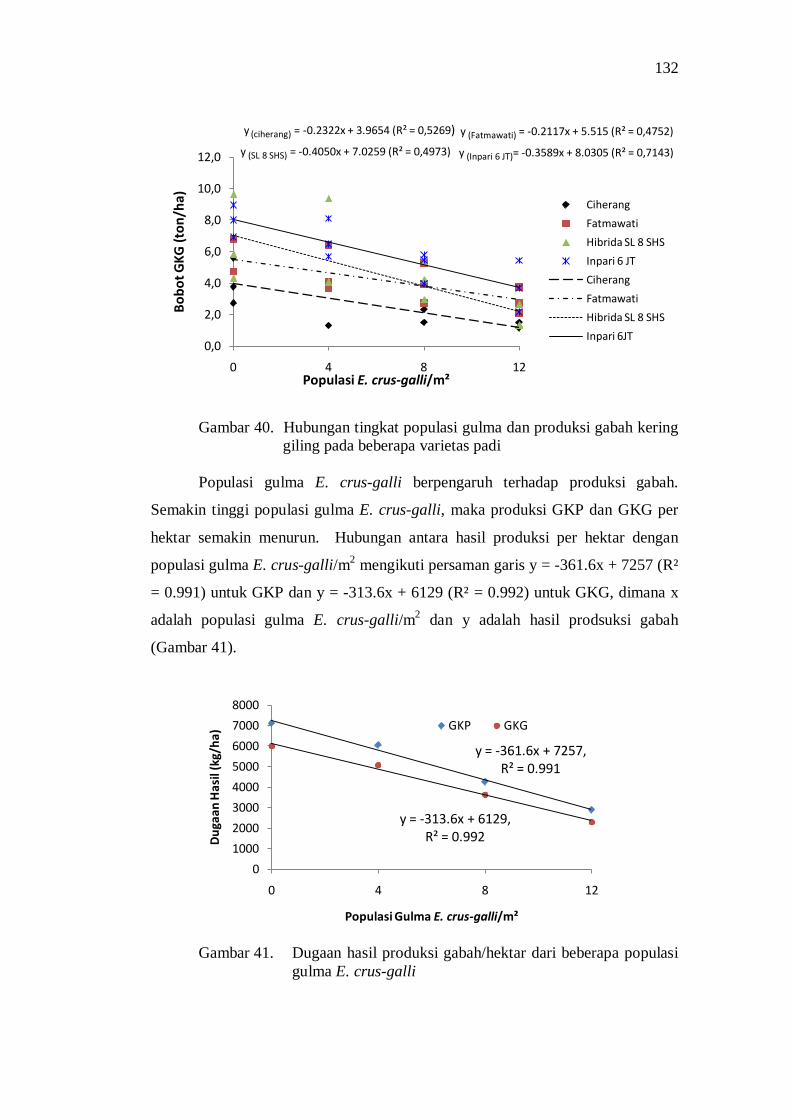
132
Gambar 40. Hubungan tingkat populasi gulma dan produksi gabah kering giling pada beberapa varietas padi
Populasi gulma E. crus-galli berpengaruh terhadap produksi gabah.
Semakin tinggi populasi gulma E. crus-galli, maka produksi GKP dan GKG per
hektar semakin menurun. Hubungan antara hasil produksi per hektar dengan
populasi gulma E. crus-galli/m2 mengikuti persaman garis y = -361.6x + 7257 (R²
= 0.991) untuk GKP dan y = -313.6x + 6129 (R² = 0.992) untuk GKG, dimana x
adalah populasi gulma E. crus-galli/m2 dan y adalah hasil prodsuksi gabah
(Gambar 41).
Gambar 41. Dugaan hasil produksi gabah/hektar dari beberapa populasi gulma E. crus-galli
y (ciherang) = -0.2322x + 3.9654 (R² = 0,5269) y (Fatmawati) = -0.2117x + 5.515 (R² = 0,4752)
y (SL 8 SHS) = -0.4050x + 7.0259 (R² = 0,4973) y (Inpari 6 JT)= -0.3589x + 8.0305 (R² = 0,7143)
0,0
2,0
4,0
6,0
8,0
10,0
12,0
0 4 8 12
Bobo
t GKG
(ton
/ha)
Populasi E. crus-galli/m²
Ciherang
Fatmawati
Hibrida SL 8 SHS
Inpari 6 JT
Ciherang
Fatmawati
Hibrida SL 8 SHS
Inpari 6JT
y = -361.6x + 7257,R² = 0.991
y = -313.6x + 6129,R² = 0.992
0
1000
2000
3000
4000
5000
6000
7000
8000
0 4 8 12
Dug
aan
Has
il (k
g/ha
)
Populasi Gulma E. crus-galli/m²
GKP GKG
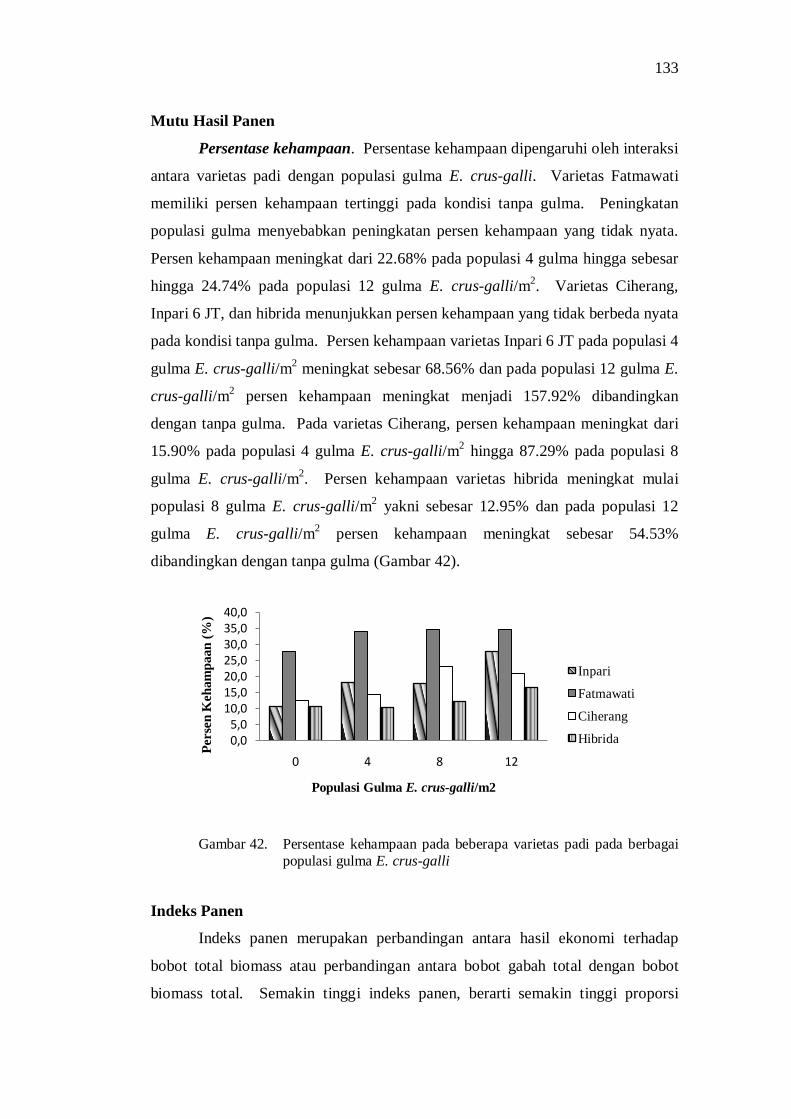
133
Mutu Hasil Panen
Persentase kehampaan. Persentase kehampaan dipengaruhi oleh interaksi
antara varietas padi dengan populasi gulma E. crus-galli. Varietas Fatmawati
memiliki persen kehampaan tertinggi pada kondisi tanpa gulma. Peningkatan
populasi gulma menyebabkan peningkatan persen kehampaan yang tidak nyata.
Persen kehampaan meningkat dari 22.68% pada populasi 4 gulma hingga sebesar
hingga 24.74% pada populasi 12 gulma E. crus-galli/m2. Varietas Ciherang,
Inpari 6 JT, dan hibrida menunjukkan persen kehampaan yang tidak berbeda nyata
pada kondisi tanpa gulma. Persen kehampaan varietas Inpari 6 JT pada populasi 4
gulma E. crus-galli/m2 meningkat sebesar 68.56% dan pada populasi 12 gulma E.
crus-galli/m2 persen kehampaan meningkat menjadi 157.92% dibandingkan
dengan tanpa gulma. Pada varietas Ciherang, persen kehampaan meningkat dari
15.90% pada populasi 4 gulma E. crus-galli/m2 hingga 87.29% pada populasi 8
gulma E. crus-galli/m2. Persen kehampaan varietas hibrida meningkat mulai
populasi 8 gulma E. crus-galli/m2 yakni sebesar 12.95% dan pada populasi 12
gulma E. crus-galli/m2 persen kehampaan meningkat sebesar 54.53%
dibandingkan dengan tanpa gulma (Gambar 42).
Gambar 42. Persentase kehampaan pada beberapa varietas padi pada berbagai populasi gulma E. crus-galli
Indeks Panen
Indeks panen merupakan perbandingan antara hasil ekonomi terhadap
bobot total biomass atau perbandingan antara bobot gabah total dengan bobot
biomass total. Semakin tinggi indeks panen, berarti semakin tinggi proporsi
0,05,0
10,015,020,025,030,035,040,0
0 4 8 12
Pers
en K
eham
paan
(%)
Populasi Gulma E. crus-galli/m2
Inpari
Fatmawati
Ciherang
Hibrida

134
bahan kering hasil fotosintesis yang dialokasikan untuk pembentukan gabah.
Indeks panen dipengaruhi oleh varietas dan populasi gulma. Varietas Inpari 6 JT
dan hibrida menunjukkan indeks panen yang lebih tinggi dibandingkan dengan
varietas Fatmawati dan Ciherang. Varietas Ciherang menunjukkan indeks panen
yang cenderung lebih rendah dibandingkan dengan varietas Fatmawati (Tabel 60).
Indeks panen dipengaruhi oleh tingkat populasi gulma. Indeks panen
menurun pada saat populasi 12 gulma E. crus-galli, yakni menurun sebesar 20%
dibandingkan dengan tanpa gulma (Tabel 60). Hasil ini menunjukkan bahwa
gulma bersaing dengan tanaman padi, secara tidak langsung mengurangi proporsi
fotosintat yang dialokasikan ke pengisian gabah.
Tabel 59. Pengaruh tingkat populasi gulma E. crus-galli terhadap indeks panen pada beberapa varietas tanaman padi
Perlakuan Indeks Panen
Varietas Inpari 6 JT 0.69a
Fatmawati 0.41b Ciherang 0.33b Hibrida SL 8 SHS 0.58a
Populasi E. crus-galli/m2 0 0.55a
4 0.49ab 8 0.54a 12 0.44b
Pembahasan
Kompetisi antara tanaman padi dengan gulma E. crus-galli dipengaruhi
oleh interaksi antara varietas padi dengan populasi gulma E. crus-galli. Interaksi
tersebut berpengaruh terhadap peubah tinggi tanaman, jumlah anakan, jumlah
daun, panjang akar, jumlah anakan produktif, panjang malai, kepadatan malai, dan
kehampaan. Tinggi tanaman, jumlah anakan total, dan jumlah daun sangat
menentukan kompetisi di atas tanah, sedangkan panjang akar akan menentukan
kemampuan bersaing di bawah permukaan tanah.
Jumlah anakan produktif, panjang malai, kepadatan malai, dan persen
kehampaan merupakan komponen produksi tanaman padi. Varietas tanaman padi

135
yang sensitif terhadap kehadiran gulma pada peubah tersebut akan menunjukkan
penurunan produksi yang lebih besar.
Varietas Fatmawati menunjukkan tinggi tanaman, jumlah anakan, jumlah
daun yang paling tinggi dibandingkan dengan varietas lainnya pada tingkat
populasi gulma E. crus-galli yang sama. Varietas Fatmawati mengalami
penurunan jumlah anakan produktif yang lebih rendah dibandingkan dengan
varietas lainnya pada tingkat populasi gulma yang sama, sedangkan panjang
malai, kepadatan malai, dan persentase hampa tidak dipengaruhi oleh populasi
gulma. Berdasarkan peubah komponen hasil ini, varietas Fatmawati merupakan
varietas yang memiliki kemampuan kompetisi yang kuat terhadap gulma E. crus-
galli. Berdasarkan hubungan antara tingkat populasi gulma dengan produksi
gabah kering giling (Gambar 40), terlihat bahwa penurunan produksi varietas
Fatmawati lebih rendah dibandingkan dengan varietas lainnya. Namun, varietas
Fatmawati ini memiliki persentase kehampaan yang tiga kali lebih besar
dibandingkan dengan varietas lainnya (Gambar 42), sehingga produksi produksi
GKG lebih rendah dibandingkan varietas Inpari 6 JT.
Varietas hibrida SL 8 SHS menunjukkan tinggi tanaman yang paling
rendah, penurunan jumlah anakan dan jumlah daun yang paling tinggi
dibandingkan dengan varietas lainnya ketika populasi gulma E. crus-galli semakin
meningkat (Gambar 30, 31, dan 32). Berdasarkan hubungan antara tingkat
populasi gulma dengan produksi gabah kering giling (Gambar 40), terlihat bahwa
penurunan produksi varietas hibrida SL 8 SHS lebih besar dibandingkan dengan
varietas lainnya.
Produksi varietas Ciherang baik dalam bentuk GKP maupun GKG
menunjukkan produksi yang paling rendah dibandingkan dengan varietas lainnya,
baik dalam kondisi ada gulma maupun tidak ada gulma E. crus-galli. Rendahnya
produksi varietas Ciherang ini antara lain disebabkan oleh rendahnya komponen
produksi. Jumlah anakan produktif varietas Ciherang menurunkan sebesar 42.2%
pada populasi 4 gulma/m2 dan menurun sebesar 88.7% pada populasi 12 gulma E.
crus-galli/m2. Varietas Ciherang mengalami penurunan panjang malai ketika
populasi 12 E. crus-galli/m2, sedangkan varietas yang lainnya tidak mengalami
penurunan panjang malai pada berbagai populasi gulma. Varietas Ciherang juga

136
mengalami penurunan kepadatan malai mulai populasi 8 gulma E. crus-galli/m2,
sedangkan Inpari 6 JT dan hibrida SL 8 SHS mengalami penurunan kepadatan
malai pada saat populasi 12 gulma E. crus-galli/m2.
Kompetisi antara tanaman padi dan gulma E. crus-galli dipengaruhi oleh
faktor tingkat populasi gulma E. crus-galli. Kehadiran gulma E. crus-galli
menyebabkan penurunan ILD, bobot kering tajuk, bobot kering akar, bobot kering
total, net assimilation rate (NAR), relative growth rate (RGR) dan crop growth
rate (CGR), dan menyebabkan peningkatan leaf area ratio (LAR), serta
menurunkan produksi gabah, baik GKP maupun GKG.
Produksi berat kering biomass dipengaruhi oleh indeks luas daun (Weng et
al. 1982). Hasil penelitian menunjukkan bahwa varietas Ciherang dengan ILD
yang paling rendah menunjukkan berat kering biomass total yang paling rendah.
Sedangkan, varietas Fatmawati dan Inpari 6 JT memiliki ILD yang tinggi
menunjukkan bobot kering biomass total yang paling tinggi. ILD yang tinggi
menentukan kapasitas fotosintesis yang lebih baik. Hal ini ditunjukkan oleh leaf
area rasio varietas Fatmawati yang lebih rendah atau lebih efisien dalam produksi
bahan kering. Menurut Mia et al. (2011), ILD yang tinggi menyebabkan kapasitas
fotosintesis lebih baik, sehingga produksi bahan kering meningkat. Chen et al.
(1991) menyatakan bahwa produksi bahan kering berkorelasi positif terhadap
hasil gabah.
Harvest indeks (HI) berkorelasi dengan produksi gabah (Cui-Jing et al.
2000). Varietas Fatmawati dan Ciherang memiliki HI yang lebih rendah
dibandingkan dengan varietas Inpari 6 JT dan hibrida. Hal ini berarti bahwa
translokasi hasil asilmilat ke pembentukan gabah pada varietas Fatmawati dan
Ciherang lebih rendah. Pada varietas Fatmawati, rendahnya translokasi asimilat
dari source ke sink yang rendah menyebabkan persen kehampaan meningkat
sehingga produksi gabah menurun, meskipun bobot 1000 butir tinggi. Pada
varietas Ciherang, rendahnya translokasi hasil asimilat ke gabah menyebabkan
bobot 1000 butir rendah, sehingga produksi juga rendah.
Penurunan produksi GKP per hektar varietas Ciherang, Fatmawati,
Hibrida, dan Inpari 6 JT berturut-turut mengikuti persamaan garis linear : y(ciherang)
= -0.275x + 4.704, y(Fatmawati) = -0.202x + 6.405, y(hibrida) = -0.476x + 8.280, dan

137
y(Inpari 6 JT) = -0.424x + 9.517, sedangkan produksi GKG per hektar mengikuti
persamaan garis linear : y(Ciherang) = -0.232x + 3.965, y(Fatmawati) = -0.211x + 5.515,
y(Hibrida) = -0.405x + 7.030, dan y(Inpari 6 JT) = -0.358x + 8.030. Berdasarkan
persamaan tersebut, terlihat bahwa varietas padi hibrida lebih peka terhadap
tingkat populasi gulma E. crus-galli yang ditunjukkan dengan slope garis dugaan
yang lebih besar dibandingkan dengan varietas lainnya, sedangkan varietas
Fatmawati lebih kuat dibandingkan dengan varietas lainnya yang ditunjukkan
dengan slope garis dugaan yang lebih kecil (Gambar 40).
Jika penurunan produksi GKP yang ditolerir adalah 10% terhadap hasil
maksimum yang dapat dicapai oleh masing-masing varietas, maka ambang
ekonomi varietas Fatmawati terjadi pada saat populasi 3.2 gulma E. crus-galli/m2,
sedangkan pada varietas hibrida terjadi pada saat populasi 1.7 gulma E. crus-
galli/m2. Konsekuensinya adalah bahwa penyiangan pada tanaman padi hibrida
harus segera dilakukan pada saat populasi gulma sudah mencapai 1.7 gulma E.
crus-galli/m2 untuk menyelamatkan 10% hasil produksi.
Hubungan antara tingkat populasi gulma E. crus-galli secara umum pada
seluruh varietas dan tingkat produksi tanaman padi mengikuti persamaan garis
linear yaitu y = -361.6x + 7257 (R² = 0.991) untuk produksi GKP (kg/ha) dan y =
-313.6x + 6129 (R² = 0.992) untuk produksi GKG (kg/ha), dimana x adalah
populasi gulma E. crus-galli/m2 dan y adalah hasil prodsuksi gabah. Berdasarkan
persamaan garis ini penurunan produksi GKP 10% terjadi pada populasi 2 gulma
E. crus-galli/m2 dan penurunan produksi GKG 10% terjadi pada populasi 1.9
gulma E. crus-galli/m2.
Kesimpulan
Kompetisi antara tanaman padi dan gulma E. crus-galli dipengaruhi oleh
varietas tanaman padi dan tingkat populasi gulma E. crus-galli. Kompetisi
menyebabkan pertumbuhan dan produksi tanaman padi menurun.
Setiap varietas memiliki kepekaan ataupun kekuatan kompetisi yang
berbeda terhadap tingkat populasi gulma E. crus-galli. Varietas Fatmawati
menunjukkan kemampuan kompetisi yang lebih kuat terhadap gulma E. crus-galli

138
dibandingkan dengan varietas lainnya, sedangkan varietas hibrida SL 8 SHS
menunjukkan varietas yang paling sensitif terhadap populasi gulma E. crus-galli.
Tingkat populasi gulma E. crus-galli berpengaruh terhadap pertumbuhan
dan produksi tanaman padi. Semakin tinggi tingkat populasi gulma E. crus-galli,
maka semakin tinggi tingkat penurunan pertumbuhan dan produksi tanaman padi
di lapangan. Peningkatan populasi gulma E. crus-galli menyebabkan penurunan
proses fisiologi tanaman padi, seperti penurunan efisiensi produksi bahan kering
(peningkatan LAR), penurunan laju asimilasi bersih, laju tumbuh relatif, dan laju
tumbuh tanaman, serta penurunan harvest indeks. Penurunan proses fisiologi
yang terjadi pada tanaman padi, menyebabkan penurunan pertumbuhan dan hasil
produksi tanaman padi di lapangan.
Penurunan produksi GKP 10% terhadap hasil maksimum yang dapat
dicapai pada varietas Fatmawati terjadi pada saat populasi 3.2 gulma E. crus-
galli/m2, sedangkan pada varietas hibrida SL 8 SHS terjadi pada saat populasi 1.7
gulma E. crus-galli/m2. Konsekuensinya adalah bahwa penyiangan pada tanaman
padi hibrida harus segera dilakukan pada saat populasi gulma sudah mencapai 1.7
gulma E. crus-galli/m2 sedangkan pada varietas Fatmawati ketika populasi
mencapai 3.2 gulma E. crus-galli/m2.

PEMBAHASAN UMUM
Keragaman Morfologi dan Genetik serta Implikasinya dalam Manajemen
Gulma
Aksesi gulma E. crus-galli menunjukkan keragaman morfologi baik di
habitat asal maupun di rumah kaca Bogor (ketinggian tempat 250 m dpl). Pada
habitat asal, karakter jumlah daun dan jumlah anakan menunjukkan keragaman
tertinggi, sedangkan keragaman terendah ditunjukkan oleh tinggi gulma (Tabel
11). Pada habitat rumah kaca Bogor, keragaman tertinggi ditunjukkan oleh
karakter sudut daun dan jumlah daun, sedangkan keragaman terendah ditunjukkan
oleh karakter panjang daun dan jumlah anakan (Tabel 14 dan Tabel 15). Menurut
Altop et al. (2011) keragaman fenotip aksesi dipengaruhi praktek budidaya
tanaman, karakteristik tanaman, lokasi geografis, dan adanya tekanan penggunaan
herbisida.
Analisis cluster berdasarkan karakter morfologi di habitat asal
menghasilkan lima kelompok aksesi pada koefisien kemiripan 0.78 (Gambar 3),
sedangkan berdasarkan analisis komponen utama, 80.2% keragaman dapat
dijelaskan oleh tiga komponen utama. Karakter pembeda pada komponen utama I
yaitu ukuran lebar daun sedang, karakter pembeda pada komponen utama II yaitu
jumlah anakan sedikit, dan karakter pembeda pada komponen utama III yaitu
ukuran panjang malai yang panjang (Tabel 12 dan Tabel 13). Hasil ini
menunjukkan bahwa aksesi gulma E. crus-galli memiliki keragaman antar lokasi
geografis. Tasrif et al (2004) juga melaporkan adanya keragaman morfologi dan
genetik ekotipe E. crus-galli di Indonesia dari geografis yang berbeda.
Analisis cluster berdasarkan karakter morfologi di habitat rumah kaca
Bogor menghasilkan lima kelompok aksesi pada koefisien kemiripan 0.5 (Gambar
5). Berdasarkan analisis komponen utama, 76.2% keragaman dapat dijelaskan
oleh 6 komponen utama (Tabel 16). Karakter pembeda pada komponen utama I
adalah ukuran panjang malai yang panjang dan tinggi tanaman rendah. Karakter
pembeda pada komponen utama II – VI berturut-turut yaitu jumlah daun sedang,
tinggi tanaman sedang, jumlah daun sedikit, umur panen sedang, dan ukuran
malai panjang (Tabel 17).

140
Perbedaan geografis memberikan perbedaan lingkungan tumbuh dari
habitat asal ke habitat rumah kaca yang menyebabkan adanya perubahan karakter
morfologi. Karakter tinggi tanaman, jumlah anakan, jumlah daun, dan lebar daun
di rumah kaca mengalami penurunan dibandingkan dengan karakter di habitat
asal, sebaliknya panjang daun meningkat dibandingkan dengan habitat asal (Tabel
11 dan Tabel 14). Perubahan karakter ini menunjukkan adanya kemampuan
gulma E. crus-galli dalam menyesuaikan diri dengan kondisi lingkungan yang
berbeda. Kondisi habitat daerah asal aksesi gulma berbeda dengan kondisi rumah
kaca dengan suhu udara, curah hujan, kelembaban udara, dan ketinggian tempat
bervariasi, serta jenis tanah sebagian besar aluvial (Tabel 9). Sementara itu, suhu
udara harian di rumah kaca berkisar antara sekitar 31.7° - 38.5°C, kelembaban
udara rendah yakni sekitar 66.6%, dan media tanah jenis tanah latosol.
Gulma E. crus-galli memiliki kemampuan adaptasi terhadap lingkungan
tumbuh yang beragam (Barret 1983). Perubahan karakter morfologi yang
ditunjukkan di rumah kaca menunjukkan adanya ekspresi fenotipik yang
tergantung lingkungan (plastisitas fenotipik) dan kemampuan gulma E. crus-galli
menyesuaikan diri terhadap lingkungan yang spesifik disebut sebagai plastisitas
adaptif. Menurut Sultan (2003) plastisitas adaptif memainkan peranan utama
dalam distribusi organisme dan pola evolusinya. Genotip yang memiliki
kemampuan plastisitas adaptif akan menghuni kondisi lingkungan yang luas.
Wiliams et al. (1995) menyatakan bahwa plastisitas adaptif juga berkontribusi
dalam mendukung sifat invasif suatu spesies dengan membiarkan suatu spesies
cepat berkoloni dan menyebar di habitat baru tanpa menjalani seleksi lokal.
Menurut Sultan (2003) kemampuan adaptasi tersebut membiarkan gulma untuk
mengoptimalkan kemampuan bertahan hidupnya dalam periode yang paling
sesuai, sehingga menyebabkan atau memunculkan adanya ras atau individu baru.
Keragaman morfologi aksesi gulma E. crus-galli selain disebabkan oleh
lingkungan tumbuh juga disebabkan oleh keragaman genetik. Hasil analisis
cluster berdasarkan marka molekuler SSR menunjukkan bahwa aksesi
mengelompok membentuk empat sub grup (Gambar 7). Sedangkan berdasarkan
karakter morfologi, aksesi mengelompok membentuk lima kelompok baik
berdasarkan habitat asal maupun rumah kaca, namun masing-masing aksesi ada

141
yang berada pada kelompok sama dan juga ada yang terpisah di kelompok lain.
Hasil ini lebih menguatkan bahwa penampakan karakter morfologi selain
disebabkan oleh keragaman genetik juga disebabkan oleh adanya kemampuan
plastisitas fenotipik aksesi gulma E. crus-galli.
Aksesi aksesi gulma E. crus-galli yang berasal dari beberapa habitat
sawah di Provinsi Jawa Barat menunjukkan keragaman genetik yang cukup
rendah (Gambar 7). Keragaman genetik yang cukup rendah ini diduga karena
gulma E. crus-galli merupakan gulma yang menyerbuk sendiri (Honk et al. 1999).
Secara umum, spesies yang menyerbuk sendiri memiliki variasi genetik yang
rendah, sebaliknya untuk spesies yang menyerbuk silang akan memiliki variasi
genetik yang tinggi (Lin et al. 2005).
Hasil analisis cluster berdasarkan marka molekuler menunjukkan bahwa
aksesi mengelompok menjadi 4 sub grup. Sub grup A memiliki aksesi yang
berasal dari seluruh aksesi Indramayu, Cianjur, Sukabumi dan sebagian besar
aksesi dari Karawang. Semua aksesi asal Subang dan Cikampek, serta 1 aksesi
asal Karawang (K9) tergabung dalam sub grup C. Pengelompokan A dan C
menunjukkan adanya pengelompokan berdasarkan zona geografi. Secara
geografis, Cianjur, Sukabumi dan Karawang merupakan wilayah Jawa Barat
bagian barat yang berbatasan langsung, dan berdasarkan dendogram
mengelompok menjadi satu cluster. Demikian juga, Subang dan Cikampek serta
Karawang juga merupakan satu cluster C dapat dimasukkan ke dalam zona
geografis Jawa Barat bagian utara. Ketiga wilayah tersebut secara administratif
berbatasan langsung.
Perkecualian terlihat pada sub grup B dan D yang memisah dari sub grup
lain yang secara geografis berjauhan. Selain itu, secara altitude, kedua tempat
tersebut mewakili dataran rendah dan dataran tinggi. Dengan demikian,
perkecualian tersebut masih mengindikasikan pewilayahan gulma E. crus-galli
secara geografis yang kuat.
Keragaman genetik yang ditemukan pada wilayah Jawa Barat bagian
utara, bagian barat dan tengah yang secara berturut-turut diwakili oleh Subang,
Karawang dan Pangalengan mengindikasikan adanya kebutuhan program
pengendalian yang berbeda. Hasil percobaan menunjukkan bahwa aksesi gulma

142
E. crus-galli memiliki keragaman potensi alelopati (Tabel 20) dan perbedaan
potensi dalam menurunkan pertumbuhan dan produksi tanaman padi (Tabel 6 dan
Tabel 25). Hasil penelitian mengindikasikan adanya perbedaan laju pertumbuhan
gulma antar lokasi tersebut. Dari dasar tersebut, cukup menjadi pembenaran perlu
adanya cara pengendalian yang berbeda.
Gulma E. crus-galli asal dataran tinggi memiliki idiotipe tanaman yang
tinggi dan panjang daun panjang. Jika dibandingkan dengan arah pemuliaan padi
yang lebih mengarah pada pembentukan padi genjah dengan ketinggian tanaman
medium (sekitar 0.7-0.9 m), maka eksistensi gulma akan dengan mudah dapat
diidentifikasi. Perbedaan ketinggian tersebut dapat menjadi salah satu cara
pengendalian yaitu dalam pemilihan varitas tanaman padi. Demikian juga
sebaliknya, di dataran rendah yang umumnya memiliki gulma E. crus-galli
dengan batang lebih pendek akan lebih efektif jika padi yang digunakan memiliki
ideotipe lebih tinggi dari pada ketinggian maksimum gulma. Terkait pemilihan
atau formulasi herbisida untuk pengendalian secara kimia, perlu ada kajian lebih
lanjut apakah gulma asal lokasi yang berbeda memiliki tingkat resistensi herbisida
yang sama atau tidak.
Penelitian ini berimplikasi bahwa perlu adanya tindakan agronomi yang
lebih terpadu terkait dengan pengendalian gulma E. crus-galli pada wilayah Jawa
Barat. Pemeliharaan jaringan irigasi untuk menekan penyebaran gulma menjadi
salah satu prioritas. Disamping itu, perlu adanya pengendalian mutu panen dan
bibit sehingga penyebaran gulma melalui kedua material tersebut dapat dikurangi.
Potensi Alelopati Gulma E. crus-galli
Gulma dapat menurunkan pertumbuhan tanaman padi melalui pengaruh
alelopati yang ditunjukkan dengan adanya penghambatan pemanjangan radikula
dan plumula pada kecambah padi umur 14 hari setelah semai oleh ekstrak akar
gulma E. crus-galli (Tabel 18). Penghambatan tersebut diduga disebabkan oleh
senyawa kimia alelopati (allelokimia) yang dihasilkan oleh akar gulma E. crus-
galli.
Tiap aksesi gulma E. crus-galli asal Jawa Barat memiliki potensi alelopati
yang berbeda yang ditunjukkan dengan perbedaan dalam penghambatan radikula

143
dan plumua. Perbedaan penghambatan tersebut diduga disebabkan oleh
perbedaan jumlah jenis senyawa dan konsentrasi senyawa yang dihasilkan oleh
tiap aksesi gulma E. crus-galli asal Jawa Barat (Tabel 20 dan Tabel Lampiran 1-
9). Beberapa di antara senyawa yang teridentifikasi merupakan senyawa yang
bersifat alelopati pada gulma E. crus-galli seperti yang telah dilaporkan oleh
Yamamoto et al. (1999) dan Xuan et al. (2006). Senyawa yang bersifat alelopati
tersebut diantaranya adalah golongan phenolic, pthalic acid, decanoid acid,
propanoid, quinon, dan sterol. Senyawa golongan phenolic merupakan senyawa
yang teridentifikasi pada hampir semua aksesi asal Jawa Barat.
Berdasarkan potensi alelopatinya, analisis cluster mengelompokkan aksesi
gulma E. crus-galli menjadi enam kelompok, yaitu aksesi dengan penghambatan
plumula rendah dan radikula rendah (I1, I2, I5, K7, dan S3), aksesi dengan
penghambatan plumula tinggi dan radikula sedang (K5, K6, Ta1), aksesi dengan
penghambatan plumula rendah dan radikula sedang (Td2, Tc1, Tf3), aksesi
dengan penghambatan plumula sedang dan radikula rendah (Te3, K9, C4), aksesi
dengan penghambatan plumula tinggi dan radikula rendah (K3), dan aksesi
dengan penghambatan plumula tinggi dan radikula tinggi (Td3).
Keragaman Aksesi Gulma dalam Penekanan terhadap Pertumbuhan dan
Produksi Padi
Penurunan pertumbuhan dan produksi tanaman padi ditentukan
diantaranya oleh derajat kompetisi gulma E. crus-galli. Aksesi gulma E. crus-
galli asal Jawa Barat memiliki kemampuan yang berbeda dalam menurunkan
pertumbuhan dan produksi tanaman padi. Perbedaan kemampuan tersebut
disebabkan oleh perbedaan derajat kompetisi total tiap aksesi gulma E. crus-galli
asal Jawa Barat, baik dalam kemampuan kompetisi terhadap sarana tumbuh
maupun potensi senyawa alelopati yang dihasilkan. Perbedaan kemampuan
kompetisi total diduga terkait dengan keragaman genetik aksesi gulma dan
kemampuan plastisitas adaptif aksesi gulma tersebut. Menurut Altop et al.
(2011), praktek budidaya, karakteristik tanaman, geografis, dan tekanan herbisida
di suatu lokasi dapat mempengaruhi keragaman aksesi.

144
Aksesi K6 yang berasal dari Karawang yang secara genetik mirip dengan
aksesi Te3 dan K5 dan memiliki potensi alelopati yang mirip dengan K5 yaitu
tingkat penghambatan (Inhibition Rate=IR) plumula tinggi dan IR radikula sedang
menunjukkan aksesi yang paling potensial dalam menurunkan pertumbuhan dan
produksi tanaman padi di lapangan. Hal ini terlihat dari kemampuan aksesi K6
yang lebih besar dibandingkan dengan aksesi lainnya dalam menurunkan
pertumbuhan tanaman padi yaitu menurunkan tinggi tanaman, jumlah daun,
panjang daun dan lebar daun, jumlah anakan, panjang akar, bobot kering tajuk dan
akar. Aksesi K6 juga menurunkan komponen hasil dan hasil produksi padi yang
lebih besar dibandingkan dengan aksesi lainnya. Hal ini terlihat dari penurunan
jumlah anakan produktif, panjang malai, kepadatan malai, bobot gabah total, dan
bobot gabah isi serta peningkatan bobot gabah hampa.
Karakter morfologi aksesi K6 yang diduga terkait dengan kemampuan
kompetisinya terhadap tanaman padi antara lain karakter panjang daun yang
panjang, lebar daun sempit (<0.7 cm), sudut daun kecil (<16°), dan umur
berbunga lambat. Dengan karakter panjang daun yang panjang, lebar daun
sempit, dan sudut daun yang kecil diduga antar daun gulma tidak saling menaungi
sehingga proses fotosintesis gulma lebih baik, tetapi daun gulma tersebut
menutupi daun tanaman padi sehingga proses fotosintesis tanaman padi
terganggu. Selain itu, dengan masa vegetatif yang lebih panjang, maka masa
kompetisi gulma dengan tanaman padi lebih lama.
Apabila dikaitkan dengan lokasi Karawang yang intensif dalam praktek
budidaya tanaman padi, diduga aksesi asal Karawang memiliki kemampuan
adaptasi yang lebih baik pada berbagai kondisi, kemampuan plastisitas fenotipik
pada kondisi lingkungan yang berbeda, sehingga memiliki kemampuan yang kuat
dalam menurunkan pertumbuhan dan produksi tanaman padi.
Respon Tanaman Padi terhadap Tingkat Populasi Gulma E. crus-galli
Pertumbuhan dan produksi tanaman padi menurun akibat berkompetisi
dengan gulma E. crus-galli. Penurunan pertumbuhan dan produksi tanaman padi
berhubungan dengan tingkat populasi gulma E. crus-galli. Hasil ini sejalan
dengan hasil penelitian Ampong-Nyarko dan Datta (1991) bahwa salah satu faktor

145
yang menentukan tingkat kompetisi antara padi dan gulma adalah kepadatan
gulma pada pertanaman padi.
Semakin tinggi tingkat populasi gulma E. crus-galli secara umum
pertumbuhan tanaman padi menurun. Pertumbuhan yang menurun tersebut
terlihat dari penurunan jumlah anakan, jumlah daun, panjang akar, dan ILD.
Komponen hasil padi yang meliputi jumlah anakan produktif, panjang malai,
jumlah gabah per malai, persentase gabah isi dan hampa juga semakin menurun
dengan semakin meningkatnya populasi gulma E. crus-galli. Penurunan
komponen hasil produksi menyebabkan penurunan hasil tanaman padi yaitu
penurunan bobot gabah kering panen dan gabah kering giling. Purba (2007)
melaporkan bahwa semakin tinggi kerapatan E. crus-galli per m2
Respon tanaman padi terhadap tingkat populasi gulma E. crus-galli di
lapangan berbeda antar varietas tanaman padi. Varietas Fatmawati yang
merupakan varietas unggul tipe baru menunjukkan tinggi tanaman, jumlah
anakan, jumlah daun yang paling tinggi dibandingkan dengan varietas lainnya
pada tingkat populasi gulma E. crus-galli yang sama. Varietas hibrida SL 8 SHS
menunjukkan tinggi tanaman yang paling rendah, penurunan jumlah anakan dan
jumlah daun yang paling tinggi dibandingkan dengan varietas lainnya ketika
populasi gulma E. crus-galli semakin meningkat (Gambar 30, 31, dan 32).
, maka
penurunan hasil tanaman padi semakin besar. Menurut Tindall et al. (2003)
penurunan hasil akibat kompetisi terjadi karena penurunan jumlah anakan, jumlah
malai, dan jumlah gabah per malai.
Berdasarkan penurunan jumlah anakan, varietas Ciherang sensitif terhadap
gulma E. crus-galli yang ditunjukkan dengan penurunan jumlah anakan produktif
sebesar 42.2% pada populasi 4 gulma E. crus-galli/m2 dan sebesar 88.7% ketika
populasi 12 gulma E. crus-galli/m2, panjang malai menurun ketika populasi 12 E.
crus-galli/m2, sedangkan varietas yang lainnya tidak mengalami penurunan
panjang malai pada berbagai populasi gulma. Varietas Ciherang juga mengalami
penurunan kepadatan malai mulai populasi 8 gulma E. crus-galli/m2, sedangkan
Inpari dan hibrida mengalami penurunan kepadatan malai pada saat populasi 12
gulma E. crus-galli/m2. Produksi varietas Ciherang baik dalam bentuk GKP

146
maupun GKG menunjukkan produksi yang paling rendah dibandingkan dengan
varietas lainnya.
Varietas Fatmawati mengalami penurunan jumlah anakan produktif yang
lebih rendah dibandingkan dengan varietas lainnya pada tingkat populasi gulma
yang sama, sedangkan panjang malai, kepadatan malai, dan persentase hampa
tidak dipengaruhi oleh populasi gulma. Berdasarkan peubah komponen hasil ini,
varietas Fatmawati merupakan varietas yang memiliki kemampuan kompetisi
yang kuat terhadap gulma E. crus-galli, namun memiliki persentase kehampaan
yang tiga kali lebih besar dibandingkan dengan varietas lainnya pada kondisi
tanpa gulma (Gambar 42), sehingga produksi GKP dan GKG Fatmawati lebih
rendah dibanding Inpari. Varietas Inpari 6 JT menunjukkan produksi GKP dan
GKG tertinggi.
Hasil penelitian ini menunjukkan bahwa tiap varietas padi sawah memiliki
tanggap yang berbeda terhadap kepadatan populasi gulma E. crus-galli. Varietas
yang memiliki postur tegak dengan tinggi tanaman tinggi, dan ukuran daun yang
luas, seperti varietas Fatmawati relatif lebih kuat dalam berkompetisi dengan
gulma E. crus-galli. Sebaliknya, varietas hibrida menunjukkan varietas yang
sensitif terhadap gulma E. crus-galli.
Fisiologi Kompetisi Padi-Gulma
Kompetisi antara tanaman padi dan gulma E. crus-galli berpengaruh
terhadap proses fisiologi tanaman padi dan proses fisiologi gulma itu sendiri.
Penurunan pertumbuhan dan produksi yang terjadi pada tanaman padi merupakan
akibat dari hambatan proses fisiologi yang terjadi pada tanaman padi. Proses-
proses fisiologi yang terjadi pada tanaman padi dapat dikuantifikasi melalui
peubah-peubah proses fisiologi, seperti net assimilation rate (NAR), relative
growth rate (RGR) dan crop growth rate (CGR), dan leaf area ratio (LAR).
Kompetisi antara tanaman padi dengan gulma terjadi baik di bawah
permukaan tanah maupun di atas permukaan tanah. Kompetisi di bawah
permukakan tanah ditandai dengan adanya penurunan panjang akar dan bobot
akar, serta perbedaan kandungan hara tajuk dan serapan hara pada tanaman padi.
Penurunan panjang akar berbeda antar varietas tanaman padi dan berbeda antar

147
tingkat populasi gulma. Penurunan panjang akar ini menyebabkan serapan hara
tanaman padi terganggu. Efisiensi serapan hara menunjukkan banyaknya bahan
kering yang diproduksi oleh tanaman setiap satuan hara yang diserap. Varietas
Inpari 6 JT menunjukkan kandungan hara P yang rendah tetapi menunjukkan
efisiensi serapan hara P yang paling tinggi diantara varietas yang diuji, sedangkan
varietas Fatmawati menunjukkan kandungan P yang tinggi, tetapi menunjukkan
efisiensi serapan hara P yang paling rendah.
Kompetisi di atas permukaan tanah terjadi dalam memperebutkan sinar
matahari untuk proses fotosintesis. Proses fotosintesis pada tanaman padi
terganggu oleh naungan daun gulma E. crus-galli. Secara umum, semua varietas
padi yang dicobakan mengalami penurunan tinggi tanaman, jumlah anakan, dan
jumlah daun. Akibatnya, ILD tanaman padi menurun.
Produksi berat kering biomass dipengaruhi oleh indeks luas daun (Weng et
al. 1982). Hasil penelitian menunjukkan bahwa varietas Ciherang dengan ILD
yang paling rendah menunjukkan berat kering biomass total yang paling rendah,
sedangkan varietas Fatmawati dan Inpari 6 JT memiliki ILD yang tinggi
menunjukkan bobot kering biomass total yang paling tinggi. ILD yang tinggi
menentukan kapasitas fotosintesis yang lebih baik. Hal ini ditunjukkan oleh leaf
area ratio varietas Fatmawati yang lebih rendah atau lebih efisien dalam produksi
bahan kering. Menurut Mia et al. (2011), ILD yang tinggi menyebabkan kapasitas
fotosintesis lebih baik, sehingga produksi bahan kering meningkat. Chen et al.
(1991) menyatakan bahwa produksi bahan kering berkorelasi positif terhadap
hasil gabah. Penurunan ILD secara langsung akan mempengaruhi laju
pertumbuhan relatif (RGR) dan laju pertumbuhan tanaman (CGR).
Untuk menghasilkan produksi yang tinggi, maka tanaman padi harus
mengalokasikan hasil asimilatnya dengan proporsi yang lebih banyak ke arah
pembentukan gabah. Hasil penelitian menunjukkan adanya perbedaan alokasi
asimilat pada tiap varietas akibat persaingan dengan gulma E. crus-galli yang
ditunjukkan dengan harvest indeks (HI). Semakin tinggi tingkat populasi gulma
E. crus-galli, maka semakin rendah proporsi hasil asimilat yang ditranslokasikan
ke bagian biji.

148
Harvest indeks (HI) berkorelasi dengan produksi gabah (Cui-Jing et al.
2000). Varietas Fatmawati dan Ciherang memiliki HI yang lebih rendah
dibandingkan dengan varietas Inpari 6 JT dan hibrida SL 8 SHS. Hal ini berarti
bahwa proporsi hasil asilmilat yang ditranslokasikan ke gabah pada varietas
Fatmawati dan Ciherang lebih rendah. Proporsi asimilat dari source ke sink
(gabah) yang rendah pada varietas Fatmawati, menyebabkan persen kehampaan
meningkat sehingga produksi gabah menurun, meskipun bobot 1000 butir tinggi.
Pada varietas Ciherang, rendahnya proporsi hasil asimilat yang ditranslokasikan
ke gabah menyebabkan bobot 1000 butir rendah, sehingga produksi juga rendah.
Hasil penelitian ini memberikan gambaran bahwa gulma E. crus-galli
yang tersebar di beberapa lokasi di Jawa Barat perlu mendapat perhatian yang
serius, terutama dalam upaya penyelamatan produksi padi nasional. Wilayah
Provinsi Jawa Barat merupakan penyumbang terbesar produksi padi nasional
dengan varietas padi yang paling luas adalah varietas Ciherang mencapai 318.225
hektar tertanam (Dinas Pertanian Provinsi Jawa Barat 2011). Infestasi gulma pada
tanaman padi varietas Ciherang perlu diwaspadai agar kehilangan hasil dapat
diselamatkan. Program penanaman varietas hibrida di seluruh Indonesia juga
perlu kehati-hatian khususnya dalam menghadapi kehilangan hasil akibat infestasi
gulma E. crus-galli. Pengendalian gulma secara terpadu perlu dilakukan mulai
dari upaya penyediaan benih bermutu yang bebas dari tercampurnya biji gulma E.
crus-galli, pengendalian gulma pada tahap awal perkembangan, pencegahan
penyebaran melalui hasil panen atau bagian-bagian tanaman yang lain,
penggunaan varietas yang tahan terhadap kompetisi gulma, serta perakitan
varietas yang tahan terhadap gulma E. crus-galli di lapangan.

KESIMPULAN DAN SARAN
Kesimpulan
1. Aksesi gulma E. crus-galli asal Jawa Barat menunjukkan keragaman
morfologi dan genetik. Aksesi dari lokasi geografis yang berbeda
menunjukkan perbedaan morfologi. Keragaman morfologi aksesi selain
disebabkan oleh genetik juga disebabkan oleh perbedaan lingkungan tumbuh.
Keragaman morfologi pada lingkungan tumbuh yang berbeda disebabkan
adanya kemampuan plastisitas fenotipik dan kemampuan mimikri aksesi
gulma E. crus-galli.
2. Analisis cluster berdasarkan karakter morfologi dari habitat asal
menghasilkan 5 kelompok pada koefisien kemiripan sebesar 0.78, sedangkan
berdasarkan karakter morfologi di rumah kaca (250 m dpl) membentuk 5
kelompok pada koefisien kemiripan 0.64 dengan anggota kelompok aksesi
yang berbeda.
3. Analisis cluster berdasarkan penanda molekuler SSR menghasilkan 4 sub
grup dengan koefisien kemiripan 0.86, dengan sebagian besar grup
mengelompok pada zona geografi Jawa Barat bagian barat dan bagian utara
(pantai utara jawa) dengan pusat keragaman adalah Subang, Karawang dan
Pangalengan. Keragaman genetik ini dapat disebabkan oleh perpindahan
material genetik melalui hasil panen atau melalui irigasi, isolasi jarak, dan
kemungkinan adanya mutasi.
4. Aksesi gulma E. crus-galli asal Jawa Barat memiliki potensi alelopati
berdasarkan penghambatan plumula dan radikula kecambah padi. Senyawa
alelopati potensial yang teridentifikasi di dalam ekstrak akar aksesi E. crus-
galli diantaranya adalah golongan senyawa phenolic, phtalic acid, decanoid
acid, propanoid, quinon, dan sterol.
5. Berdasarkan potensi alelopatinya, analisis cluster menghasilkan enam
kelompok aksesi pada koefisien kemiripan 0.72 yaitu aksesi dengan IR
plumula rendah dan IR radikula rendah, IR plumula tinggi dan IR radikula
sedang, IR plumula rendah dan IR radikula sedang, IR plumula sedang dan IR

150
radikula rendah, IR plumula tinggi dan IR radikula rendah, dan IR plumula
tinggi dan IR radikula tinggi.
6. Aksesi gulma E. crus-galli menunjukkan perbedaan potensi dalam
menurunkan pertumbuhan dan hasil tanaman padi. Kemampuan ini terkait
dengan sifat morfologi dan potensi alelopati aksesi tersebut. Aksesi K6 asal
Karawang menunjukkan kemampuan tertinggi dalam menurunkan
pertumbuhan dan produksi tanaman padi, baik di rumah kaca maupun di
lapangan. Kemampuan ini terkait dengan karakter morfologi aksesi K6 yaitu
panjang daun panjang, lebar daun sempit, sudut daun kecil, dan umur
berbunga lambat, serta memiliki IR plumula tinggi dan IR radikula sedang.
7. Gulma E. crus-galli memiliki derajat kompetisi yang lebih besar
dibandingkan dengan tanaman padi ketika populasi gulma lebih tinggi
daripada populasi tanaman padi berdasarkan koefisien pendesakan.
Berdasarkan nilai agresivitas, gulma E. crus-galli lebih kuat berkompetisi
dibandingkan tanaman padi ketika populasi padi dan gulma seimbang
ataupun populasi gulma lebih tinggi daripada populasi tanaman padi.
8. Derajat kompetisi gulma ditentukan oleh tingkat populasi gulma E. crus-galli
di lapangan. Semakin tinggi populasi gulma E. crus-galli, maka
pertumbuhan dan produksi tanaman padi semakin menurun.
9. Setiap varietas memiliki respon yang berbeda terhadap tingkat populasi
gulma E. crus-galli. Varietas Fatmawati menunjukkan varietas yang lebih
kompetitif sedangkan varietas hibrida SL 8 SHS menunjukkan varietas yang
sensitif terhadap populasi gulma E. crus-galli.
10. Kompetisi antara tanaman padi dengan gulma E. crus-galli menghambat
proses fisiologi tanaman padi yang ditunjukkan dengan penurunan peubah
proses fisiologi, seperti ILD, NAR, RGR, CGR, dan peningkatan LAR.

151
Saran
1. Tindakan agronomi secara terpadu perlu dilakukan dalam pengendalian
gulma E. crus-galli, termasuk dalam hal pemeliharaan jaringan irigasi untuk
menekan penyebaran gulma dan perpindahan material genetik.
2. Penggunaan benih padi yang bebas dari benih gulma E.crus-galli sangat
dianjurkan untk mencegah penyebaran antar aksesi gulma E. crus-galli.
3. Pengendalian gulma E. crus-galli dilakukan sedini mungkin pada tahapan
perkembangan tanaman padi untuk menghindari persaingan tanaman padi
dengan gulma E. crus-galli.
4. Untuk menyelamatkan kehilangan 10% hasil produksi akibat gulma pada
varietas Fatmawati penyiangan harus dilakukan ketika populasi gulma
mencapai 3.2 gulma E. crus-galli/m2, sedangkan pada varietas hibrida SL 8
SHS ketika populasi 1.7 gulma E. crus-galli/m2
5. Sensitivitas terhadap gulma E. crus-galli perlu dimasukkan dalam program
pemuliaan tanaman padi, termasuk kepekaannya terhadap alelopati gulma E.
crus-galli.
.

DAFTAR PUSTAKA
Ahn JK, Chung IM. 2000. Allelopathic potential of rice hulls on termination and seedling growth of barnyardgrass. Agron. J. 92: 1162–1167.
Ali MA. 1985. Crop-weed competition. Abstracts of papers of Ann. Conf. Ind. Soc. Weed Sci., pp: 78.
Ali MA, Sankaran S. 1984. Crop weed competition in direct seeded lowland and upland bunded rice. Ind. J. Weed Sci., 19: 90-96.
Altop EK, Mennan, H. 2011. Genetic and morphology diversity of Echinochloa crus-galli population from different origins. Phytoparasitica 39: 93-102.
Ampong-Nyarko K, De Datta SK. 1991. A Handbook for Weed Control in Rice. International Rice Research Institute. Los Banos. Phillipines. 191 p.
Anten NPR, Hirose T. 1998. Biomass allocation and light partitioning among dominant and subordinate individuals in Xanthium canadense stands. Ann. Bot. 82: 665–673.
Aoki D, Yamaguchi H. 2008. Genetic relationship between Echinochloa crus-galli and Echinochloa oryzicola accessions inferred from internal transcribed spacer and chloroplast DNA sequences. Weed Biol. Manag. 8 : 233–242.
Ass GJ, Raman R, Crump NS. 2003. An investigation of genetic variation in Carthamus lanatus in New South Wales, Australia, using intersimple sequence repeats (ISSR) analysis. Weed Res. 43: 208-213.
Azmi M, Baki BB. 1995. The succession of noxious weeds in tropical asian rice fields with emphasis on Malaysia rice ecosystems, p. 140-148. The 5th Asian-Pacific Weed Science Society Conference. Tsukuba, Japan, July, 24-30.
Badan Litbang Pertanian. 2011. http://www.litbang.deptan.go.id
Badan Pusat Statistik [BPS]. 2008.
[12 Oktober 2011].
http://www.bps.go.id. [16 Desember 2008].
Badan Pusat Statistik [BPS]. 2010. http://www.bps.go.id. [12 Agustus 2010].
Badan Pusat Statistik [BPS]. 2011. http://www.bps.go.id. [18 Oktober 2011].
Baki BB, Azmi M. 2003. Echinochloa aggregates in Malaysia- ecology and management. In Kim KU, Labadra L (eds). Echinochloa Control in Rice. Kyungpook National University. Pp37-52.
Baltazar AM, De Datta SK. 1992. Weed management in rice. Weed Abstacts 41: 495-507.

153
Bahrendt S, Hanf M. 1979. Grass Weeds in World Agriculture. Limburgerhof. BASF. 137 p.
Barret SCH. 1983. Crop mimicry in weeds. Economic Botany 37: 255-282.
Becker M, Johnson DE, Wopereis MCS, Sow A. 2003. Rice yield gaps in irrigated systems along an agro-ecological gradient in West Africa. J. Plant Nutr. Soil Sci. 166: 61–67.
Caton BP, Mortimer M, Hill JE. 2004. Weeds of Rice in Asia. IRRI. Los Banos. 116p.
Chen WF, Xu ZJ, Zhan LB, Yang SR. 1991. Studies on canopy properties and their relationship to dry matter production in Japonica rice cultivars with different plant types. Chinese J. Rice Sci. 5: 67-74.
Chin DV. 2001. Biologi and Management of Barnyardgrass, red sprangletop and weedy rice. Weed Biol. Manag. (1): 37.
Chin DV. 2003. Ecophysiologycal characteristics of barnyardgrass and its management in Vietnam. In : Kim KU, Labrada R (eds). Echinochloa Control in Rice. Kyungpook National University. Pp.21-27.
Clements DR, Benoit DL, Murphy SD, Swanton CJ. 1996. Tillage effects on weed seed return and seedbank composition. Weed Sci. 44: 314–322.
Chung IM, Kim KH, Ahn JK, Lee SB, Kim SH, Hahn SJ. 2003. Comparison of allelopathic potential of rice leaves, straw, and hull extracts on barnyardgrass. Agron. J. 95: 1063-1070.
Cui-Jing A, Kusutani, Toyata M, Asanuma K, Cui J. 2000. Studies on the varietal differences of harvest index and morphological characteristic of rice. Jap. J. Crop Sci. 69: 359-364.
Danquah EY, Hanley SJ, Brookes RC, Aldam A, Karp A. 2002. Isolation and characterization of microsatellites in Echinochloa (L.) Beauv. spp. Molec. Ecol. Note 2: 54-56.
De Datta SK. 1981. Principle and Practices of Rice Production. John Wiley and Sons Inc. New York. 148p.
De Wit CT. 1960. On competition. Versl. ladbouwk. Onderz. 66(8). 82p.
De Witt TJ, Scheiner SM. 2004. Phenotypic Plasticity : functional and conceptual approach. Oxford University Press. Oxford.
Dinas Pertanian Provinsi Jawa Barat. 2011. Inventarisasi luas penyebaran varietas padi musim tanam 2009/2010. http://diperta.jabarprov.go.id. [10 Oktober 2011].

154
Dirjen PLA. 2005. Strategi dan Kebijakan Pengelolaan Lahan. Direktorat Jenderal Pengelolaan Lahan dan Air, Departemen Pertanian. Jakarta
Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 19:11-15.
Duke JA. 1996. Handbook of Energy Crops. http://www.hort.purdue.edu.
Eussen JHH, Zulfadli M. 1981. Upland rice-weed competiton as affected by nitrogen application and the time and duration of the competition. P : 97-107. Dalam Mangoensoekarjo S (ed.). Prosiding Konferensi VI Himpunan Ilmu Gulma Indonesia (HIGI). Sumatera Utara, 12-14 Februari.
Fishel F. 2000. Barnyardgrass-Billion Dollar Grass. Integrated Pest and Crop Management Newsletter. 109 p.
Fossati T, Labra M, Castiglione S, Failla O, Scienza A, Sala F. 2001. The use of AFLP and SSR molecular markers to decipher homonyms and synonyms in grapevine cultivars: the case of the varietal group known as ‘‘Schiave’’. Theor. Appl. Genet. 102: 200–205
Froud-Williams RJ, Chancellor RJ, Drennan DSH. 1984. The effects of seed burial and soil disturbance on emergence and survival of arable weeds in relation to minimal cultivation. J. Appl. Ecol. 21: 629–641.
Galinato MI, Moody K, Piggin CM. 1999. Upland Rice Weeds of South and Southeast Asia. International Rice Research Institute. Los Banos. Phillipines. 155p.
Gealy DR, Wailes EJ, Estorninos LJ, Chavezrc. 2003. Rice cultivar differences in suppression of barnyardgrass (Echinochloa crus-galli) and economics of reduced propanil rates. Weed Sci. 51: 601–609.
Gibson KD, Foin TC, Hill JE. 1999. The relative importance of root and shoot competition between water-seeded rice and Echinochloa phyllopogon. Weed Research 39: 181-190.
Guntoro D, Chozin MA, Santosa E, Soekisman T, Burhan AH. 2009. Kompetisi antara ekotipe Echinochloa crus-galli pada beberapa tingkat populasi dengan padi sawah. J. Agron. Indonesia 37 (3) : 202 – 208.
Haefele SM, Johnson DE, Mbodj D, Wopereis MCS, Miezan KM. 2004. Field screening of diverse rice genotypes for weed competitiveness in irrigated lowland ecosystems. Field Crops Res. 88: 39–56.
Hiu LB. 1998. Statistical Genomics : linkage, mapping, and QTL analysis. CRC press LLC. p 611.

155
Holm LG, Plucknett DL, Pancho JV, Herberger JP. 1977. The World’s Worst Weeds. Distribution and Biology. East-West Center, University Press of Hawai, Honolulu, Hawai. Pp. 32-46.
Honk A, Martinkov Z. 1996. Geographic variation in seed dormancy among populations of Echinochloa crus-galli. Oecologia 108:419 – 423.
Inderjit, Keating KI. 1999. Allelopathy: principles, procedures, processes, and promises for biological control. In. : Sparks DL (ed). Adv. Agron. 67: 141-231.
Islam MF, Karim SMR. 2003. Effect of population density of Echinochloa crus-galli and Echinochloa colona on rice. P : 275-281. Proceedings I The 19th
Islam MdF, Karim SMR, Haque SMA, Islam MdS. 2003. Effect of population density of Echinochloa crus-galli and Echinochloa colonum on Rice. Pakistan J. Agron. 2(3): 120-125.
Asian-Pacific Weed Science Society Conference. Manila-Philippines, March, 17-21.
Itoh K. 1991. Life Cycles of Rice Field Weeds and Their Management in Malaysia. Tropical Agriculture Research Center. Japan. 92 p.
Jarret RL, Gawel N. 1995. Molecular markers, genetic diversity and systematics in Musa. In: Gowen S (ed.). Bananas and Plantains. London, Chapman and Hall. pp. 66-83.
Johnson DE, Wopereis MCS, Mbodj D, Diallo S, Pewers S, Haefele, SM. 2004. Timing of weed management and yield losses due weeds in irrigated rice in the Sahel. Field Crops Res. 85: 31–42.
Juanedi A, Chozin MA, Kim KH. 2006. Perkembangan terkini kajian alelopati. Hayati 13(2): 79-84.
Kim KU. 1994. Ecophysiology of Echinochloa species and their management, p. 18-25. In Sastroutomo SS, Auld BA. (Eds.). Proceeding of an International Workshop on Appropriate Weed Control in Southeast Asia. Kuala Lumpur, March, 12-17.
Kropff MJ, van Laar HH. 1993. Modelling Crop-Weed Interactions. CAB International. Great Britain. 277 p.
Lin WX, He HQ, Chen XX, Xiong J, Song BQ, Liang YY, Liang KJ. 2005. Use of ISSR molecular marker approach to estimate genetic diversity in rice and barley allelopathy. http://www.regional.org.au/au/allelopathy. [23 Februari 2007].
Liu J, Wang GX, Wei L, Wang CM. 2008. Reproductive allocation patterns in different density populations of spring wheat. J. Integr. Plant Biol. 50: 141–146.

156
Maun MA, Barrett SCH. 1986. The biology of Canadian weeds.77. Echinochloa crus-galli (L.) Beauv. Can. J.Plant Sci. 66: 739-759.
McGregor CE, Lambert CA, Greyling MM, Louw JH, Warnich L. 2000. A comparative assessment of DNA fingerprinting techniques (RAPD, ISSR, AFLP and SSR) in tetraploid potato (Solanum tuberoseum L.) germplasm. Euphytica. 113 :135-144.
Mia MAB, Shamsuddin ZH. 2011. Physio-morphological appraisal of aromatic fine rice (Oryza sativa L.) in relation to yield potential. Int. J. Bot. 7(3): 223-229.
Moenandir J. 1993. Pengantar Ilmu dan Pengendalian Gulma : Ilmu Gulma-Buku I. Rajawali Press. Jakarta. 107 hal.
Moodie M, Finch RP, Marshall G. 1997. Analysis of genetic variation in wild mustard (Sinapsis arvensis) using molecular markers. Weed Sci. 45: 102-107.
Nissen SJ, Masters RA, Lee DJ, Rowe ML. 1995. DAN-based marker system to determine genetic diversity of weedy species and their application to biocontrol. Weed Sci. 43: 504-513.
Ortiz R, Vuylsteke D, Swennen R. 1993. Phenotypic variation and grouping of Musa germplasm. Agronomy Abstracts. Am. Soc. Agronomy, Madison, Wi: 192. Abstract.
Partzsch M, Bachmann U. 2011. Is Campanula glomerata threatened by competition from expanding grasses? Results from a 5-year pot-experiment. Plant Ecol. 212: 251–261
Perera KK, Ayres PG, Gunasena HPM. 1992. Root growth and the relative importance of root and shoot competition in the interactions between rice (Oryza sativa) and Echinochloa crus-galli. Weed Res. 32: 67-76.
Plante XT, Hendry P. 2011. The consequences of phenotipic plasticity for ecological speciation. J. Evol. Biol. 24: 326-342.
Powell W, Morgante M, Andre C, Hanafey M, Vogel J, Tingey S, Rafalski A. 1996. The comparison of RFLP, RAPD, AFLP, and SSR (microsatellite) Markers for Germplasm analysis. Molec. Breeding 2: 225-238.
Purba E. 2007. Respons padi terhadap kerapatan jajagoan (Echinochloa crus-galli). Jurnal Ilmu-ilmu Pertanian Indonesia 1: 62-68.
Puspendra, Gupta K, Rajeev KV, Prasad M. 2002. Molecular Markers : Principles and Methodology. In : Jain SM, Brar DS, Ahloowalia BS (eds.) Molecular Techniques in Crop Improvement. p9-54. Kluwer Academic publishers. Netherland.

157
Putnam AR, Weston LA. 1986. Adverse impact of allelopathy in agricultural systems. In : Putnam AR, Tang CS (ed). The Science of Allelopathy. New York: John Wiley & Sons. pp 43-56.
Ranasinghe LL, Crabtree GD. 1999. Plant characteristic associated with rice (Oryza sativa L.)-barnyardgrass (Echinochloa crus-galli L. Beauv.) competition. P : 99-104. Proceedings I (A) The 17th
Rauf AW, Syamsuddin T, Sihombing SR. 2007. Peranan pupuk NPK pada tanaman padi. Dinas Pertanian. Irian Jaya.
Asian-Pacific Weed Science Society Conference. Bangkok-Thailand, November, 22-27.
Ribaut JM, Bänzinger M, Betran J, Jiang C, Edmeades GO, Dreher K, Hoisington D. 2002. Use of Molecular Markers in Plant breeding : Drought Tolerance Improvement in Tropical maize. In : Manjit SK (ed). Quantitative Genetics, Genomics and Plant Breeding. CAB International. UK.
Rice EL. 1974. Allelopathy. Academic Press. New York.35p.
Rice EL. 1995. Biological Control of Weeds and Plant Diseases: Advances in Applied Allelopathy. Norman: Univ of Oklahoma.
Rohlf FJ. 1998. On applications of geometric morphometrics to studies of ontogeny and phylogeny. Sys. Bio. 47(1): 147-158.
Sastroutomo S. 1990. Ekologi Gulma. PT. Gramedia Pustaka Utama. Jakarta. 216 hal.
Santamaria L, Figuerola J, Pilon JJ, Mjelde M, Green AJ, de Boer T, King RA, Gornall RJ. 2003. Plant performance across latitude : the role of plasticity and local adaptation in an aquatic plant. Ecology 84(9): 2454-2461.
Savary S, Willocquet L, Elazegui FA, Castilla NP, Teng PS. 2000. Rice pest constraints in tropical Asia: quantification of yield losses due to rice pests in a range of production situations. Plant Dis. 84, 357–369.
Savary S, Srivastava RK, Singh HM, Elazegui FA. 1997. A characterization of rice pests and quantification of yield losses in the rice–wheat system of India. Crop Protect. 16: 387–398.
Scotti I, Magni F, Paglia GP, Morgante M. 2002. Trinucleotide microsatellites in Norway spruce (Picea abies) : their features and the development of molecular markers. Theor. Appl. Genet. 106: 40-50.
Schlichting CD. 1986. The evolution of phenotipic plasticity in plants. Ann. Rev. Ecol. Syst. 17: 667-693.
Seigler DS. 2006. Basic pathways for the origin of allelopathic compounds. In : Manuel J. Reigosa MJ, Pedrol N, González L (eds). Allelopathy: A Physiological Process with Ecological Implications. Pp.11-61.

158
Singh M, Saxena MC, Abu-Irmaileh BE, Al-thababi SA, Haddad NI. 1996. Estimation of critical period of weed kontrol. Weed Sci. 44: 272 – 283.
Singh HP, Batish DR, Kohli RK. 2003. Allelopathic interaction and allelochemicals: new possibilities for sustainable weed management. Crit. Rev. Plant Sci. 22:239-311.
Smith JRJr. 1968. Weed competition in rice. Weed Sci. 16: 252-254.
Smith RJJr. 1983. Weeds of major economic importance in rice and yield losses due to weed competition. In: Proceeding of the Conference on Weed Control in Rice, International Rice Research Institute : 19-36.
Sobir TO, Murata M, Motoyoshi F. 2000. Molecular characterization of the SCAR markers tightly linked to the Tm-2 locus of the genus Lycopersion. Theor. Appl. Genet. 101: 64–69.
Soejono, Indriyani S, Widhyastuti NK. 2001. Rekonstruksi genetik 12 kultivar pisang (Musa paradisiacal L.) koleksi Kebun Raya Purwodadi Pasuruan. Dalam Wardiyati, Ashari, Aini dan Suryanto (eds). Prosiding Seminar Nasional Hortikultura, Kongres PERHORTI. BUKU I. Fakultas Pertanian Unibraw. Malang. Hal 540-545.
Soerjani, et al. 1987. Weeds of Rice in Indonesia. Balai Pustaka. Jakarta. 716 hal.
Son DH, Thil HL, Noguchi HK. 2010. Allelopathic potential and isolation process of allelopathic substances in Barnyardgrass (Echinochloa crus-galli). Omonrice 17: 143-146.
Suardi D, Pane H. 1983. Daya saing beberapa varietas padi terhadap gulma. Jurnal Penelitian dan Pengembangan Pertanian 3: 63-66.
Sultan SE. 2000. Phenotipic plasticity for plant development, function and life history. Trends Plant Sci. 5: 537-542.
Sultan SE. 2003. Phenotipic plasticity in plants : a case study in ecological development. Evolution & Development 5(1): 25-33.
Sultana R. 2000. Competitive ability of wet-seeded boro rice against Echinochloa crusgalli and Echinochloa colonum. M.S. Thesis, BAU, Mymensingh, Bangladesh, pp: 36-50.
Sulyo Y. 1997. Beberapa teknik sidik DNA yang dapat digunakan sebagai penanda genetik dalam pemuliaan. Kelompok peneliti plasma nutfah dan pemuliaan, Balai Penelitian Tanaman Hias. Jakarta. 6 halaman.
Sung SJS, Leather GR, Hale MG. 1987. Development and germination of Barnyardgrass (Echinochloa crus-galli) seeds. Weed Sci. 35: 211-215.

159
Susanto U, Daradjat AA, Suprihatno B. 2003. Perkembangan pemuliaan padi sawah di Indonesia. Jurnal Litbang Pertanian 22(3): 125-131.
Sutrisno DP, Turanto S. 1981. Pengaruh jawan (Echinochloa crus-galli L) terhadap pertumbuhan dan produksi padi IR-36. Hal 229-235. Dalam Mangoensoekarjo S (ed.). Prosiding Konferensi VI Himpunan Ilmu Gulma Indonesia (HIGI). Sumatera Utara, 12-14 Februari.
Swennen R, Vuylsteke D, Ortiz R. 1995. Phenotypic diversity and patterns of variation in West and Central African Plantains (Musa spp., AAB Group Musaceae). Econ. Bot. 49 (3):320-327.
Takeda T. 1961. Studies on the photosynthesis and production of drymatter in the community of the rice plants. Japan J. Bot. : 129-137.
Tasrif A, Juraimi AS, Kadir J, Napis S, Sastroutomo SS. 2003. Keragaman morfologi dan molekuler ekotipe gulma jejagoan (Echinochloa crus-galli var. crus-galli L.P. Beauv). J. Gulma Tropika (1): 43-48.
Tasrif A, Juraimi AS, Kadir J, Sastroutomo S S, Napis S. 2004. Genetic diversity of Echinochloa crus-galli var. crus-galli (L.) Beauv. (Barnyardgrass: Poaceae) ecotypes in Malaysia and Indonesia as revealed by RAPD markers. Asian J. Plant Sci. 3: 231-238.
Thompson JD. 1991. Phenotipic plasticity as a component of evolutionary change. Trend Ecol. Evol. 6: 246-249.
Tindall KV, Williams BJ, Stout MJ, Geaghan JP, Leonard BR, Webster EP. 2005. Yield components and quality of rice in response to graminaceous weed density and rice stink bug populations. Crop Protection 24 (11) : 991-998.
Tjitrosemito S. 1994. Integrated management of paddy and aquatic weeds in Indonesia, p. 20-31. Proceedings of the International Seminar Biological Control and Integrated Management of Paddy and Aquatic Weeds in Asia. Japan, Oktober, 19-25.
Tobing IE, Chozin MA. 1980. Masa kritis padi sawah berumur genjah terhadap persaingan gulma. Buletin Agonomi (XI): 1-6.
Trung, HM, Tan NT, Cung HA. 1995. Present status and prospect of weed control in rice in Vietnam. In Proceeding of the 15th
Tungate KD, Israel DW, Watson DM, Rufty TW. 2007. Potential changes in weed competitiveness in an agroecological system with elevated temperatures. Environ. Exp. Bot. 60 : 42–49.
Asian-Pasific Weed Science Society Conference. Tsukuba. Japan. p.601-606.

160
Vuylsteke D, Swennen R, Wilson GF, De Langhe. 1988. Phenotypic variation among in vitro propagated plantain (Musa spp. cv. AAB). Sci. Hort. 36 (1-2): 79-88.
Waterhouse DF. 1994. Biological Control of Weeds: Southeast Asian Prospects. ACIAR. Canberra.
Weng JH, Takeda T, Agata W, Hakeyama S. 1982. Studies on dry matter production in rice plant. Varietal differences in dry matter productivity before heading. Japanese J. Crop. Sci. 51: 510-518.
Whilley RW. 1979. Intercropping –its importance and research needs. Part I. Competition and yield advantages. Field Crop 32 :1-10.
Williams DG, Mack RN, Black RA. 1995. Ecophysiology of introduced Pennisetum setaceum on Hawaii: the role of phenotypic plasticity. Ecology 76: 1569-1580.
Xuan TD, Chung M, Khanh TD, Tawata S. 2006. Identification of phitotoxic substance from early growth of barnyardgrass (Echinochloa crus-galli) root exudates. J. Chem. Ecol. 32:895-906.
Yabuno T. 1983. Biology of Echinochloa species. In Weed Control in Rice. IRRI. P.307-318.
Yamamoto T, Yokotani-Tomita K, Kosemura S, Yamamura S, Yamada K, Hasegawa K. 1999. Allelopathic substance exuded from a serious weed, germinating barnyardgrass (Echinochloa crus-galli L.) roots. J. Plant Growth Regul. 18: 65-67.
Yamasue Y. 2003. Echinochloa aggregates in Malaysia- ecology and management. In Kim KU, Labadra L (eds). Echinochloa Control in Rice. Kyungpook National University. Pp37-52.
Yuan ZY, Li LH, Han XG, Jiang FH, Zhao MX, Lin GH. 2004. Effects of plant size on the nitrogen use strategy in an annual herb, Helianthus annuus (Sunflower). Acta Bot. Sinica 46: 889–895.
Zimdahl RL. 2004. Weed–Crop Competition: A Review 2nd. Blackwell Publishing, Ames, Iowa.
Zoschke A. 1990. Yield loss in tropical rice as influenced by the competition of weed flora and the timing of its elimination. In : Grayson BT, Green MB, Copping LG (eds). Pest Management in Rice. Elsevier Science, London. Pp. 301-313.

L A M P I R A N
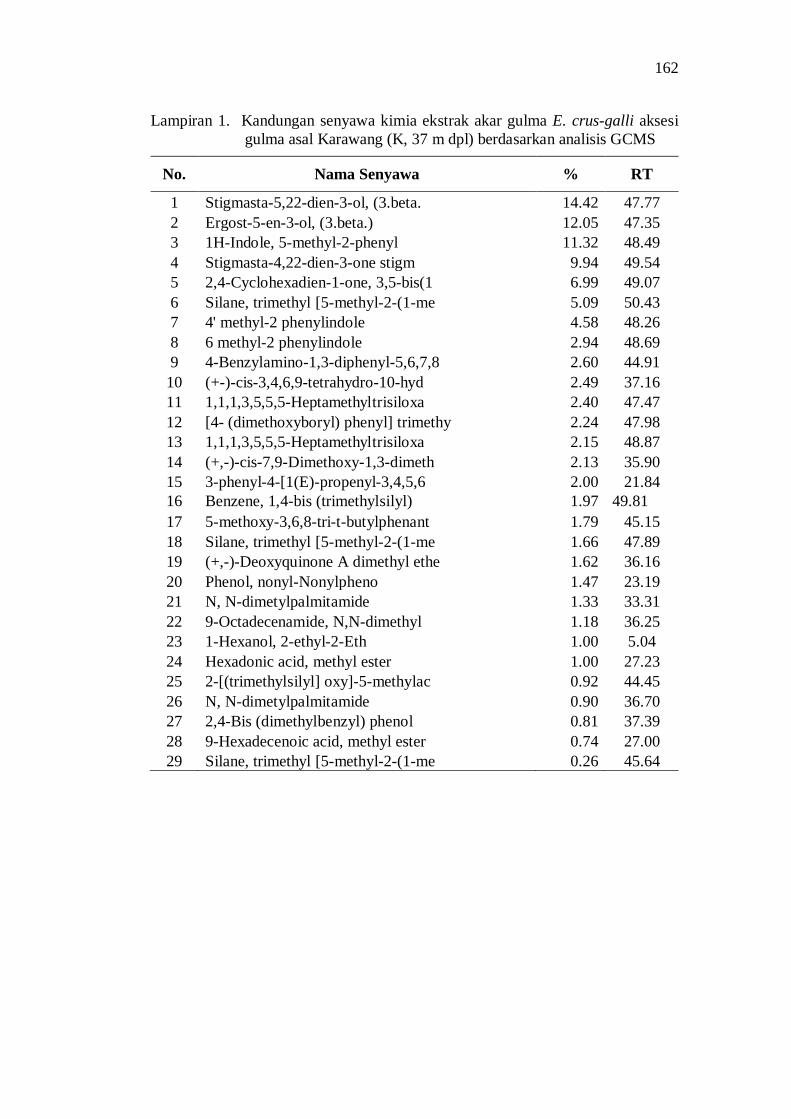
162
Lampiran 1. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Karawang (K, 37 m dpl) berdasarkan analisis GCMS
No. Nama Senyawa % RT
1 Stigmasta-5,22-dien-3-ol, (3.beta. 14.42 47.77 2 Ergost-5-en-3-ol, (3.beta.) 12.05 47.35 3 1H-Indole, 5-methyl-2-phenyl 11.32 48.49 4 Stigmasta-4,22-dien-3-one stigm 9.94 49.54 5 2,4-Cyclohexadien-1-one, 3,5-bis(1 6.99 49.07 6 Silane, trimethyl [5-methyl-2-(1-me 5.09 50.43 7 4' methyl-2 phenylindole 4.58 48.26 8 6 methyl-2 phenylindole 2.94 48.69 9 4-Benzylamino-1,3-diphenyl-5,6,7,8 2.60 44.91
10 (+-)-cis-3,4,6,9-tetrahydro-10-hyd 2.49 37.16 11 1,1,1,3,5,5,5-Heptamethyltrisiloxa 2.40 47.47 12 [4- (dimethoxyboryl) phenyl] trimethy 2.24 47.98 13 1,1,1,3,5,5,5-Heptamethyltrisiloxa 2.15 48.87 14 (+,-)-cis-7,9-Dimethoxy-1,3-dimeth 2.13 35.90 15 3-phenyl-4-[1(E)-propenyl-3,4,5,6 2.00 21.84 16 Benzene, 1,4-bis (trimethylsilyl) 1.97 49.81 17 5-methoxy-3,6,8-tri-t-butylphenant 1.79 45.15 18 Silane, trimethyl [5-methyl-2-(1-me 1.66 47.89 19 (+,-)-Deoxyquinone A dimethyl ethe 1.62 36.16 20 Phenol, nonyl-Nonylpheno 1.47 23.19 21 N, N-dimetylpalmitamide 1.33 33.31 22 9-Octadecenamide, N,N-dimethyl 1.18 36.25 23 1-Hexanol, 2-ethyl-2-Eth 1.00 5.04 24 Hexadonic acid, methyl ester 1.00 27.23 25 2-[(trimethylsilyl] oxy]-5-methylac 0.92 44.45 26 N, N-dimetylpalmitamide 0.90 36.70 27 2,4-Bis (dimethylbenzyl) phenol 0.81 37.39 28 9-Hexadecenoic acid, methyl ester 0.74 27.00 29 Silane, trimethyl [5-methyl-2-(1-me 0.26 45.64
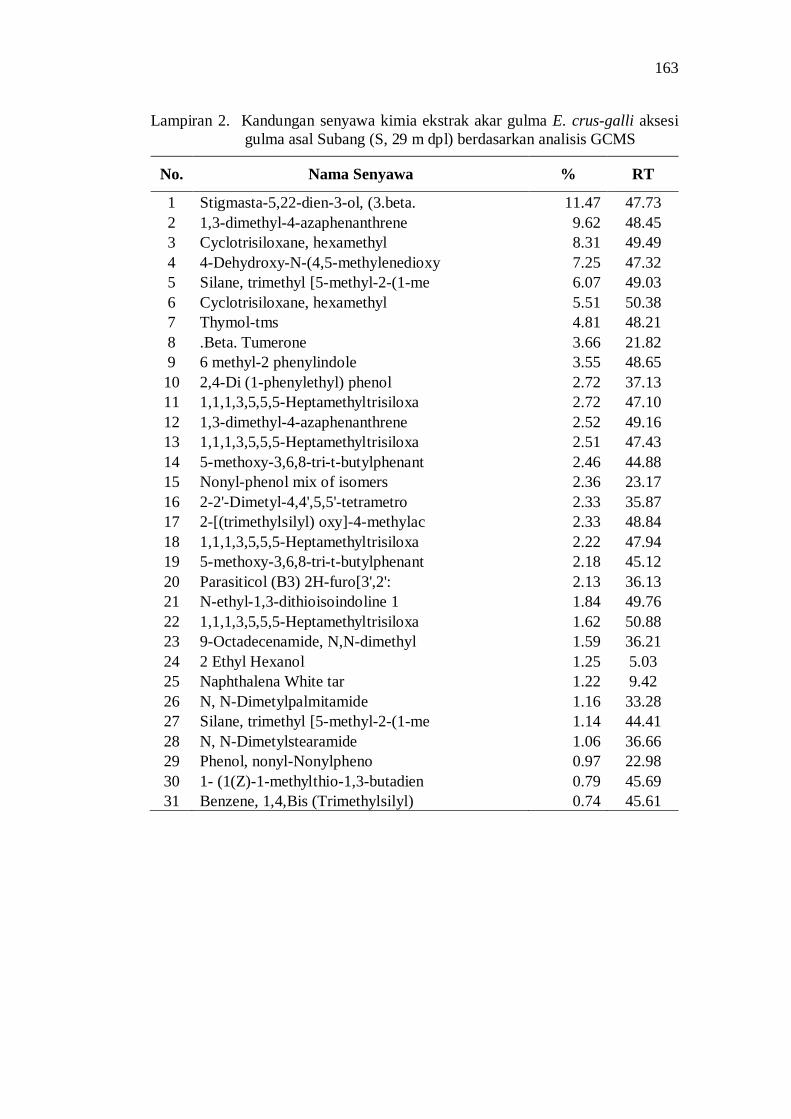
163
Lampiran 2. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Subang (S, 29 m dpl) berdasarkan analisis GCMS
No. Nama Senyawa % RT
1 Stigmasta-5,22-dien-3-ol, (3.beta. 11.47 47.73 2 1,3-dimethyl-4-azaphenanthrene 9.62 48.45 3 Cyclotrisiloxane, hexamethyl 8.31 49.49 4 4-Dehydroxy-N-(4,5-methylenedioxy 7.25 47.32 5 Silane, trimethyl [5-methyl-2-(1-me 6.07 49.03 6 Cyclotrisiloxane, hexamethyl 5.51 50.38 7 Thymol-tms 4.81 48.21 8 .Beta. Tumerone 3.66 21.82 9 6 methyl-2 phenylindole 3.55 48.65 10 2,4-Di (1-phenylethyl) phenol 2.72 37.13 11 1,1,1,3,5,5,5-Heptamethyltrisiloxa 2.72 47.10 12 1,3-dimethyl-4-azaphenanthrene 2.52 49.16 13 1,1,1,3,5,5,5-Heptamethyltrisiloxa 2.51 47.43 14 5-methoxy-3,6,8-tri-t-butylphenant 2.46 44.88 15 Nonyl-phenol mix of isomers 2.36 23.17 16 2-2'-Dimetyl-4,4',5,5'-tetrametro 2.33 35.87 17 2-[(trimethylsilyl) oxy]-4-methylac 2.33 48.84 18 1,1,1,3,5,5,5-Heptamethyltrisiloxa 2.22 47.94 19 5-methoxy-3,6,8-tri-t-butylphenant 2.18 45.12 20 Parasiticol (B3) 2H-furo[3',2': 2.13 36.13 21 N-ethyl-1,3-dithioisoindoline 1 1.84 49.76 22 1,1,1,3,5,5,5-Heptamethyltrisiloxa 1.62 50.88 23 9-Octadecenamide, N,N-dimethyl 1.59 36.21 24 2 Ethyl Hexanol 1.25 5.03 25 Naphthalena White tar 1.22 9.42 26 N, N-Dimetylpalmitamide 1.16 33.28 27 Silane, trimethyl [5-methyl-2-(1-me 1.14 44.41 28 N, N-Dimetylstearamide 1.06 36.66 29 Phenol, nonyl-Nonylpheno 0.97 22.98 30 1- (1(Z)-1-methylthio-1,3-butadien 0.79 45.69 31 Benzene, 1,4,Bis (Trimethylsilyl) 0.74 45.61
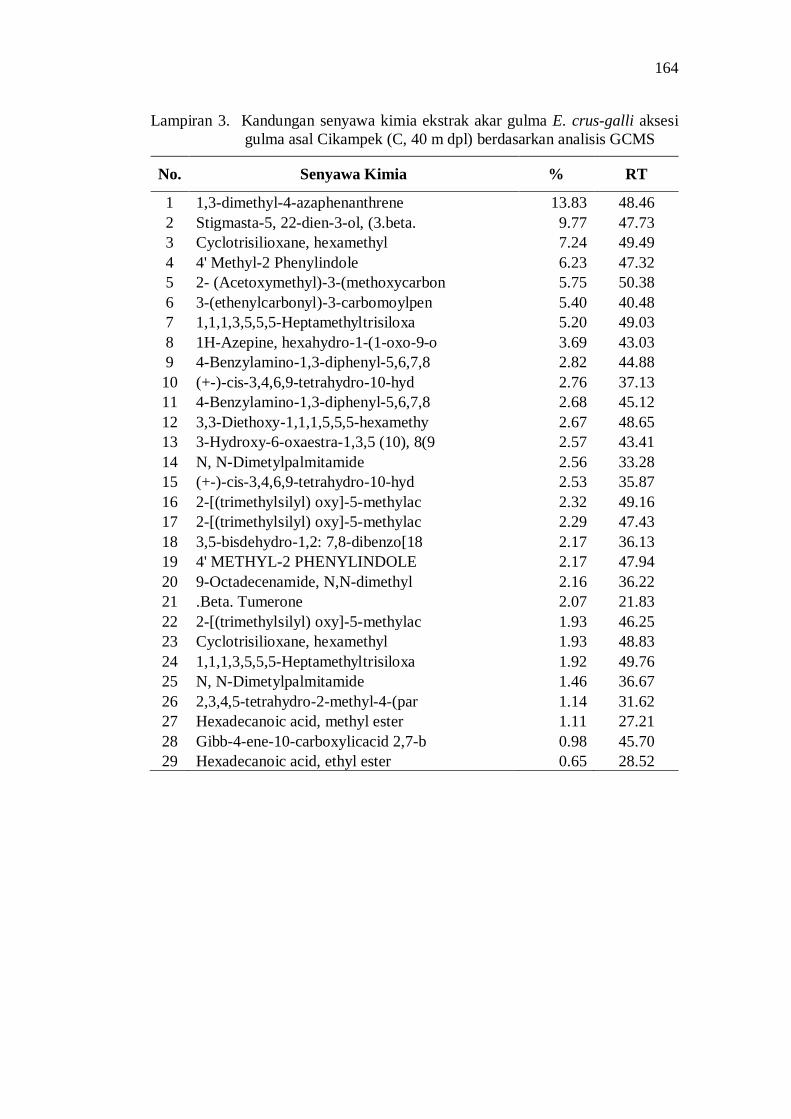
164
Lampiran 3. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Cikampek (C, 40 m dpl) berdasarkan analisis GCMS
No. Senyawa Kimia % RT
1 1,3-dimethyl-4-azaphenanthrene 13.83 48.46 2 Stigmasta-5, 22-dien-3-ol, (3.beta. 9.77 47.73 3 Cyclotrisilioxane, hexamethyl 7.24 49.49 4 4' Methyl-2 Phenylindole 6.23 47.32 5 2- (Acetoxymethyl)-3-(methoxycarbon 5.75 50.38 6 3-(ethenylcarbonyl)-3-carbomoylpen 5.40 40.48 7 1,1,1,3,5,5,5-Heptamethyltrisiloxa 5.20 49.03 8 1H-Azepine, hexahydro-1-(1-oxo-9-o 3.69 43.03 9 4-Benzylamino-1,3-diphenyl-5,6,7,8 2.82 44.88 10 (+-)-cis-3,4,6,9-tetrahydro-10-hyd 2.76 37.13 11 4-Benzylamino-1,3-diphenyl-5,6,7,8 2.68 45.12 12 3,3-Diethoxy-1,1,1,5,5,5-hexamethy 2.67 48.65 13 3-Hydroxy-6-oxaestra-1,3,5 (10), 8(9 2.57 43.41 14 N, N-Dimetylpalmitamide 2.56 33.28 15 (+-)-cis-3,4,6,9-tetrahydro-10-hyd 2.53 35.87 16 2-[(trimethylsilyl) oxy]-5-methylac 2.32 49.16 17 2-[(trimethylsilyl) oxy]-5-methylac 2.29 47.43 18 3,5-bisdehydro-1,2: 7,8-dibenzo[18 2.17 36.13 19 4' METHYL-2 PHENYLINDOLE 2.17 47.94 20 9-Octadecenamide, N,N-dimethyl 2.16 36.22 21 .Beta. Tumerone 2.07 21.83 22 2-[(trimethylsilyl) oxy]-5-methylac 1.93 46.25 23 Cyclotrisilioxane, hexamethyl 1.93 48.83 24 1,1,1,3,5,5,5-Heptamethyltrisiloxa 1.92 49.76 25 N, N-Dimetylpalmitamide 1.46 36.67 26 2,3,4,5-tetrahydro-2-methyl-4-(par 1.14 31.62 27 Hexadecanoic acid, methyl ester 1.11 27.21 28 Gibb-4-ene-10-carboxylicacid 2,7-b 0.98 45.70 29 Hexadecanoic acid, ethyl ester 0.65 28.52
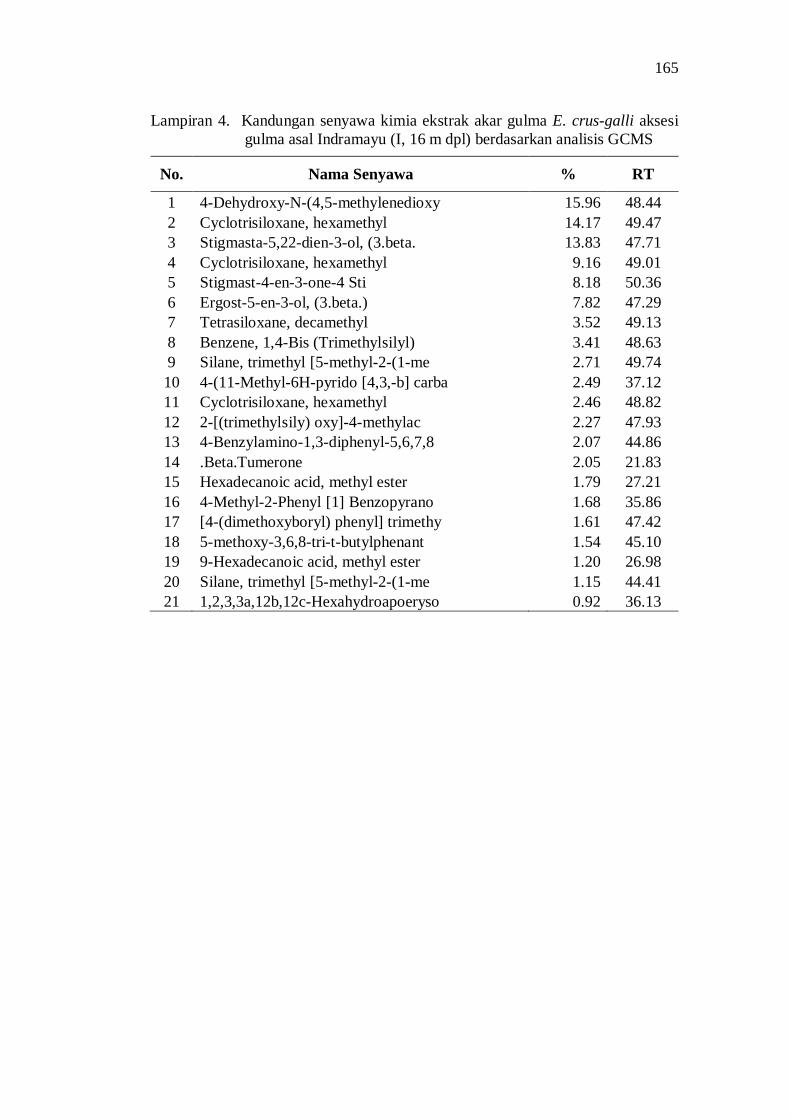
165
Lampiran 4. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Indramayu (I, 16 m dpl) berdasarkan analisis GCMS
No. Nama Senyawa % RT
1 4-Dehydroxy-N-(4,5-methylenedioxy 15.96 48.44 2 Cyclotrisiloxane, hexamethyl 14.17 49.47 3 Stigmasta-5,22-dien-3-ol, (3.beta. 13.83 47.71 4 Cyclotrisiloxane, hexamethyl 9.16 49.01 5 Stigmast-4-en-3-one-4 Sti 8.18 50.36 6 Ergost-5-en-3-ol, (3.beta.) 7.82 47.29 7 Tetrasiloxane, decamethyl 3.52 49.13 8 Benzene, 1,4-Bis (Trimethylsilyl) 3.41 48.63 9 Silane, trimethyl [5-methyl-2-(1-me 2.71 49.74 10 4-(11-Methyl-6H-pyrido [4,3,-b] carba 2.49 37.12 11 Cyclotrisiloxane, hexamethyl 2.46 48.82 12 2-[(trimethylsily) oxy]-4-methylac 2.27 47.93 13 4-Benzylamino-1,3-diphenyl-5,6,7,8 2.07 44.86 14 .Beta.Tumerone 2.05 21.83 15 Hexadecanoic acid, methyl ester 1.79 27.21 16 4-Methyl-2-Phenyl [1] Benzopyrano 1.68 35.86 17 [4-(dimethoxyboryl) phenyl] trimethy 1.61 47.42 18 5-methoxy-3,6,8-tri-t-butylphenant 1.54 45.10 19 9-Hexadecanoic acid, methyl ester 1.20 26.98 20 Silane, trimethyl [5-methyl-2-(1-me 1.15 44.41 21 1,2,3,3a,12b,12c-Hexahydroapoeryso 0.92 36.13
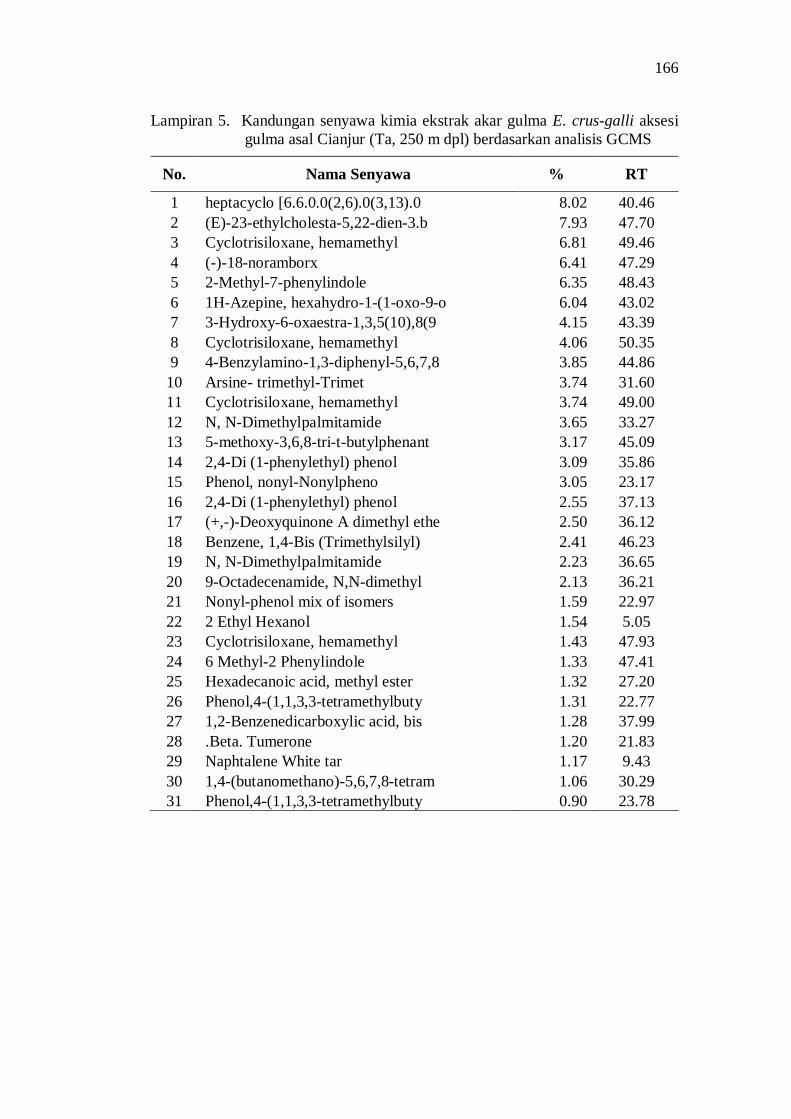
166
Lampiran 5. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Cianjur (Ta, 250 m dpl) berdasarkan analisis GCMS
No. Nama Senyawa % RT
1 heptacyclo [6.6.0.0(2,6).0(3,13).0 8.02 40.46 2 (E)-23-ethylcholesta-5,22-dien-3.b 7.93 47.70 3 Cyclotrisiloxane, hemamethyl 6.81 49.46 4 (-)-18-noramborx 6.41 47.29 5 2-Methyl-7-phenylindole 6.35 48.43 6 1H-Azepine, hexahydro-1-(1-oxo-9-o 6.04 43.02 7 3-Hydroxy-6-oxaestra-1,3,5(10),8(9 4.15 43.39 8 Cyclotrisiloxane, hemamethyl 4.06 50.35 9 4-Benzylamino-1,3-diphenyl-5,6,7,8 3.85 44.86
10 Arsine- trimethyl-Trimet 3.74 31.60 11 Cyclotrisiloxane, hemamethyl 3.74 49.00 12 N, N-Dimethylpalmitamide 3.65 33.27 13 5-methoxy-3,6,8-tri-t-butylphenant 3.17 45.09 14 2,4-Di (1-phenylethyl) phenol 3.09 35.86 15 Phenol, nonyl-Nonylpheno 3.05 23.17 16 2,4-Di (1-phenylethyl) phenol 2.55 37.13 17 (+,-)-Deoxyquinone A dimethyl ethe 2.50 36.12 18 Benzene, 1,4-Bis (Trimethylsilyl) 2.41 46.23 19 N, N-Dimethylpalmitamide 2.23 36.65 20 9-Octadecenamide, N,N-dimethyl 2.13 36.21 21 Nonyl-phenol mix of isomers 1.59 22.97 22 2 Ethyl Hexanol 1.54 5.05 23 Cyclotrisiloxane, hemamethyl 1.43 47.93 24 6 Methyl-2 Phenylindole 1.33 47.41 25 Hexadecanoic acid, methyl ester 1.32 27.20 26 Phenol,4-(1,1,3,3-tetramethylbuty 1.31 22.77 27 1,2-Benzenedicarboxylic acid, bis 1.28 37.99 28 .Beta. Tumerone 1.20 21.83 29 Naphtalene White tar 1.17 9.43 30 1,4-(butanomethano)-5,6,7,8-tetram 1.06 30.29 31 Phenol,4-(1,1,3,3-tetramethylbuty 0.90 23.78
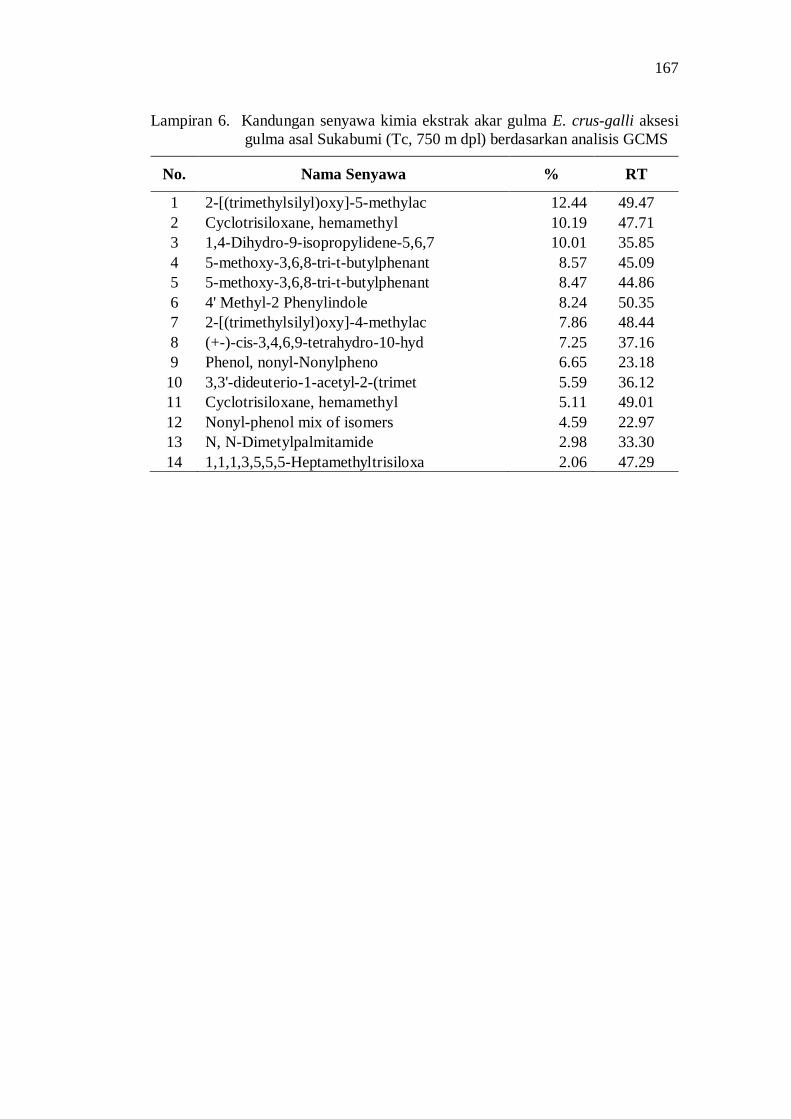
167
Lampiran 6. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Sukabumi (Tc, 750 m dpl) berdasarkan analisis GCMS
No. Nama Senyawa % RT
1 2-[(trimethylsilyl)oxy]-5-methylac 12.44 49.47 2 Cyclotrisiloxane, hemamethyl 10.19 47.71 3 1,4-Dihydro-9-isopropylidene-5,6,7 10.01 35.85 4 5-methoxy-3,6,8-tri-t-butylphenant 8.57 45.09 5 5-methoxy-3,6,8-tri-t-butylphenant 8.47 44.86 6 4' Methyl-2 Phenylindole 8.24 50.35 7 2-[(trimethylsilyl)oxy]-4-methylac 7.86 48.44 8 (+-)-cis-3,4,6,9-tetrahydro-10-hyd 7.25 37.16 9 Phenol, nonyl-Nonylpheno 6.65 23.18
10 3,3'-dideuterio-1-acetyl-2-(trimet 5.59 36.12 11 Cyclotrisiloxane, hemamethyl 5.11 49.01 12 Nonyl-phenol mix of isomers 4.59 22.97 13 N, N-Dimetylpalmitamide 2.98 33.30 14 1,1,1,3,5,5,5-Heptamethyltrisiloxa 2.06 47.29
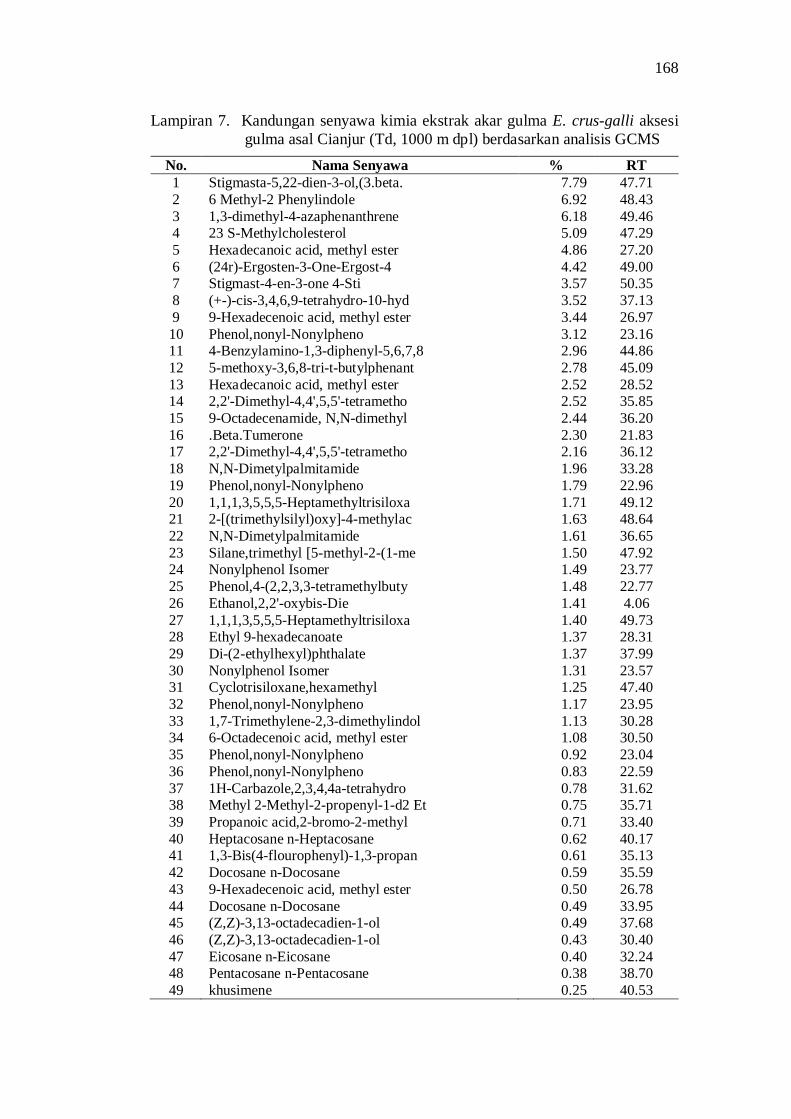
168
Lampiran 7. Kandungan senyawa kimia ekstrak akar gulma E. crus-galli aksesi gulma asal Cianjur (Td, 1000 m dpl) berdasarkan analisis GCMS
No. Nama Senyawa % RT 1 Stigmasta-5,22-dien-3-ol,(3.beta. 7.79 47.71 2 6 Methyl-2 Phenylindole 6.92 48.43 3 1,3-dimethyl-4-azaphenanthrene 6.18 49.46 4 23 S-Methylcholesterol 5.09 47.29 5 Hexadecanoic acid, methyl ester 4.86 27.20 6 (24r)-Ergosten-3-One-Ergost-4 4.42 49.00 7 Stigmast-4-en-3-one 4-Sti 3.57 50.35 8 (+-)-cis-3,4,6,9-tetrahydro-10-hyd 3.52 37.13 9 9-Hexadecenoic acid, methyl ester 3.44 26.97 10 Phenol,nonyl-Nonylpheno 3.12 23.16 11 4-Benzylamino-1,3-diphenyl-5,6,7,8 2.96 44.86 12 5-methoxy-3,6,8-tri-t-butylphenant 2.78 45.09 13 Hexadecanoic acid, methyl ester 2.52 28.52 14 2,2'-Dimethyl-4,4',5,5'-tetrametho 2.52 35.85 15 9-Octadecenamide, N,N-dimethyl 2.44 36.20 16 .Beta.Tumerone 2.30 21.83 17 2,2'-Dimethyl-4,4',5,5'-tetrametho 2.16 36.12 18 N,N-Dimetylpalmitamide 1.96 33.28 19 Phenol,nonyl-Nonylpheno 1.79 22.96 20 1,1,1,3,5,5,5-Heptamethyltrisiloxa 1.71 49.12 21 2-[(trimethylsilyl)oxy]-4-methylac 1.63 48.64 22 N,N-Dimetylpalmitamide 1.61 36.65 23 Silane,trimethyl [5-methyl-2-(1-me 1.50 47.92 24 Nonylphenol Isomer 1.49 23.77 25 Phenol,4-(2,2,3,3-tetramethylbuty 1.48 22.77 26 Ethanol,2,2'-oxybis-Die 1.41 4.06 27 1,1,1,3,5,5,5-Heptamethyltrisiloxa 1.40 49.73 28 Ethyl 9-hexadecanoate 1.37 28.31 29 Di-(2-ethylhexyl)phthalate 1.37 37.99 30 Nonylphenol Isomer 1.31 23.57 31 Cyclotrisiloxane,hexamethyl 1.25 47.40 32 Phenol,nonyl-Nonylpheno 1.17 23.95 33 1,7-Trimethylene-2,3-dimethylindol 1.13 30.28 34 6-Octadecenoic acid, methyl ester 1.08 30.50 35 Phenol,nonyl-Nonylpheno 0.92 23.04 36 Phenol,nonyl-Nonylpheno 0.83 22.59 37 1H-Carbazole,2,3,4,4a-tetrahydro 0.78 31.62 38 Methyl 2-Methyl-2-propenyl-1-d2 Et 0.75 35.71 39 Propanoic acid,2-bromo-2-methyl 0.71 33.40 40 Heptacosane n-Heptacosane 0.62 40.17 41 1,3-Bis(4-flourophenyl)-1,3-propan 0.61 35.13 42 Docosane n-Docosane 0.59 35.59 43 9-Hexadecenoic acid, methyl ester 0.50 26.78 44 Docosane n-Docosane 0.49 33.95 45 (Z,Z)-3,13-octadecadien-1-ol 0.49 37.68 46 (Z,Z)-3,13-octadecadien-1-ol 0.43 30.40 47 Eicosane n-Eicosane 0.40 32.24 48 Pentacosane n-Pentacosane 0.38 38.70 49 khusimene 0.25 40.53
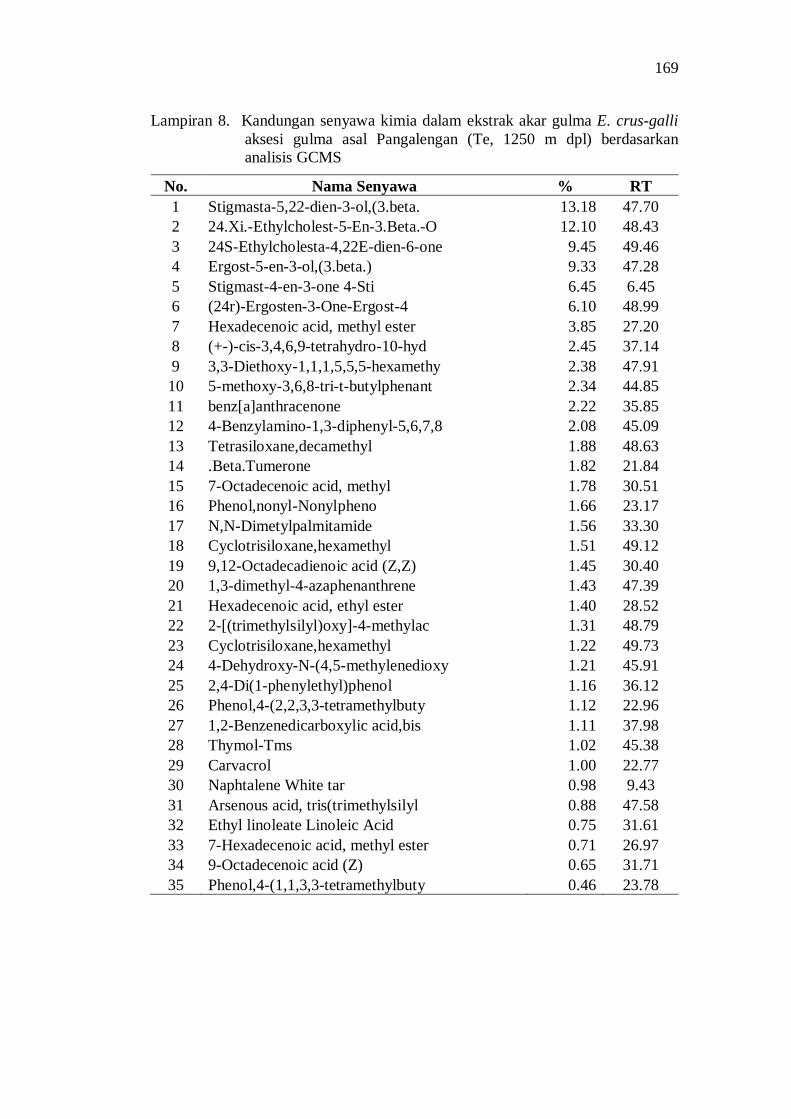
169
Lampiran 8. Kandungan senyawa kimia dalam ekstrak akar gulma E. crus-galli aksesi gulma asal Pangalengan (Te, 1250 m dpl) berdasarkan analisis GCMS
No. Nama Senyawa % RT 1 Stigmasta-5,22-dien-3-ol,(3.beta. 13.18 47.70 2 24.Xi.-Ethylcholest-5-En-3.Beta.-O 12.10 48.43 3 24S-Ethylcholesta-4,22E-dien-6-one 9.45 49.46 4 Ergost-5-en-3-ol,(3.beta.) 9.33 47.28 5 Stigmast-4-en-3-one 4-Sti 6.45 6.45 6 (24r)-Ergosten-3-One-Ergost-4 6.10 48.99 7 Hexadecenoic acid, methyl ester 3.85 27.20 8 (+-)-cis-3,4,6,9-tetrahydro-10-hyd 2.45 37.14 9 3,3-Diethoxy-1,1,1,5,5,5-hexamethy 2.38 47.91 10 5-methoxy-3,6,8-tri-t-butylphenant 2.34 44.85 11 benz[a]anthracenone 2.22 35.85 12 4-Benzylamino-1,3-diphenyl-5,6,7,8 2.08 45.09 13 Tetrasiloxane,decamethyl 1.88 48.63 14 .Beta.Tumerone 1.82 21.84 15 7-Octadecenoic acid, methyl 1.78 30.51 16 Phenol,nonyl-Nonylpheno 1.66 23.17 17 N,N-Dimetylpalmitamide 1.56 33.30 18 Cyclotrisiloxane,hexamethyl 1.51 49.12 19 9,12-Octadecadienoic acid (Z,Z) 1.45 30.40 20 1,3-dimethyl-4-azaphenanthrene 1.43 47.39 21 Hexadecenoic acid, ethyl ester 1.40 28.52 22 2-[(trimethylsilyl)oxy]-4-methylac 1.31 48.79 23 Cyclotrisiloxane,hexamethyl 1.22 49.73 24 4-Dehydroxy-N-(4,5-methylenedioxy 1.21 45.91 25 2,4-Di(1-phenylethyl)phenol 1.16 36.12 26 Phenol,4-(2,2,3,3-tetramethylbuty 1.12 22.96 27 1,2-Benzenedicarboxylic acid,bis 1.11 37.98 28 Thymol-Tms 1.02 45.38 29 Carvacrol 1.00 22.77 30 Naphtalene White tar 0.98 9.43 31 Arsenous acid, tris(trimethylsilyl 0.88 47.58 32 Ethyl linoleate Linoleic Acid 0.75 31.61 33 7-Hexadecenoic acid, methyl ester 0.71 26.97 34 9-Octadecenoic acid (Z) 0.65 31.71 35 Phenol,4-(1,1,3,3-tetramethylbuty 0.46 23.78
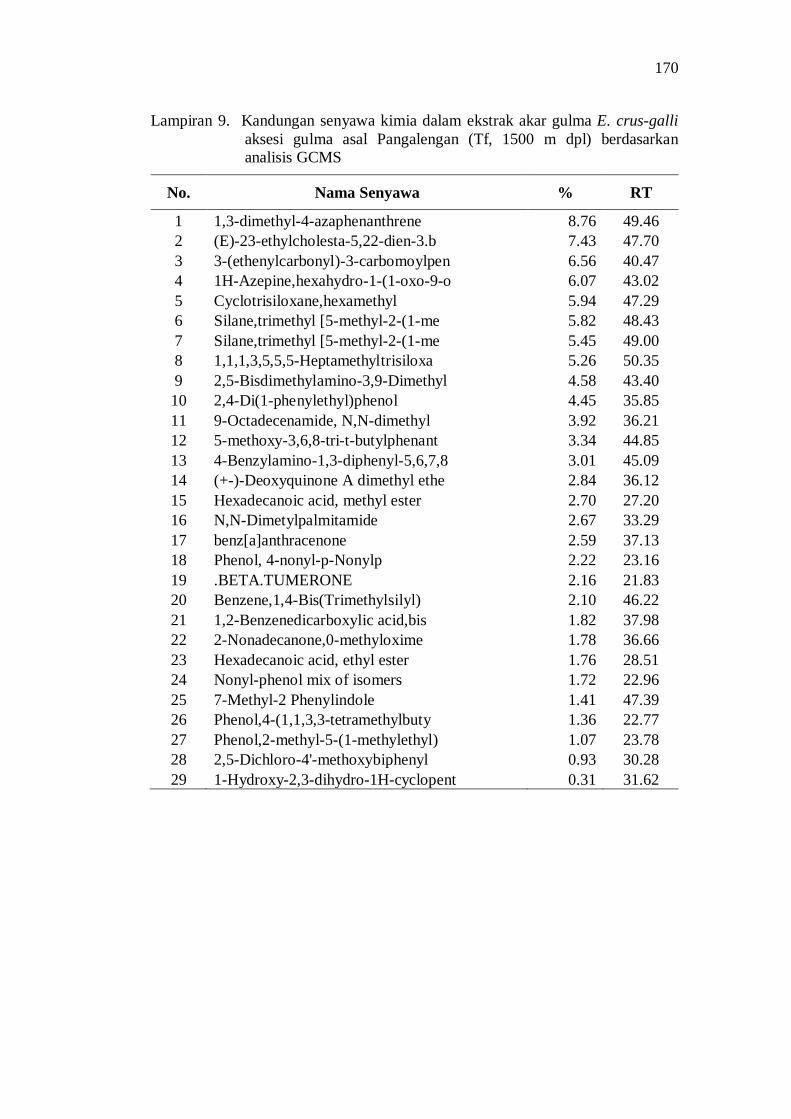
170
Lampiran 9. Kandungan senyawa kimia dalam ekstrak akar gulma E. crus-galli aksesi gulma asal Pangalengan (Tf, 1500 m dpl) berdasarkan analisis GCMS
No. Nama Senyawa % RT
1 1,3-dimethyl-4-azaphenanthrene 8.76 49.46 2 (E)-23-ethylcholesta-5,22-dien-3.b 7.43 47.70 3 3-(ethenylcarbonyl)-3-carbomoylpen 6.56 40.47 4 1H-Azepine,hexahydro-1-(1-oxo-9-o 6.07 43.02 5 Cyclotrisiloxane,hexamethyl 5.94 47.29 6 Silane,trimethyl [5-methyl-2-(1-me 5.82 48.43 7 Silane,trimethyl [5-methyl-2-(1-me 5.45 49.00 8 1,1,1,3,5,5,5-Heptamethyltrisiloxa 5.26 50.35 9 2,5-Bisdimethylamino-3,9-Dimethyl 4.58 43.40 10 2,4-Di(1-phenylethyl)phenol 4.45 35.85 11 9-Octadecenamide, N,N-dimethyl 3.92 36.21 12 5-methoxy-3,6,8-tri-t-butylphenant 3.34 44.85 13 4-Benzylamino-1,3-diphenyl-5,6,7,8 3.01 45.09 14 (+-)-Deoxyquinone A dimethyl ethe 2.84 36.12 15 Hexadecanoic acid, methyl ester 2.70 27.20 16 N,N-Dimetylpalmitamide 2.67 33.29 17 benz[a]anthracenone 2.59 37.13 18 Phenol, 4-nonyl-p-Nonylp 2.22 23.16 19 .BETA.TUMERONE 2.16 21.83 20 Benzene,1,4-Bis(Trimethylsilyl) 2.10 46.22 21 1,2-Benzenedicarboxylic acid,bis 1.82 37.98 22 2-Nonadecanone,0-methyloxime 1.78 36.66 23 Hexadecanoic acid, ethyl ester 1.76 28.51 24 Nonyl-phenol mix of isomers 1.72 22.96 25 7-Methyl-2 Phenylindole 1.41 47.39 26 Phenol,4-(1,1,3,3-tetramethylbuty 1.36 22.77 27 Phenol,2-methyl-5-(1-methylethyl) 1.07 23.78 28 2,5-Dichloro-4'-methoxybiphenyl 0.93 30.28 29 1-Hydroxy-2,3-dihydro-1H-cyclopent 0.31 31.62